


Потенциальные остеоиндуктивные эффекты наночастиц гидроксиапатита на мезенхимальные стволовые клетки посредством взаимодействия эндотелиальных клеток
Аннотация
Нано-гидроксиапатит (нано-ГА) привлек значительное внимание в области регенеративной медицины. Взаимодействия эндотелиальных клеток (ЕС) с мезенхимальными стволовыми клетками (МСК) необходимы для реконструкции кости, но способ взаимодействия нано-НА в этом процессе остается неизвестным. Здесь мы исследовали цитотоксичность и остеоиндуктивное действие наночастиц НА (HANP) на МСК, используя модель непрямого совместного культивирования, опосредованную ЭК, и выделили основные механизмы. Было обнаружено, что в субцитотоксической дозе HANP увеличивают жизнеспособность и экспрессию генов остеобластов, а также минерализованные узелки и продукцию щелочной фосфатазы МСК. Эти явления основаны на HIF-1α, секретируемом EC, который запускает сигнальный каскад ERK1 / 2. Кроме того, была создана двухэтапная математическая модель клеточного происхождения для количественного анализа влияния HIF-1α на остеогенную дифференцировку МСК. Он продемонстрировал, что HIF-1α оказывает дозозависимый стимулирующий эффект на скорость остеогенной дифференцировки МСК до 1500 пг / мл, что согласуется с приведенными выше результатами. Наши данные предполагают, что кооперативные взаимодействия между HANP, EC и MSC, вероятно, служат для стимуляции регенерации кости. Кроме того, двухэтапная модель клеточного происхождения полезна в системе in vitro для оценки потенциального влияния эффекторных молекул на инженерию костной ткани.
Введение
Восстановление костных дефектов, вызванных травмой, врожденным пороком развития или хирургической резекцией, представляет собой серьезную проблему для ортопедической хирургии [1]. Гидроксиапатит (ГА), типичная биоактивная керамика, использовался в качестве заменителя кости [2]. Однако нежелательные механические и остеоиндуктивные свойства ограничивают его клиническое применение [3]. В последние годы нано-ГК продемонстрировала более оптимальную лучшую биоактивность и улучшенные механические характеристики благодаря своим уникальным бионическим характеристикам и вызвала значительный интерес в биомедицинских областях, связанных с регенеративной медициной [4]. Когда нано-ГК имплантируется в дефекты кости, ему будут подвергаться многочисленные клетки, участвующие в восстановлении кости. Таким образом, необходимо оценить биологическое поведение нано-ГА. Несколько линий доказательств прямо продемонстрировали, что наночастицы HA (HANP) могут захватываться мезенхимальными стволовыми клетками пуповины человека Wharton, полученными из желе (hWJ-MSC), и клетками остеобластов, что приводит к усилению остеогенной дифференцировки [5,6,7]. Дуа и др. ранее сообщалось о способности HANPs способствовать интеграции сконструированного хряща в хрящ de novo [8]; Напротив, HANP ингибируют ангиогенную способность эндотелиальных клеток пупочной вены человека (HUVEC) [9]. Что касается здоровья человека, то необходимо более полное понимание влияния HANP на регенерацию кости, и постоянное применение искусственных костей на основе нанотехнологий делает такие исследования еще более актуальными.
Костная регенерация неизбежно сопровождается инвазией новообразований. ЭК представляют собой внутреннюю клеточную выстилку сосудистой системы, которая служит для пассивной доставки крови, а также играет роль в индукции, определении и управлении регенерацией органов, а также поддержании гомеостаза и метаболизма [10, 11]. МСК являются частью периэндотелиальной ниши и обладают способностью к самообновлению и множественной дифференцировке под воздействием определенных физиологических и биохимических микросредств в своих резидентных нишах [12]. Tsai et al. обнаружили, что ECs могут секретировать эндотелин-1, чтобы направлять МСК к дифференцировке остео- и хондро-клонов [13]. Кроме того, Салех и др. использовали анализ данных микроматрицы для идентификации белков, секретируемых HUVEC, и связанных с ними перекрестных сигнальных путей, которые взаимодействуют с мембраносвязанными рецепторами MSC для усиления пролиферации и остеогенной дифференцировки [14]. В инженерии костной ткани HANPs могут контактировать с новообразованиями и подвергаться эндоцитозу с помощью ECs, что, как было показано, изменяет физиологическую функцию этих клеток [9, 15]. Это также может влиять на окружающие клетки-остеопрогениторы и влиять на восстановление костей, изменяя паракринную передачу сигналов. Однако, хотя прямое влияние HANP на МСК было изучено, все еще отсутствует четкое понимание того, могут ли HANP косвенно вызывать остеогенную дифференцировку МСК через ЭК, что важно для нашего понимания влияния HANP в отношении для восстановления костей.
В этом исследовании, чтобы получить более полное представление о биологических эффектах HANP на взаимодействие между EC и MSC, была создана модель непрямого совместного культивирования с использованием HUVEC и hWJ-MSC. Используя эту систему, оценивали цитотоксичность и остеоиндуктивное действие HANP на hWJ-MSC через HUVEC-опосредованную паракринную передачу сигналов. Чтобы определить ключевые факторы, влияющие на HANP-индуцированные взаимодействия эндотелиальных клеток с MSC, растворимые факторы в супернатанте от HUVEC, которые были стимулированы HANP, были оценены с акцентом на связанные механизмы как на уровне гена, так и на уровне белка. Результаты показали, что фактор, индуцируемый гипоксией (HIF) -1α, играет решающую роль в этих взаимодействиях.
Чтобы количественно наблюдать и прогнозировать влияние HIF-1α на процесс остеогенеза, была создана математическая модель, которая объединяет двухэтапную клеточную линию с HIF-1α. Здесь, путем анализа эмпирических данных, была использована двухэтапная модель клеточного происхождения для прогнозирования количества МСК и степени дифференцировки в любой момент времени на основе определенной начальной плотности посева клеток и концентрации HIF-1α, что, в свою очередь, может дайте соответствующие рекомендации по начальным условиям культивирования и времени инкубации. Результаты этого исследования помогут пролить свет на взаимодействие между заменителями кости на основе наночастиц и биологическими системами, что может способствовать разработке инновационных биоматериалов для использования в регенеративной медицине.
Материалы и методы
Подготовка и определение характеристик частиц
HANP при 20 нм (np20), 20 * 80 нм (np80) и микрочастицы HA (m-HAP) с чистотой ≥ 99,0% были приобретены у Nanjing Emperor Nano Material Company Ltd (Нанкин, Китай). Размер и форму частиц наблюдали с помощью просвечивающей электронной микроскопии (ТЕМ; FEI Tecnai G2 Spirit Bio-Twin, FEI, Хиллсборо, Орегон, США) и сканирующей электронной микроскопии (SEM; LEO1530VP, Германия). Гидродинамический размер и дзета-потенциал частиц ГА (ГАП) определяли с помощью Zetasizer Nano ZS90 и Mastersizer 3000 (Malvern Instruments, Малверн, Великобритания).
Подготовка клеток и культивирование
Все протоколы экспериментов были одобрены этическим комитетом Нанкинского медицинского университета. HUVEC и hWJ-MSC были получены из свежей пуповины человека, как описано ранее, после получения письменного информированного согласия доноров [16, 17]. Вкратце, пуповину и пупочную вену промывали фосфатно-солевым буфером (PBS), содержащим 1% пенициллин и стрептомицин (PS; Hyclone, GE Healthcare Life Sciences, Pasching, Austria). Затем пупочную вену заполняли 0,1% коллагеназой I (Sigma, Сент-Луис, Миссури, США) и инкубировали в течение 15 мин при 37 ° C. После сбора HUVEC культивировали в среде ЕС (ЕСМ) (Sciencell, Сан-Диего, Калифорния, США).
Впоследствии кровеносные сосуды были удалены, а желе Уортона разрезали на 1 мм 2 кусочков, а затем поместите на 25 см 2 колбы для тканевых культур (Corning Incorporated, Корнинг, Нью-Йорк, США). Эти клетки инкубировали в L-DMEM (GIBCO Life Technology, Гранд-Айленд, Нью-Йорк, США) с добавлением 10% фетальной бычьей сыворотки (GIBCO) и 1% PS.
МСК hWJ оценивали для подтверждения фенотипа с использованием моноклональных антител к CD13, CD29, CD34, CD44, CD45, CD51 и CD105 (BD Biosciences, Сан-Хосе, Калифорния, США). HUVEC оценивали с использованием фактора фон Виллебранда (vWF; Shanghai ChangDao Biotech Co, Ltd., Шанхай, Китай). В этих экспериментах использовались HUVEC между пассажами 3-7 и hWJ-MSC между пассажами 3-5.
Суспензии частиц в концентрации 1 мг / мл в PBS затем разбавляли L-DMEM до конечной концентрации. Как показано на рис. 1, HUVEC инкубировали при указанной концентрации HAP в течение 18 часов. Культуральную среду центрифугировали при 15000 об / мин при 4 ° C в течение 15 минут, и супернатанты с добавлением 10% FBS использовали в качестве кондиционированной среды (CM) для hWJ-MSCs для выполнения следующих экспериментов. CM состояла из остеогенной среды, дополненной жидкостью для остеогенной индукции, которая содержала 10 мМ β-глицерофосфата, 50 мкг / мл L-аскорбиновой кислоты-2-фосфата и 0,1 мкМ дексаметазона (Sigma-Aldrich, Сент-Луис, Миссури, США). Кроме того, 2-метоксиэстрадиол (2-MeOE2) (Selleck Chemicals, Хьюстон, Техас, США) использовали в качестве специфического ингибитора HIF-1α. В группе 2-MeOE2 (+) hWJ-MSC культивировали с CM из HUVEC, которые предварительно обрабатывали 5 мкМ 2-MeOE2 в течение 40 минут перед добавлением HAP. PD98059 использовали в качестве специфического ингибитора MEK. В группе PD98059, hWJ-MSC культивировали с CM, содержащим 5 мкМ PD98059.
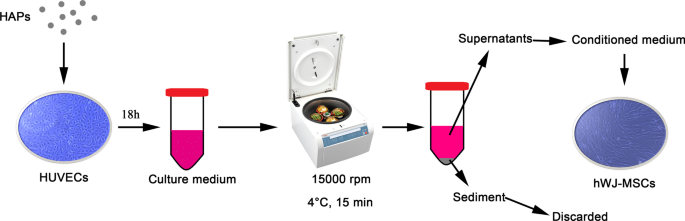
Иллюстрация непрямого совместного культивирования HAP / hWJ-MSC, опосредованного HUVEC. Сокращения: HAPs частицы гидроксиапатита, hWJ-MSCs пуповина человека мезенхимальные стволовые клетки Wharton, полученные из желе, HUVECs эндотелиальные клетки пупочной вены человека
Определение жизнеспособности и количества ячеек
Жизнеспособность клеток оценивали с помощью набора для анализа MTS (MTS; Bestbio, Beyotime Biotechnology, Шанхай, Китай). hWJ-MSC оставляли для прикрепления на 24 часа, а затем культивировали с CM в течение 24 и 72 часов. Поглощение формазана оценивали при 490 нм с использованием считывающего устройства для микропланшетов (SpectraMax M2; Molecular Devices LLC, Саннивейл, Калифорния, США). Поглощение также было преобразовано в количество клеток с использованием стандартных калибровочных кривых в тех же условиях.
Количественная полимеразная цепная реакция в реальном времени (ОТ-ПЦР)
Реагент TRIzol (Invitrogen, Carlsbad, CA, USA) использовали для выделения общей РНК из клеток hWJ-MSC, инкубированных в CM в течение указанного времени. Комплементарную ДНК транскрибировали из 1,0 мкг РНК с использованием набора PrimeScript First Strand cDNA Synthesis (TaKaRa, Tokyo, Japan) в термоциклере T3 (Mastercycler 5333; Eppendorf, Гамбург, Германия). Уровни экспрессии указанных генов анализировали с использованием набора FastStart Universal SYBR Green Master (ROX) (Roche, Базель, Швейцария) на системе количественной амплификации в реальном времени (7900HT Fast; Applied Biosystems, Foster City, CA, USA). Относительная экспрессия мРНК целевого гена была нормализована до GAPDH, а затем определена с использованием метода \ (2 ^ {{- \ Delta \ Delta C_ {t}}} \). Последовательности праймеров для генов-мишеней перечислены в таблице 1.
Окрашивание ализариновым красным S (ARS) и количественный анализ
hWJ-МСК культивировали в 12-луночных планшетах с остеогенной средой в течение до 14 дней, а затем наблюдали минерализацию внеклеточного матрикса с помощью окрашивания ARS (Leagene, Leagene Biotechnology, Пекин, Китай) после. Вкратце, образцы фиксировали абсолютным этиловым спиртом в течение 15 минут, а затем окрашивали в 1% (мас. / Об.) ARS (pH, 4,2) при комнатной температуре в течение 5 минут. Окрашенные клетки дважды промывали бидистиллированной водой и затем фотографировали. Для количественного анализа процесса минерализации в каждую лунку добавляли 300 мкл 10% (мас. / Об.) Моногидрата цетилпиридиния хлорида (BOMEI, BOMEI Biotechnology, Хэфэй, Китай) и планшеты инкубировали в течение 30 мин. Всего 90 мкл каждого образца переносили в 96-луночный планшет, и затем измеряли оптическую плотность при 405 нм в трех экземплярах.
Окрашивание щелочной фосфатазой (ЩФ) и количественный анализ
После того, как hWJ-MSC культивировали в 12-луночных планшетах с остеогенной средой в течение до 14 дней, проводили окрашивание ALP с использованием набора BCIP / NBT Alkaline Phosphatase Color Development Kit (Beyotime). Вкратце, МСК hWJ фиксировали в 4% параформальдегиде. Затем образцы окрашивали в смеси нитросинего тетразолия и 5-бром-4-хлор-3-индолилфосфата в течение 4 ч и фотографировали. Для количественной оценки синтеза ЩФ клетки лизировали в ледяном буфере для лизиса RIPA (Beyotime) в течение 30 мин. Лизаты клеток центрифугировали при 12000 об / мин при 4 ° C в течение 10 минут, и супернатанты подвергали количественному анализу ALP с использованием набора для анализа ALP (Beyotime). Оптическую плотность измеряли в трех экземплярах при 405 нм и пересчитывали в активность ЩФ с использованием стандартной кривой.
Иммуноферментный анализ (ELISA)
HUVEC были засеяны 2 × 10 5 ячеек / лунка. CM собирали из HUVEC, культивированных с HAP в течение 18 часов (дополнительный файл 1), а супернатант подвергали анализу ELISA (рис. 1). Набор для ELISA человеческого HIF-1α (Anhui Joyee Biotechnics, Anhui, China) использовали в соответствии с инструкциями производителя.
Иммунофлуоресценция
HUVEC высевали на предметные стекла в 12-луночные планшеты (7,6 × 10 4 клеток / лунка). После воздействия CM в течение 18 часов клетки фиксировали в 4% параформальдегиде (Biosharp, Пекин, Китай) и пермеабилизировали 0,1% Triton X-100 (Beyotime) в PBS перед инкубацией с 1% антителом против HIF-1α [EPR16897 ] (ab179483, Abcam, UK) в течение ночи при 4 ℃. Затем клетки инкубировали с 1% CoraLite594 - конъюгированным козьим анти-кроличьим IgG (H + L) (Proteintech, США) в темноте в течение 1 часа. Затем ядра окрашивали DAPI (Beyotime, Шанхай, Китай), который добавляли к клеткам и реагировали в течение 30 с. Образцы исследовали с помощью лазерного конфокального микроскопа (Olympus, Япония). Интенсивность флуоресценции определяли количественно с помощью программного обеспечения для анализа ImageJ v.1.4 (Bethesda, MD, США).
Вестерн-блоттинг
hWJ-МСК инкубировали в CM в течение 24 часов (киназа, регулируемая внеклеточными сигналами (ERK) 1/2, p-ERK1 / 2) или в остеогенной среде в течение 7 дней (фактор транскрипции runt-related (RUNX) -2, тип 1 коллаген / коллаген 1 (COL I)). Затем клетки лизировали в буфере для лизиса RIPA в течение 30 мин. Лизаты клеток центрифугировали, и супернатанты хранили при -20 ° C для анализа. После 12% SDS-PAGE белки переносили на мембрану из поливинилидендифторида (PVDF). В качестве первичных используемых антител были анти-p-ERK1 / 2, анти-ERK1 / 2, анти-RUNX-2, анти-COL I и GAPDH (1:1000, кроличьи поликлональные антитела; Cell Signaling Technology, Бостон, Массачусетс, США. ). После удаления несвязавшихся антител мембрану инкубировали со вторичными антителами в течение 1 ч. Сигнал на мембранах регистрировали с помощью системы визуализации хемилюминесцентного геля (LAS4000M; GE Healthcare Biosciences AB, Упсала, Швеция). Отношение p-ERK к ERK и RUNX-2 / COL I к GAPDH было количественно определено с помощью программного обеспечения для анализа ImageJ v.1.4 (Bethesda).
Оценка апоптоза клеток
hWJ-MSC были засеяны с плотностью 10 5 клеток на лунку в 6-луночных планшетах. Прилипшие клетки обрабатывали указанными концентрациями HIF-1α в течение указанного времени. Затем клетки собирали и метили FITC-аннексином V и PI (Fcmacs, Нанкин, Китай) в течение 15 минут в темноте. Все образцы были протестированы с использованием проточного цитометра FACScan (BD Bioscience, Сан-Хосе, Калифорния, США). Данные были проанализированы с помощью FlowJo v10 (BD Biosciences).
Двухэтапная модель клеточного происхождения
Принимая во внимание огромный потенциал HAP и сложность анализа системы совместного культивирования, требовалась математическая модель, которая могла бы обеспечить количественный анализ, и достоверное прогнозирование требовалось для понимания роли HIF-1α в остеогенной дифференцировке hWJ- МСК.
hWJ-MSC культивировали с 0, 300, 500, 1000, 1500, 2000, 3000 и 4000 пг / мл HIF-1α, а также с жидкостью для остеогенной индукции. После подбора данных для этих концентраций мы используем концентрации HIF-1α (продуцируемого HUVEC) в контрольной, m-HAP, np80 и np20 группах (240, 300, 325 и 375 пг / мл соответственно) для протестируйте подгоночные уравнения с помощью MATLAB (MathWorks, Натик, Массачусетс, США). Чтобы упростить модель, мы считали их сходные модели роста при различных концентрациях HIF-1α идентичными. Средняя степень дифференциации использовалась для соответствия уравнению степени дифференциации времени. Скорость пролиферации, скорость апоптоза и степень остеогенной дифференцировки МСК hWJ в разных группах определялись в определенное время.
Упрощенная двухэтапная модель клеточного клонирования, которая была подобна многоступенчатой модели клеточного клонирования [18, 19], была создана согласно экспериментальным данным. C 0 и C 1 представляют номер соты hWJ-MSC и терминальных сот соответственно. C 0 и C 1 регулируются:
$$ \ left \ {\ begin {gather} \ frac {{{\ text {d}} C_ {0}}} {{{\ text {d}} t}} =\ left [{\ frac {{K - C_ {0} - C_ {1}}} {K} p - (p - 1)} \ right] \ upsilon_ {0} C_ {0} \ hfill \\ \ frac {{{\ text {d}} C_ {1}}} {{{\ text {d}} t}} =\ left ({2 - \ frac {{K - C_ {0} - C_ {1}}} {K} p - p} \ right) \ upsilon_ {0} C_ {0} - AC_ {1} \ hfill \\ \ end {gather} \ right. $$Здесь p , подверженный влиянию HIF-1α и времени, представляет собой вероятность репликации hWJ-MSC. Соответственно, d =1 - p - это скорость дифференцировки, которая может быть получена путем подбора оценочных и экспериментальных данных о количестве клеток. Параметр v 0 определяет, насколько быстро клетки делятся на каждой стадии клонирования (в частности, v =Ln2 / c , где c - продолжительность клеточного цикла). Скорость апоптоза терминальных клеток обозначена A . Для простоты мы не учли, что скорость апоптоза будет незначительно меняться со временем, и поэтому A =4,5% - постоянная величина. К обозначает экологическую способность, потому что клетки не могут подвергаться неограниченной пролиферации [20]. HIF-1α увеличивает скорость дифференцировки hWJ-MSC, что приводит к скорости дифференцировки, моделируемой с помощью:
$$ \ begin {выровнено} d &=\ frac {{d_ {0}}} {{1 + (r * H) ^ {m}}} \\ p &=1 - d \\ \ end {выровнено} $$Здесь H представляет собой концентрацию HIF-1α; г 0 обозначает скорость дифференцировки при 0 пг / мл HIF-1α; r представляет собой интенсивность регуляции (здесь представляет интенсивность регуляции HIF-1α на МСК); а m соответствует коэффициенту Хилла [21], который показывает взаимосвязь между скоростью дифференцировки МСК и концентрацией HIF-1α.
Статистический анализ
Все данные, которые соответствуют требованиям нормальности и гомоскедастичности, выражены как среднее ± стандартное отклонение (SD) из трех или более независимых экспериментов. Программное обеспечение SPSS 24.0 (SPSS Inc., Чикаго, США) использовалось для выполнения статистического анализа с помощью одностороннего ANOVA или двустороннего ANONA. А P значение <0,05 считалось статистически значимым. Статистический анализ представлен с использованием GraphPad Prism 5 (GraphPad Software, La Jolla, CA).
Результаты
Характеристика HAP
Как показано на рис. 2, HAP были приготовлены с определенным размером и формой. Диаметр np20 с формой, близкой к сферической, составлял в среднем 20 нм, а np80 был стержнеобразным со средней длиной 80 нм и шириной 20 нм. M-HAP также были почти сферическими по форме и диаметром примерно 12 мкм. Все частицы имели отрицательный поверхностный заряд в L-DMEM. Было высказано предположение, что отрицательные значения дзета-потенциала оказывают значительное благоприятное влияние на прикрепление и пролиферацию костных клеток, а также на прямое соединение костей и образование новой кости [22, 23]. Частицы, которые наблюдались в L-DMEM, имеют тенденцию к агрегированию в водных системах. Их гидродинамический размер также был протестирован, что также могло быть важным фактором, влияющим на их биологическое поведение.

Характеристика HAP. Микрофотографии ПЭМ а np20 и b np80 и микрофотография c , сделанная с помощью SEM м-ГАП. г Характеристика HAP ( n =6). Сокращения: ТЕМ просвечивающая электронная микроскопия, SEM сканирующая электронная микроскопия, HA гидроксиапатит, м-ГАП микрочастицы ГАП
Косвенная токсичность HAP по отношению к hWJ-MSC
Чтобы оценить непрямую токсичность HAP для hWJ-MSC, жизнеспособность клеток измеряли с помощью анализов MTS. КМ с 50 мкг / мл HANP может значительно стимулировать жизнеспособность hWJ-MSC через 24 часа и 72 часа и особенно через 24 часа. Однако КМ с HANP 100 мкг / мл снижала жизнеспособность клеток на 15–20% по сравнению с контролем через 72 часа. Более того, КМ с 25 мкг / мл np20, но не np80, стимулировала жизнеспособность клеток через 24 часа. Эти явления подтвердили, что 50 мкг / мл HANP были субцитотоксической концентрацией, которая использовалась во всех последующих экспериментах (рис. 3).
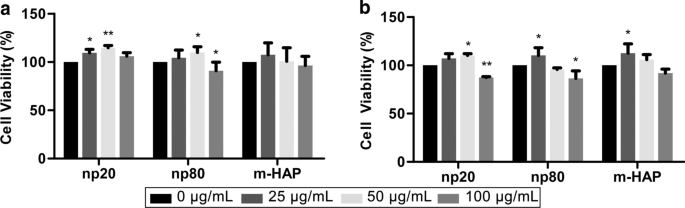
Непрямая токсичность HAP по отношению к МСК hWJ. Жизнеспособность hWJ-MSC, косвенно культивированных с HAP, измеряли для a 24 и b 72 ч. * P <0,05; ** P <0,01 по сравнению с контролем. Контрольная группа состояла из клеток, инкубированных в CM без обработки HAP, и жизнеспособность клеток была нормализована как процент от контроля. Сокращения: HAPs частицы гидроксиапатита, м-ГАП микрочастицы ГАП, hWJ-МСК мезенхимальные стволовые клетки пуповины человека, полученные из студня Wharton
Косвенный остеоиндуктивный эффект HAP на hWJ-MSC
Чтобы идентифицировать непрямые остеоиндуктивные эффекты HANP на hWJ-MSC, экспрессию остеогенных генов оценивали с помощью количественного анализа RT-PCR. Уровень транскрипции связанного с runt фактора транскрипции 2 (RUNX-2) в группах HANP, особенно np20, демонстрировал заметное повышение с 7 по 21 день (фиг. 4a). Экспрессия гена коллагена типа I (Col I) в группе np20 продемонстрировала усиление с 7 по 21 день, тогда как группа np80 продемонстрировала устойчивое увеличение с 7 по 14 день (фиг. 4b). Уровень мРНК остеокальцина (OCN) явно повышался в группах HANP на 14 день, что указывает на ускоренную скорость остеогенеза (фиг. 4c). Уровни мРНК щелочной фосфатазы (ЩФ) явно увеличиваются в группах HANP (рис. 4d), что указывает на остеогенную дифференцировку МСК hWJ. Однако группа m-HAP продемонстрировала ограниченные изменения в отношении уровней этих трех остеогенных генов по сравнению с контрольной группой (рис. 4). Кроме того, экспрессия маркеров плюрипотентности, NANOG, OCT3 / 4 и SOX2, снизилась в группах HANP по сравнению с контролем (рис. 4e – g), что означает, что hWJ-MSC в группах HANP дифференцировались, особенно в группа NP20. Сходные результаты были получены с помощью вестерн-блоттинга (рис. 5c, d), что указывает на то, что группа np 20 может косвенно усиливать экспрессию RUNX-2 и COL I в hWJ-MSC.
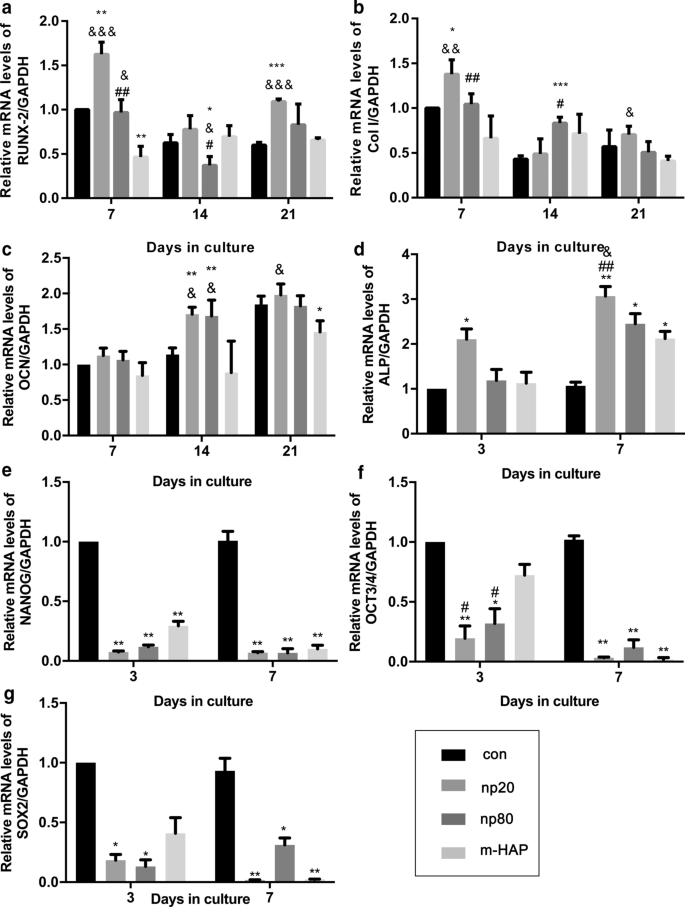
Косвенные эффекты HAP на экспрессию генов, связанных с остеогенной дифференцировкой. а RUNX-2, б Столбец I, c OCN, d ALP, e NANOG, f OCT3 / 4 и g Уровни гена SOX2 в hWJ-MSC, культивированных с CM в течение указанного времени. * P <0,05; ** P <0,01; *** P <0,001 по сравнению с контрольной группой; & P <0,05; && P <0,01; &&& P <0,001 по сравнению с группой m-HAP; # P <0,05; ## P <0,01 по сравнению с группой np20. Клетки, инкубированные в остеогенной среде без обработки HAP, использовали в качестве контрольной группы. Сокращения: HAPs частицы гидроксиапатита, м-ГАП микрочастицы ГАП, hWJ-МСК пуповина человека мезенхимальные стволовые клетки Wharton, полученные из желе, RUNX-2 фактор транскрипции 2, связанный с runt, Col I коллаген I типа, OCN остеокальцин, ALP щелочная фосфатаза, SOX 2 HMG-box 2, относящийся к SRY
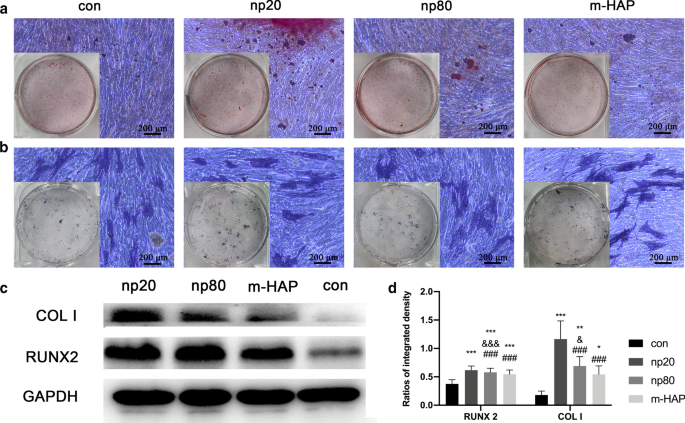
Косвенное влияние HAP на отложение внеклеточного кальция и активность ЩФ. МСК hWJ инкубировали в остеогенной среде в течение 14 дней. а Затем визуализировали внеклеточное отложение кальция с помощью окрашивания по ARS; б Активность ALP hWJ-MSC оценивали с помощью окрашивания ALP, масштабные линейки:200 мкм. c Вестерн-блоттинг показал экспрессию RUNX-2 и COL I МСК hWJ в остеогенной среде на 7 день. d Денситометрические измерения RUNX-2 и COL I из части ( c ). Клетки, инкубированные в остеогенной среде без обработки HAP, использовали в качестве контрольной группы. * P <0,05; ** P <0,01; *** P <0,001 по сравнению с контрольной группой; & P <0,05; &&& P <0,001 по сравнению с группой m-HAP; ### P <0,001 по сравнению с группой np20. Сокращения: HAPs частицы гидроксиапатита, м-ГАП микрочастицы ГАП, hWJ-МСК пуповина человека мезенхимальные стволовые клетки Wharton, полученные из желе, ALP щелочная фосфатаза
Чтобы визуально наблюдать непрямое остеоиндуктивное действие HANP на hWJ-MSC, клетки инкубировали с указанной остеогенной средой в течение 14 дней с последующим окрашиванием ARS и ALP. Как показано на фиг. 5a, b, повышенное количество минерализованных клубеньков и более высокая активность ALP hWJ-MSC наблюдались в группах HANP по сравнению с m-HAP и контрольной группами. Кроме того, m-HAP, как и контроль, продемонстрировал ограниченное влияние на остеогенную дифференцировку hWJ-MSC.
HAP активировали передачу сигналов ERK1 / 2 в hWJ-MSC, косвенно культивируемых совместно с HUVEC
Чтобы исследовать влияние HAP на паракринную функцию HUVEC, использовали иммунофлуоресценцию и ELISA-тесты для идентификации возможного белка, способствующего остеогенной дифференцировке hWJ-MSC. Как показано на рис. 6a – c, внутриклеточное и внеклеточное производство HIF-1α значительно облегчалось HANP (особенно np20), в то время как влияние m-HAP на его продукцию было ограниченным.
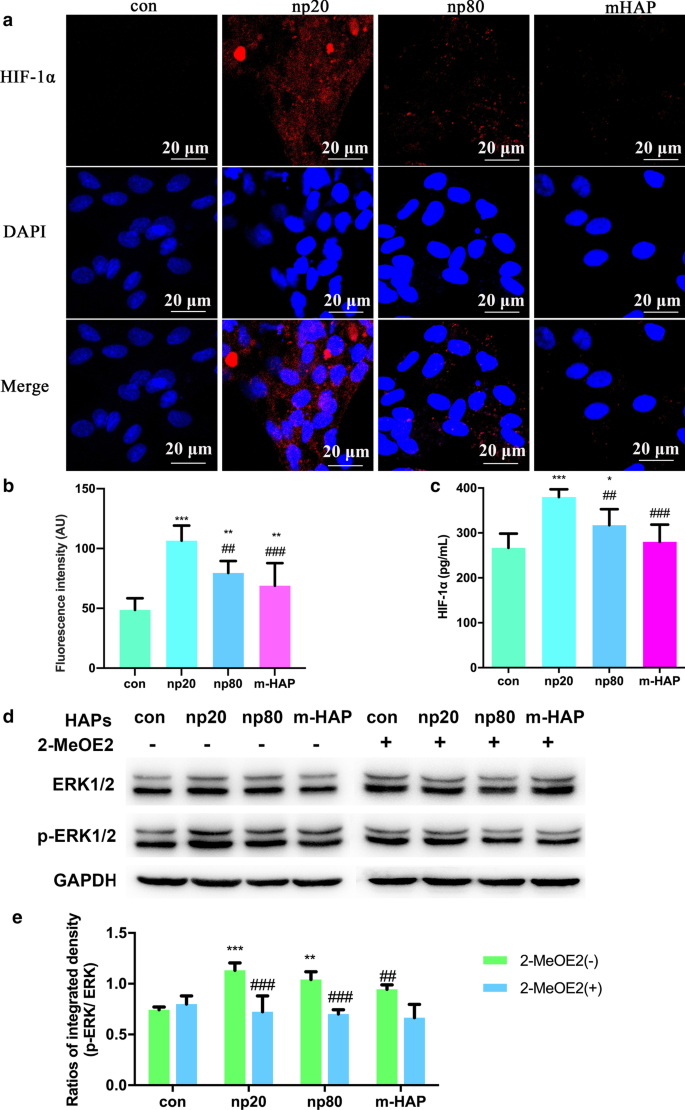
HAP активируют передачу сигналов ERK1 / 2 в hWJ-MSC, косвенно совместно культивируемых с HUVEC. а Иммунофлуоресценция HIF-1α выполняется в HUVEC, обработанных с / без HAP в течение 18 часов. б Интенсивность флуоресценции HIF-1α из части ( a ). c Внеклеточную концентрацию HIF-1α в культуральной среде HUVEC, обработанных с / без HAP в течение 18 часов, измеряли с помощью ELISA. Масштабные линейки:20 мкм. * P <0,05; ** P <0,01; *** P <0,001 по сравнению с контролем; ## P <0,01; ### P <0,001 по сравнению с группой np20. Клетки без обработки HAP использовали в качестве контрольной группы. МСК hWJ обрабатывали КМ в течение 24 часов. г Вестерн-блоттинг, указывающий на активацию ключевых киназ в путях ERK1 / 2. е Денситометрические измерения p-ERK1 / 2 из части ( b ). ** P <0,01; *** P <0,001 по сравнению с контролем; ## P <0,01, ### P <0,001 по сравнению с группой 2-MeOE2 (-). Клетки, инкубированные в CM без обработки HAP, использовали в качестве контрольной группы. Сокращения: HAPs частицы гидроксиапатита, м-ГАП микрочастицы ГАП, hWJ-МСК пуповина человека мезенхимальные стволовые клетки Wharton, полученные из желе, HUVECs эндотелиальные клетки пупочной вены человека, ERK киназа, регулируемая внеклеточными сигналами, HIF-1α фактор, индуцируемый гипоксией 1α
Чтобы более точно понять путь передачи сигнала дифференцировки hWJ-MSCs, активируемых HIF-1α, мы исследовали ключевые регуляторы пути ERK1 / 2 с помощью Вестерн-блоттинга. Как показано на фиг. 6d, e, в то время как уровни белка общего ERK1 / 2 оставались неизменными, уровни p-ERK1 / 2 были отчетливо повышены в hWJ-MSCs, культивируемых с HANP, и это было особенно верно в группе np20. Однако m-HAP мало влиял на уровни p-ERK1 / 2 в hWJ-MSC, аналогично его влиянию на продукцию HIF-1α в HUVEC. Важно отметить, что повышенные уровни p-ERK1 / 2 в hWJ-MSC, активированные HIF-1α, могут блокироваться 2-MeOE2, специфическим ингибитором HIF-1α, что указывает на то, что HIF-1α функционирует выше сигнального пути ERK1 / 2. в hWJ-MSC.
HIF-1α способствует остеогенной дифференцировке hWJ-MSC через путь ERK1 / 2
Чтобы определить, был ли HIF-1α необходим для наблюдаемой стимуляции остеогенной дифференцировки hWJ-MSC, к этим культурам клеток применяли специфический ингибитор HIF-1α (2-MeOE2). Как показано на фиг. 7, отложение минерализованного матрикса и активность ЩФ в группе hWJ-MSC, обработанной 2-MeOE2 (+), были ослаблены, культивируемые в остеогенной среде, что указывает на то, что HIF-1α необходим для остеогенной дифференцировки hWJ- МСК. Основываясь на этих результатах, мы дополнительно исследовали роль пути ERK1 / 2 в остеогенной дифференцировке hWJ-MSCs, активируемых HIF-1α. Отложение минерализованного матрикса и активность ALP в hWJ-MSC, культивированных с остеогенной средой, подавлялись после введения PD98059, специфического ингибитора MEK.
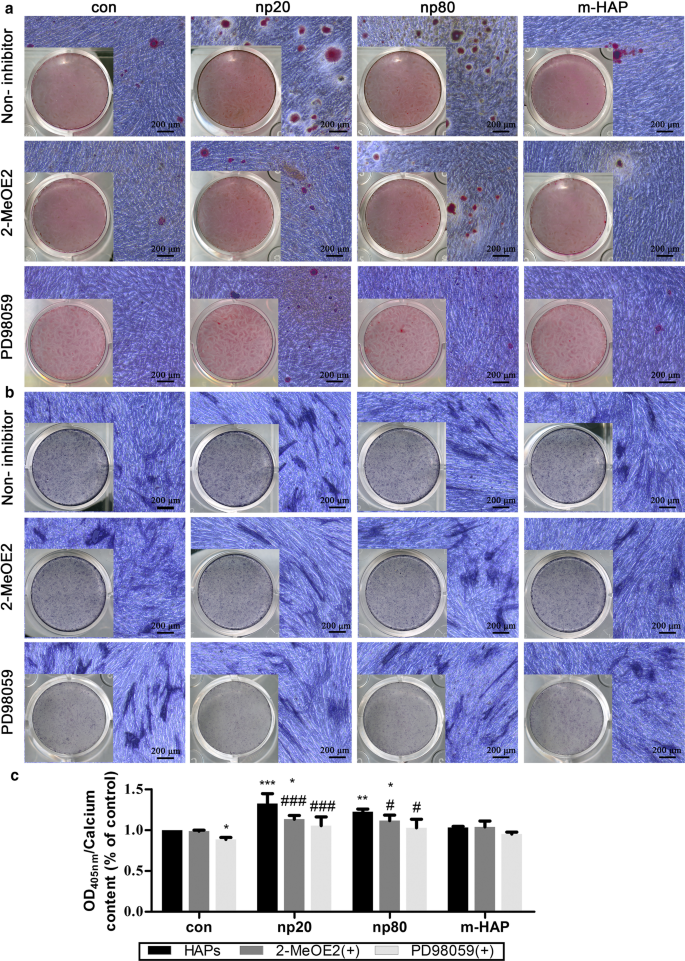
HIF-1α способствует остеогенной дифференцировке МСК hWJ через путь ERK1 / 2. hWJ-MSC инкубировали в остеогенной среде с PD98059 или без него в течение 14 дней. а Отложение внеклеточного кальция визуализировали с помощью окрашивания ARS. б Активность ЩФ hWJ-МСК оценивали с помощью окрашивания ЩФ. Масштабные линейки:200 мкм. c Quantitative analysis of extracellular calcium matrix. * P <0,05; ** P <0,01; *** P < 0.001 versus control; # P <0,05; ### P < 0.001 versus the np20 group. Cells incubated in osteogenic medium without HAPs and PD98059 treatment were used as the control group. Abbreviations:HAPs hydroxyapatite particles, m-HAP micro-sized HAP particles, hWJ-MSCs human umbilical cord Wharton’s jelly-derived mesenchymal stem cells, HUVECs human umbilical vein endothelial cells, ERK extracellular signal-regulated kinase, HIF-1α hypoxia-inducible factor 1α, ALP alkaline phosphatase
Two-Stage Cell-Lineage Models
To quantitatively reveal the intrinsic connection between the concentration of HIF and osteogenic differentiation of hWJ-MSCs, 0–4000 pg/mL of HIF-1α was used to treat eight groups of hWJ-MSCs. A two-stage cell-lineage mathematical model was used to analyze the proliferation, apoptosis, and osteogenic differentiation rates of these hWJ-MSCs treated with different concentrations of HIF-1α. As shown in Fig. 8a, fitting data were employed to obtain the simulated formula (\(\frac{d}{{d_{0} }} =\frac{1}{{0.14H^{2} - 0.43H + 1}}\)) and curve (blue curve), showing that the differentiation rate first increased and then decreased with the increase in HIF concentration. More specifically, the differentiation rate reached a peak at 1500 pg/mL HIF-1α.

Two-stage cell-lineage models. hWJ-MSCs were incubated in a defined concentration of HIF-1α for the indicated times. а Relative differentiation rates of hWJ-MSCs at different concentrations of HIF-1α, b relative ALP activity (differentiation degrees of hWJ-MSCs) and relative osteoblast cells on different culture days. c Three-dimensional surface of differentiation degree evolving with time and HIF-1α. Cell number with an initial seeding density of 1500 cells per well (in a 96-well plate), as well as d 0, e 375, and f 1500 pg/mL HIF-1α. Abbreviations:hWJ-MSCs human umbilical cord Wharton’s jelly-derived mesenchymal stem cells, HIF-1α hypoxia-inducible factor 1α, ALP alkaline phosphatase
According to our study, the concentrations of HIF-1α produced by HUVECs in the control, m-HAP, np80, and np20 groups were 240, 300, 325, and 375 pg/mL, respectively. The black square represents the differentiation rate promoted by 240, 300, 325, and 375 pg/mL HIF-1α, and this matches well with the simulated curve. In addition, the differentiation degrees of the hWJ-MSCs (relative ALP activity) treated with different concentrations of HIF-1α increased similarly increased with time. Therefore, in order to simplify the model, we consider their growth patterns to be similar or even identical. We found that the increase in ALP activity from Day 0 to Day 7 was proportional to the number of osteoblasts, and osteoblast cells reached their peak at the platform period. Therefore, after fitting the ALP activity with the relative osteoblasts cell number curve (osteoblasts cell number / maximum osteoblasts cell number), the maximum of ALP activity was predicted and relative ALP activity (ALP activity/ maximum of ALP activity), representing the differentiation degree, was acquired (Fig. 8b). Combining these two studies, the three-dimensional surface of the differentiation degree evolving with time and HIF-1α was obtained (Fig. 8c).
In order to estimate the optimal culture time, it was necessary to simulate cell numbers. After elucidating the differentiation rate under different concentrations of HIF-1α, we simulated the size of each population. The simulation utilized an initial seeding density of 1500 cells per well (in a 96-well plate) using different concentrations of HIF-1α. As shown in Fig. 8d–f, the experimental data (black square) match well with the simulated total cell numbers (blue curve), which is a sum of the number of hWJ-MSCs and osteoblasts, and this supports the ability of this model to predict the number of hWJ-MSCs and osteoblasts at any time point. Moreover, the osteoblast cell number approached the platform period at 21, 18, and 15 days with 0, 375, and 1500 pg/mL HIF-1α, respectively. This model provides the optimum culture time for guiding tissue engineering, and it also provides direct evidence that HIF-1α accelerates the osteogenic differentiation of hWJ-MSCs.
Обсуждение
With recent advances in nanobiomaterials, nano-based artificial bone substitutes have been an area of intense investigation. The accumulating evidence suggests that there are complex interactions between cells and nanobiomaterials due to their capacity to penetrate cell membranes and increase internal retention times [24, 25]. A previous study revealed that collagen/alginate nanofilms can adsorb onto the MSC membrane to activate intracellular signaling cascades and promote osteogenic differentiation [26]. Elegant experiments by Wu and his colleagues clearly demonstrated that TiO2 nanotubes can improve vascularization and osteogenic differentiation by facilitating paracrine effects and cell junctions via EC-MSC interactions [27]. For the purpose of developing excellent candidates for bone tissue engineering, it is necessary to clarify the direct crosstalk between nano-based bone substitutes and cells implicated in bone repair as well as their indirect interactions. However, our current understanding of this is still limited. In the present study, we utilized an indirect co-culture model to further elucidate the biological effects of HANPs on MSCs in regard to the indirect interactions mediated by ECs.
Cytotoxicity is a primary issue for assessing the biocompatibility of any nanobiomaterial. Although our previous study found that HANPs did not directly influence the viability or apoptosis of hWJ-MSCs, they may still exert different impacts via the mediation of other cells [28]. Thus, it was necessary to evaluate the cytotoxic effects of HANPs on hWJ-MSCs mediated by HUVECs. Interestingly, after incubation in CM for 24 h and 72 h, hWJ-MSC viability was maintained and even elevated in the 0–50 µg/mL HANP groups, especially in the np20 group, indicating the existence of effector molecules in the CM. When the concentration of HANPs reached 100 µg/mL, they became cytotoxic to the hWJ-MSCs. However, 0–100 µg/mL m-HAP had no influence on hWJ-MSC viability (Fig. 3). Jiang et al. have shown that engineered nanoparticles of a particular size can have distinct endocytic routes and kinetics associated with altered downstream signaling involved in regulating target cell functions [29]. In our previous study, we showed that np20 and np80 were endocytosed by HUVECs, and this was followed by morphologic changes and the appearance of large vacuoles, indicating the activated state of the HUVECs. Additionally, np20, with their faster uptake speed and increased accumulation, might result in a stronger activation of HUVECs, possibly resulting in increased hWJ-MSC viability via paracrine signaling. Conversely, few m-HAPs can be endocytosed by HUVECs, and this might account for their limited influence on the metabolism of hWJ-MSCs [9].
To further explore the potential osteoinductive effect of activated HUVECs, a subcytotoxic dose of 50 µg/mL HAPs was used in subsequent studies. The CM collected from the activated HUVECs promoted extracellular calcium deposition, ALP activity, and osteogenic proteins expression in hWJ-MSCs, as well as the mRNA expression of osteogenic genes (Figs. 4, 5). Runx2, an essential transcription factor involved in specifying the osteoblast lineage [30], showed a substantial enhancement in the np20 group, indicating a strong osteoinductive effect on hWJ-MSCs (Fig. 4a and Fig. 5c, d). The np20 group demonstrated a 1.5-fold improvement in COLI expression at Day 7 (Figs. 4b, 5c, d) and a double increase at Day 14, which implied the presence of additional differentiated osteoblasts in the HANP-treated groups (Fig. 4b) [30]. OCN is a mature stage bone marker [31], and this gene showed a significant increase in the HANP groups at Day 14 (Fig. 4c), indicating that np20 and np80 can accelerate bone maturation compared to m-HAP. ALP is an early marker of osteoblast differentiation, and it obviously increased with culture time in each group, especially the np20 group, revealing that additional transformation occurred from MSCs to osteoblasts (Fig. 4d). Pluripotency markers, NANOG, OCT3/4, and SOX2 imply the capacity for differentiation [32]. As shown in Fig. 4e–g, the decreased expression in the genes of the HANP groups implied that most of the hWJ-MSCs in HAP groups had transformed into osteoblasts.
Our data demonstrated that the endocytosis of HANPs by HUVECs was associated with an improved osteogenic differentiation of hWJ-MSCs. However, the cause of this outcome is currently unclear. In terms of the paracrine function of HUVECs, we focused on soluble differentiation-inducing proteins in the supernatant of activated HUVECs. HIF-1α signaling is essential in coupling ossification and angiogenesis during bone regeneration [33, 34]. Heikal et al. reported that injured ECs secrete more HIF-1α even under normoxia conditions [35]. It has also been shown that exposure to HANPs inhibits the angiogenic ability of HUVECs [9]. Thus, we measured the concentrations of HIF-1α in the CM, and the results showed that the HIF-1α content increased in the HANP treatment groups compared to the m-HAP and control groups (Fig. 6a). To identify the role of HIF-1α in the osteogenic differentiation of hWJ-MSCs, we used 2-MeOE2, which is a specific HIF-1α inhibitor, was used. The decreased concentration of HIF-1α paralleled the impaired mineralized matrix deposition and ALP activity in these hWJ-MSCs, indicating that HANPs can promote the HIF-1α production of HUVECs to facilitate the osteogenesis of hWJ-MSCs (Fig. 7).
To properly apply HANPs for use in bone tissue engineering, it is necessary to gain further insights into the mechanisms by which HANPs promote the osteogenic differentiation of hWJ-MSCs mediated by HUVECs. The ERK1/2 pathway is downstream of HIF-1α [36] and is fundamental to the differentiation of MSCs [37]. In this work, the concentrations of HIF-1α in the CM coincide well with the p-ERK1/2 levels in the hWJ-MSCs (Fig. 6b, c). When 2-MeOE2 was applied, the p-ERK1/2 expression in the hWJ-MSCs failed to be activated, indicating that HIF-1α functioned upstream of ERK1/2 signaling. To directly address the role of ERK1/2 signaling in the osteogenic differentiation of hWJ-MSCs, PD98059, a specific MEK inhibitor, was used. The suppression of ERK1/2 signaling resulted in the lowest osteogenic differentiation of hWJ-MSCs. One possible reason for this occurrence is that the ERK1/2 pathway plays a key role in both HIF-1α signaling and in the apoptosis and proliferation signaling pathways, which could be responsible for the observed changes in osteogenic differentiation in these cells [38, 39]. Additionally, this could also be related to the presence of vascular endothelial growth factor (VEGF). VEGF is one of the downstream effectors of HIF-1α signaling [33], and it can also promote the osteogenic differentiation of MSCs via activation of the ERK1/2 pathway [37]. Our previous study found that np20 induced the production of VEGF in HUVECs [9]; therefore, it is possible that the suppression of the ERK1/2 pathway may result in inhibition of VEGF, which would lead to the decreased osteogenic differentiation of hWJ-MSCs. According to the available experimental results, we can summarize as follows. HANPs are able to more optimally process better direct [5] and indirect osteoinductive effects than m-HAPs. Compared to autogenous bone grafts and bone allografts, there is an extensive source of HANP and without secondary damage and potential immunogenicity. However, compared to m-HAPs, HANPs can suppress the angiogenic ability of HUVECs [9] and exhibit slight cytotoxicity in both a time- and dose-dependent fashion.
Recently, growing evidence has demonstrated the importance of HIF-1α in the bone regeneration. However, few studies have been able to quantitatively predict the MSC differentiation rate under specific initial conditions, such as the HIF-1α concentration. Taking cell proliferation, apoptosis, and osteogenic differentiation into account, we present a mathematical model that combines a two-stage cell lineage with HIF-1α that is highly correlated with our experimental data. By fitting the differentiation rate of hWJ-MSCs in 0–4000 pg/mL HIF-1α, we acquired the equations for describing the differentiation rate, HIF-1α concentration, and time. As shown in Fig. 8d, this model can depict the cell number map under different HIF-1α concentrations, so that it is possible to explore the intrinsic dynamics of the two-stage system [40]. Additionally, this model mathematically validates the effect of HIF-1α on the osteogenic differentiation of hWJ-MSCs. Moreover, based on a multi-stage cell-lineage model and logistic model, our model is sufficiently stable to enable long-term predictions without falling into the trap of population unlimited explosion [41].
By using the existing experimental data, both the cell number and differentiation rate can be predicted with a defined initial cell seeding density and HIF-1α concentration. As such, the optimum incubation time is also obtained. Consequently, we can predict the optimum concentration of HIF-1α and determine the most optimal time for osteogenesis, which is important for efficient tissue engineering. A two-stage cell-lineage model is applicable for predicting the proliferation and differentiation of stem cells, which have two cell lineages. On this basis, the model founded on the initial conditions and existing experimental data can be established to identify the optimum culture conditions in vitro, which will assist in optimizing bone repair in vivo.
Заключение
In this study, we explored the specific biological effects of HANPs on hWJ-MSCs mediated by HUVECs. Compared to m-HAPs, both np20 and np80 showed slight cytotoxicity in both a time- and dose-dependent fashion. Importantly, the size of the HANPs appeared to have no significant impact on this cytotoxicity. Our data also showed that HANPs, especially np20, were capable of facilitating HUVECs to secrete increased levels of HIF-1α, which directly correlated with the enhanced osteogenic differentiation of hWJ-MSCs via the activation of the ERK1/2 pathway (Fig. 9). More remarkably, the results from the two-stage cell-lineage model suggested that HIF-1α exerted a dose-dependent stimulatory effect on the osteogenic differentiation rate of hWJ-MSCs. Additionally, the optimum concentration of HIF-1α and incubation time were estimated based on the initial conditions using an in vitro model, which could be invaluable in the future for tissue engineering applications. Collectively, these observations provide evidence that HANPs may improve bone regeneration by modulating cell–cell interactions.

A schematic illustration of the possible mechanisms
Доступность данных и материалов
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
Сокращения
- HA:
-
Hydroxyapatite
- HANPs:
-
HA nanoparticles
- hWJ-MSCs:
-
Human umbilical cord Wharton’s jelly-derived mesenchymal stem cells
- HUVECs:
-
Human umbilical vein endothelial cell
- m-HAP:
-
Micro-sized HAP particles
- PBS:
-
Физиологический раствор с фосфатным буфером
- ECM:
-
EC medium
- 2-MeOE2:
-
2-Methoxyestradiol
- ELISA:
-
Иммуноферментный анализ
- RUNX-2:
-
Runt-related transcription factor 2
- Col I:
-
Type I collagen
- OCN:
-
Osteocalcin
- ALP:
-
Щелочная фосфатаза
- SOX 2:
-
SRY-related HMG-box 2
- ERK:
-
Extracellular signal-related kinases
- VEGF:
-
Фактор роста эндотелия сосудов
Наноматериалы
- Батареи специального назначения
- Солнечный элемент
- Анализ организации актина и фокальной адгезии в клетках U2OS на полимерных наноструктурах
- Нанотехнологии инактивируют раковые стволовые клетки
- Изучение in vitro влияния наночастиц Au на клеточные линии HT29 и SPEV
- Эффекты взаимодействия в сборке магнитных наночастиц
- Нацеливание на эндотелиальные клетки с помощью многофункциональных наночастиц GaN / Fe
- Синергетические эффекты наночастиц Ag / BiV1-xMoxO4 с повышенной фотокаталитической активностью
- Стимуляция роста клеток SH-SY5Y наночастицами золота, модифицированными 6-меркаптопурином и проникающим в нейро…
- Нанокомпозиты на основе оксида графена, украшенные наночастицами серебра в качестве антибактериального аге…