


Изучение in vitro влияния наночастиц Au на клеточные линии HT29 и SPEV
Аннотация
Модели клеточных культур - отличные инструменты для определения потенциальной токсичности наночастиц и фундаментальных исследований рака. Таким образом, информация о потенциальной токсичности AuNP и воздействии на здоровье человека необходима для использования наноматериалов в клинических условиях. Цель нашего исследования - изучить эффекты AuNP на клеточные линии эпителиального происхождения:непрерывные и онкогенные. Использовали инокулированную эмбриональным эпителием почки свиньи (SPEV) линию клеток и линию клеток колоректальной карциномы (HT29). В тестовых культурах оценивали пролиферацию клеток, некроз / апоптоз и образование многоклеточных сфероидов. Мы продемонстрировали, что концентрации AuNP 6–12 мкг / мл снижают пролиферацию клеток SPEV и HT29 и увеличивают количество клеток на ранних и поздних стадиях апоптоза и некроза. Было показано, что небольшие концентрации AuNP (1–3 мкг / мл) стимулируют образование многоклеточных сфероидов клетками HT29 и SPEV. Однако более высокие концентрации AuNP (6–12 мкг / мл) оказывали как цитотоксическое, так и анти-когезионное действие на клетки в суспензии. Большую чувствительность к действию AuNP показала линия HT29 (6 мкг / мл) по сравнению с клетками SPEV (12 мкг / мл). Это экспериментальное исследование влияния AuNP на клеточные линии SPEV и HT29 оправдывает их дальнейшее применение в лечении противоопухолевых заболеваний, опосредованных AuNP.
Фон
Производство и исследование наночастиц золота (AuNPs) имеют не только высокий потенциал для широкого терапевтического применения золота [1, 2], но также сделали их пригодными для конкретных биомедицинских приложений, таких как таргетная терапия [3, 4]. Недавние сообщения показали, что использование AuNP дает возможность для новых противоопухолевых методов лечения с пониженным риском развития резистентности. Так, несколько исследований доказали противоопухолевую активность наночастиц против рака груди, печени, желудка, толстой кишки, легких [5, 6].
Известно, что наночастицы (НЧ) могут модулировать судьбу клетки, индуцировать или предотвращать мутации, инициировать межклеточную коммуникацию и модулировать клеточную структуру [7, 8]. Кроме того, AuNP имеют преимущества перед другими металлическими NP из-за их биосовместимости и противоопухолевой активности [8,9,10,11,12]. Цитотоксические и генотоксические эффекты AuNP связаны с их формой, размером, зарядом, концентрацией, временем взаимодействия и т. Д. [12,13,14]. Таким образом, информация об их потенциальной токсичности и воздействии на здоровье человека необходима для использования наноматериалов в клинических условиях.
В настоящее время, несмотря на большие успехи таргетной терапии, проблема избирательной доставки AuNP в ткани-мишени остается нерешенной. В некоторых исследованиях отмечена разная скорость захвата НЧ эпителиальными клетками разного происхождения [15, 16]. Тем не менее, исследований, объясняющих этот феномен, недостаточно, хотя они могут помочь в достижении ткане-селективного нацеливания на AuNPs. Анатомические или физиологические различия между разными эпителиями могут объяснить различия в скорости захвата и транспорта AuNP. В частности, на скорость поглощения могут влиять свойства плазматической мембраны клеток и связывание наночастиц с гликопротеинами и протеогликанами клеточной поверхности, а также способность клеток к везикулярному транспорту [17]. Таким образом, учитывая невозможность исключительно избирательного взаимодействия наночастиц с клетками-мишенями, актуальным является сравнительное изучение особенностей их взаимодействия с нормальными и онкогенными клетками во избежание нежелательных последствий терапии рака [8,9,10].
Хотя модели in vivo ценны для оценки биологической токсичности наночастиц, модели клеточных культур очень полезны для доклинических физиологических и токсикологических исследований. В настоящее время клеточные культуры широко используются в различных областях биологии, медицины, ветеринарии и биотехнологии. Использование клеточных культур позволяет исследовать биологические процессы, которые сложно, а иногда и невозможно, изучать на уровне организмов. Важную роль клеточных культур играет биотехнология в производстве многих вакцин, тест-систем и биологически активных веществ. Клеточные культуры используются для диагностики заболеваний различной этиологии, в качестве тест-объектов при тестировании новых фармакологических, лечебных и косметических средств, а также пищевых добавок [18].
В данной работе на моделях клеточных культур мы попытались изучить особенности действия AuNP на эпителиальные клетки непрерывного и онкогенного происхождения клеточных линий. Монослойная культура эпителиальных клеток SPEV (инокулированная линия эмбрионального эпителия почек свиньи) и клеток HT-29 (линия клеток карциномы толстой кишки) может рассматриваться как модель нормальных и раковых эпителиальных тканей при применении противоопухолевой терапии с помощью AuNP. Несколько традиционных анализов цитотоксичности, включая адгезию, пролиферацию, некроз / апоптоз и многоклеточные сфероиды, были использованы для подтверждения клеточной цитотоксичности AuNP.
Методы
Культура клеток SPEV
Клетки SPEV культивировали в пластиковых колбах в среде DMEM (Sigma, США) с 5% FCS ( v / v ) (HyClone, США) с добавлением пенициллина / стрептомицина (PAA, Австрия) и амфотерицина B (5 мкг / мл) (5% CO 2 , Влажность 95%) по данным [19]. Концентрация посева составляла 0,5–2 × 10 4 . клеток / см 2 . Питательную среду заменяли каждые 2 дня. Клетки пассировали при 100% слиянии [20]. Клеточная линия SPEV росла и сохраняла первоначальную морфологическую структуру монослоя во время серийных пассажей без признаков дегенерации клеток в культуре.
Культура клеток HT29
Клетки HT29 культивировали в пластиковых колбах (Nunc, Дания) в среде RPMI-1640 (Sigma, США) с 10% FCS ( v / v ) (HyClone, США) с добавлением 2 мМ L-глутамина (Sigma, США) и 40 мг / мл гентамицина (Sigma, США) в стандартных условиях (5% CO 2 , Влажность 95%) [21]. Оптимальная плотность клеток составляла 0,5–4,0 × 10 4 . клеток / см 2 . Клетки нам любезно предоставил Банк линий клеток из тканей человека и животных Института экспериментальной патологии, онкологии и радиобиологии им. Р.Е. Кавецкого НАН Украины.
Манипуляции с AuNP
AuNP были любезно предоставлены Институтом биохимии и физиологии растений и микроорганизмов РАН. AuNP были синтезированы цитратным методом [22]. Средний размер наночастиц составлял 15 нм. Начальная концентрация золота составляла 57 мкг / мл. Результаты темнопольной электронной микроскопии, изображения AuNP 15 нм и спектры экстинкции AuNP 15 нм (b) показаны на рис. 1 (рис. 1a; примечание –– диаграмма распределения по размерам) [23]. AuNP вводили в клетки путем пассивной диффузии при 37 ° C. Исследуемые концентрации составляли 1, 3, 6 и 12 мкг / мл. Клетки без AuNP в тех же условиях были взяты за контроль.
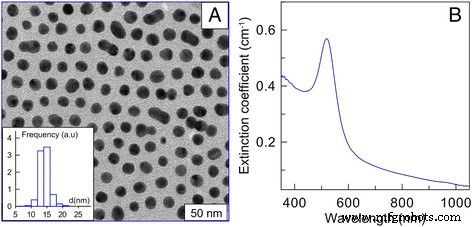
Результаты темнопольной электронной микроскопии, а изображение AuNP 15 нм (примечание –– диаграмма распределения по размерам), б Спектры экстинкции AuNP 15 нм
Клетки адгезии и пролиферации
О морфофункциональном состоянии культур клеток судили по адгезивным свойствам и пролиферативной активности. Адгезивные свойства клеток SPEV и HT29 оценивали визуально с помощью инвертированного микроскопа; количество прилипших и уплощенных клеток подсчитывали через 30, 60, 120, 180 и 1440 минут после начала культивирования.
Динамику пролиферации клеток SPEV и HT29 изучали в течение 1–4 суток. Для проверки прироста количества клеток в исследуемых культурах к условиям исследования их ферментативно (1:1 (0,25% раствор трипсина:ЭДТА, ПАК)) отделяли от пластика и подсчитывали количество клеток. Общее количество культивированных клеток подсчитывали традиционным методом в камере Горяева.
Апоптотические / некротические процессы
Апоптотические и некротические процессы в клетках SPEV и HT29, подвергнутых действию AuNP, исследовали через 4 дня с красителями Аннексин-V (BD, США), 7-амино-актиномицин (7AAD) (BD) на приборе FACS Calibur Becton-Dickinson. Результаты были проанализированы с помощью программного обеспечения WinMDI v.2.8.
Многоклеточные сфероиды
Многоклеточные сфероиды (МС) были созданы для оценки in vitro воздействия AuNP на миграцию и агрегационный потенциал исследуемых клеток. Сфероидная (3-D) модельная система клеток SPEV и HT29 культивировалась обычным методом, описанным в [24] и модифицированным в нашей лаборатории [25]. Вкратце, суспензия клеток подсчитывалась с использованием трипанового синего и равного количества клеток (5 × 10 4 клеток / см 2 ) были посажены в полностью дополненную питательную среду. Генерация МС была достигнута обработкой клеточной культуры с 0,24% карбоксиметилцеллюлозы (КМЦ) в 24-луночных планшетах, покрытых 1% агаром, с вращением (80 об / мин) в течение 24 часов. После этого трехмерную культуру клеток поддерживали в стандартных условиях. Для исследования зависимости размера и количества МС от концентрации AuNP были созданы МС в присутствии AuNP. Дальнейшее культивирование проводили в течение 48 ч при постоянном вращении пластин. На следующем этапе были получены микрофотографии методом темного поля на микроскопе Carl Zeiss Stemi 2000. Морфологию МС изучали с помощью программы Axio Vision Release 4.7 (Zeiss). Эта программа позволяет измерять геометрические размеры клеточных агрегатов. Затем был рассчитан объем всех МС, которые были в файлах. Он использовался по следующей формуле: V =0,4 × а × b 2 , где a и b –– геометрические диаметры МС [24]. Для статистического анализа все агрегаты клеток были сгруппированы по размеру от 1 × 10 −4 мм 3 до 1 × 10 −2 мм 3 с шагом 1 × 10 −3 мм 3 . Количество РС и медиана объемов РС были оценены для каждой группы.
Статистический анализ
Однофакторный дисперсионный анализ и t Стьюдента test использовались для статистической обработки данных с помощью пакета программ Statistica 8. Порог значимости составил 0,05. Результаты представлены в виде средних значений и стандартных ошибок (M ± SE).
Результаты
Влияние AuNP на адгезию клеток SPEV и HT-29
Адгезия клеток является показателем функционального состояния клеток и необходима для дальнейшего роста культуры. Когда адгезия прекращалась, клетки становились уплощенными и приобретали соответствующую морфологию. Адгезионные свойства клеток SPEV представлены на рис. 2.
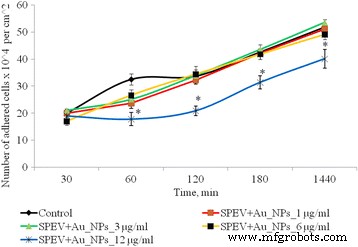
Динамика адгезии клеток SPEV после воздействия AuNP, * p ≤ 0,05 значимо по сравнению с контролем
После 1 ч культивирования клеток SPEV с AuNP в концентрации 1, 3 и 6 мкг / мл количество прикрепившихся клеток было ниже по сравнению с контрольным значением. Процент уплощенных клеток в образцах с AuNP для этих концентраций достоверно не отличался от контроля. Адгезия замедлялась после 1 ч инкубации с AuNP в концентрации 12 мкг / мл. Количество прикрепившихся клеток на квадратный сантиметр уменьшилось в 1,8 раза по сравнению с контролем. Эта тенденция к адгезии сохранялась на протяжении всех периодов испытаний. Через 24 ч наблюдения количество прилипших клеток было меньше контроля в 1,3 раза. В то же время инкубация AuNP в малых концентрациях (1 и 3 мкг / мл с опухолевыми клетками (HT29) не оказывала существенного влияния на количество адгезивных клеток. Повышение концентрации AuNP до 6 и 12 мкг / мл приводило к снижению количество опухолевых клеток в адгезивной фракции в 1,16 и 1,28 раза соответственно (рис. 3). На полученные данные могут влиять несколько процессов. Одним из них является цитостатическое / цитотоксическое действие AuNP на фракцию адгезии как опухоли, так и линии эмбриональных клеток, что приводит к гибели клеток, переходу к апоптозу или некрозу. Другой процесс - снижение адгезии клеток под влиянием AuNP и перенос клеток во фракцию суспензии. Примечательно, что оба процесса могут осуществляться одновременно, и каждый из них может способствовать уменьшению количества живых клеток во фракции адгезии.
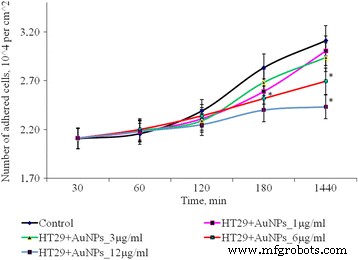
Динамика адгезии клеток HT29 после воздействия AuNP, * p ≤ 0,05 значимо по сравнению с контролем
Влияние AuNP на пролиферацию клеток SPEV и HT-29
Изучено влияние AuNP в диапазоне концентраций 1–12 мкг / мл на пролиферативные процессы в культуре клеток SPEV (рис. 4). На 2–4 сутки культивирования с AuNP в концентрации 1, 3 и 6 мкг / мл количество клеток достоверно не отличалось от контроля. На 4-е сутки культивирования с AuNP в концентрации 3 и 6 мкг / мл этот показатель снизился в 1,15 и 1,23 раза соответственно по сравнению с контролем. Уменьшение количества клеток в 1,5 раза (дни 2 и 3) и в 1,15 раза на день 4 культивирования с AuNPs в концентрации 12 мкг / мл наблюдалось в культуре SPEV по сравнению с контролем. Таким образом, концентрация AuNP, составляющая 12 мкг / мл, замедляла рост клеток в течение наблюдаемого периода времени.
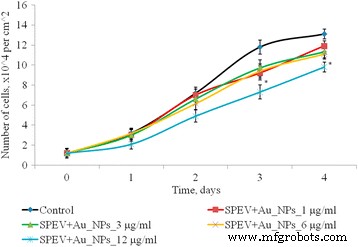
Пролиферация клеток SPEV после воздействия AuNP, * p ≤ 0,05 значимо по сравнению с контролем
Влияние AuNP в концентрациях от 1 до 12 мкг / мл на количество клеток HT 29 в монослойной культуре показано на рис. 5. В течение первых 3 дней инкубации количество клеток в контроле и в присутствии AuNPs статистически не различались. На 4-е сутки культивирования отмечалось дозозависимое уменьшение количества клеток в 2D-культуре. Так, после 4 дней культивирования при низких концентрациях AuNP (1 и 3 мкг / мл) количество клеток HT 29 существенно не отличается по сравнению с контролем. Но при более высоких концентрациях AuNP (6–12 мкг / мл) количество клеток HT29 было ниже, чем в контроле, в 1,33 и 1,44 раза соответственно.
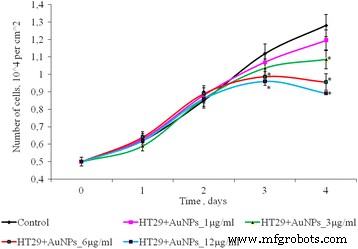
Пролиферация клеток HT29 после воздействия AuNP, * p ≤ 0,05 значимо по сравнению с контролем
Влияние AuNP на апоптотические / некротические процессы в клетках SPEV и HT-29
Клетки SPEV и HT-29 в присутствии AuNP культивировали в течение 4 дней в стандартных условиях. Культивирование клеток SPEV и HT29 с AuNP в концентрации 1 и 3 мкг / мл и индексы апоптотических / некротических процессов существенно не отличались от контроля (таблицы 1 и 2).
Культивирование с AuNP в концентрации 6–12 мкг / мл увеличивало процентное содержание клеток аннексина V + / 7AAD +, аннексина V- / 7AAD + и аннексина V + / 7AAD, а также снижало процент живых клеток. Количество Аннексина V + / 7AAD + SPEV клеток был выше контрольного значения на 7,8 ± 0,7% ( p ≤ 0,05) с 12 мкг / мл AuNP. Количество Аннексина V + / 7AAD + Клеток HT 29 было выше контрольного значения на 3,2 ± 0,4% ( p ≤ 0,05) с 6 мкг / мл AuNP и на 4,8 ± 0,6% ( p ≤ 0,05) с 12 мкг / мл AuNP.
Влияние AuNP на создание многоклеточных сфероидов из клеток SPEV и HT29
Чтобы определить зависимость размера и количества многоклеточных сфероидов (МС) от концентрации AuNP, были созданы МС при различных концентрациях AuNP в течение 48 часов. Наши данные продемонстрировали разнообразную способность клеток HT29 и SPEV образовывать многоклеточные сфероиды в одних и тех же условиях микросреды (рис. 6 и 7).
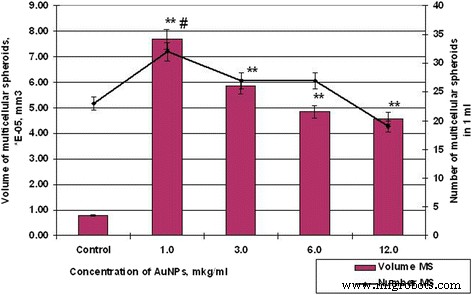
Количество и объем клеток MS SPEV после инкубации с AuNP, # p ≤ 0,01 (для количества МС); ** p ≤ 0,01 (для объема МС)
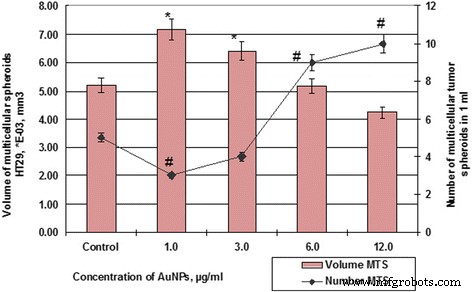
Количество и объем клеток MS HT29 после инкубации с AuNP. # p ≤ 0,01 (для количества МС); ** p ≤ 0,01 (для объема МС)
Так, если контрольные образцы клеток HT29 в течение 48 ч образовывали сфероиды в среднем объеме 5,19 × 10 −3 мм 3 , средний сфероид объема SPEV-клеток составлял 0,79 × 10 −5 мм 3 . В то же время влияние AuNP на клетки HT29 и SPEV имело ту же тенденцию. Присутствие AuNP в клеточном микроокружении стимулировало образование многоклеточных сфероидов в обеих культурах. Таким образом, когда концентрация AuNP составляла 1 и 3 мкг / мл, объем MS для SPEV увеличивался в 9,7 и 7,4 раза соответственно по сравнению с контролем (рис.6), те же концентрации AuNP также стимулировали увеличение объема MS для HT29 в 1,4 и 1,2 раза соответственно (рис. 7).
Дальнейшее увеличение концентрации AuNP привело к снижению среднего объема МС в обеих культурах. Повышение концентрации AuNP с 1 до 12 мкг / мл уменьшило объем МС HT29 с 7,18 × 10 −3 мм 3 до 4,24 × 10 −3 , в 1,69 раза, по данным контроля. Что касается SPEV, при увеличении концентрации AuNP с 1 до 12 мкг / мл объем MS уменьшился с 7,69 до 4,58 × 10 −5 мм 3 , в 1,68 раза, по данным контроля. Однако увеличение концентрации AuNP совпадает с уменьшением объема MS и коррелирует с увеличением количества сфероидов в культуре клеток HT29 (рис. 6 и 7). Количество МС HT29 увеличивалось с 3 до 10 на поле зрения при концентрации AuNP от 1 до 12 мкг / мл. В то же время количество MS SPEV уменьшилось с 32 до 19 соответственно.
Полученные данные (рис. 6 и 7) демонстрируют, что AuNP способны влиять на когезионные взаимодействия в межклеточной системе. Наши данные показывают, что небольшие концентрации AuNP (1–3 мкг / мл) стимулировали образование многоклеточных сфероидов как эмбриональных, так и опухолевых клеток. Однако более высокие концентрации AuNP (6–12 мкг / мл) оказывали как цитотоксическое, так и анти-когезионное действие на клетки в суспензии. Этот процесс способствовал формированию большего количества МС HT29 с уменьшенным средним объемом. Что касается SPEV, высокая концентрация AuNP может иметь цитостатический эффект, который снижает количество клеток в адгезивной фракции и количество MS в суспензии. Ранее авторы сообщали, что углеродные наночастицы уменьшают адгезию клеток к субстрату, стимулируют перенос клеток в суспензию и приводят к образованию многоклеточных сфероидов [25, 26]. В литературе есть данные о способности AuNP разрушать структуру актин / миозиновых микрофиламентов и снижать пролиферацию, адгезию и дифференцировку клеток [27]. Наши данные подтвердили это предположение.
Обсуждение
Мы оценили влияние AuNP на пролиферацию, некроз / апоптоз и образование многоклеточных сфероидов эпителиальных клеток непрерывного и онкогенного происхождения клеточной линии. Было показано, что AuNPs в концентрации 6–12 мкг / мл снижали количество клеток SPEV и HT29 и увеличивали количество клеток на ранних и поздних стадиях апоптоза и некроза. Небольшие концентрации AuNP стимулируют образование многоклеточных сфероидов клетками HT29 и SPEV. Однако более высокие концентрации AuNP оказывали как цитотоксическое, так и анти-когезионное действие на клетки в суспензии. Большую чувствительность к действию AuNP продемонстрировала линия HT29 (6 мкг / мл) по сравнению с клетками SPEV (12 мкг / мл)
Эффекты AuNPs на клеточную морфологию и цитоскелет только недавно привлекли больше внимания, а лежащий в основе механизм и предстоящие последствия не были изучены глубоко [28,29,30]. В связи с этим для всех новых типов AuNP важно оценивать их путь поглощения эндоцитами и внутриклеточную локализацию как функцию времени. Для различных типов AuNP было описано, что эффекты зависят от внутриклеточной концентрации AuNP и являются временными, когда после повторяющихся делений клеток внутриклеточные концентрации AuNP экспоненциально снижаются, и эффекты больше не наблюдаются. Также необходимо оценить возможное эндосомное ускользание AuNP. Поскольку было описано, что дефекты цитоскелета явно зависят от концентраций AuNP, следует тестировать широкий диапазон концентраций частиц, чтобы попытаться оценить максимальную нагрузочную способность клетки без каких-либо эффектов. Кроме того, поскольку цитоскелет также участвует во многих внутриклеточных сигнальных путях, остается исследовать, приводит ли индуцированное AuNPs нарушение цитоскелета к вторичным эффектам [31].
Поскольку НЧ имеют определенные физические размеры, занимаемый ими внутриклеточный объем может приводить к изменениям клеточной морфологии или влиять на структуру сети клеточного цитоскелета [28, 29, 31]. Последующие эффекты также могут быть связаны с высокими требованиями позы NP к клеточному эндоцитозу. Было описано, что AuNP оказывают сильное влияние на морфологию нескольких типов клеток, таких как клетки легкого карциномы человека A549 [32]. Также было описано, что AuNPs оказывают зависимое от концентрации действие на актиновые фибриллы фибробластов кожи человека [33, 34]. Миронова и др. [35, 36] далее показали, что филаменты цитоскелета разрушаются в зависимости от времени воздействия AuNP, концентрации и размера NP, хотя на уровни экспрессии белка актина или β-тубулина это не влияет.
Используемый тип клеток также имеет большое значение, поскольку разные типы клеток, даже если они тесно связаны, могут совершенно по-разному реагировать на один и тот же тип наноматериалов [37, 38]. Предпочтительно, следует тестировать те типы клеток, которые наиболее вовлечены в (будущее) биомедицинское применение НЧ (например, эпителиальные, эндотелиальные клетки) или множественные клетки, происходящие из разных зародышевых листков. При исследовании цитотоксических эффектов следует минимизировать использование типов раковых клеток, так как это может привести к ошибочным результатам [39]. Раковые клетки обладают несколькими специфическими характеристиками и измененными внутриклеточными сигнальными путями, которые предназначены для усиления пролиферации и поддержания жизнеспособности клеток, что сделает их менее склонными к некоторым цитотоксическим эффектам, опосредованным NP.
По нашему мнению, связывание AuNP с поверхностными функциональными группами (например, трансмембранными белками) клеток может быть обратимым или необратимым, приводя к временным или постоянным структурным повреждениям [40, 41]. Возможные последствия изменений биомеханических свойств (например, твердости и эластичности), адгезии и электрических свойств поверхности клеток очевидны. Таким образом, изменения твердости или эластичности могут влиять на структурную гибкость поверхности, выработку механической энергии для деления клеток и подвижность клеток. Что касается адгезии, клеточное микроокружение обычно состоит из внеклеточного матрикса со специфическими молекулами, которые позволяют клеткам прикрепляться к своему окружению [42]. Поверхностный заряд, несомненно, играет важную роль во взаимодействии между клетками и их окружением.
Другие авторы также сообщили о том, что НЧ преимущественно локализуются в митохондриях и вызывают окислительный стресс, а также потенцируют структурные повреждения [40]. Недавняя статья Pan et al. описывает, что 1,4 нм AuNPs вызывают некроз через окислительный стресс и повреждение митохондрий в клетках Hela [43]. Накопление наночастиц в клеточной среде при биодеградации небезопасно, поскольку может разрушить органеллы и даже вызвать генетические мутации.
Изменения, происходящие в клетках при апоптозе, аналогичны для большинства типов клеток. В апоптозных клетках происходят изменения липидного состава плазматической мембраны:фосфатидилсерин переносится из цитоплазматической части бислоя на внешнюю сторону, вызывая активацию каспазного каскада, конденсацию хроматина и нарушение цепи транспорта электронов в митохондриях и, в конечном итоге, задержку синтеза АТФ. Запрограммированная гибель клеток может быть вызвана рецепторно-опосредованными физиологическими стимулами, вызванными генетическими нарушениями, воздействием химических или физических факторов, а также другими изменениями в клетках. Мы наблюдали этот эффект с 6–12 мкг / мл AuNP.
Многоклеточные агрегаты (сфероиды, эмбриоидные тела) представляют собой прерывистый уровень между растущими клетками монослоя и культурой ткани. Сфероиды являются объективной моделью трехмерного роста и организации клеток, межклеточного взаимодействия и влияния условий микроокружения, например концентрации AuNP, на интенсивность пролиферации, а также на адгезию клеток и образование микроагрегатов. Формирование МС - хорошо зарекомендовавший себя метод культивирования как опухолевых, так и эмбриональных клеточных линий [24, 44, 45]. В нашей работе образование и рост сфероидов достигается за счет добавления КМЦ как части искусственного внеклеточного матрикса и покрытия поверхности 1% агаром, который ингибирует адгезию клеток к поверхности и стимулирует агрегацию клеток. В этих условиях культивирование РС может осуществляться до тех пор, пока агрегаты не образуют центральный некроз из-за ограниченного роста клеточной массы или спонтанной дифференцировки эмбриональных клеток.
В литературе есть сведения о взаимодействии AuNP с линией клеток рака толстой кишки и линиями эмбриональных клеток [46, 47]. Согласно этим данным, воздействие даже очень низких концентраций AuNP может оказывать повреждающее действие на клетки-предшественники нервных клеток человеческого эмбриона и HT29, вызывая стресс для пролиферации, дифференцировки и апоптотической гибели клеток.
Имеются опубликованные данные о том, что эффект AuNP основан на накоплении фазы G0 / G1, истощении фазы S и G2 / M, а также на снижении уровня АТФ в клетках плоскоклеточной карциномы полости рта человека (HSC-3) [48]. Регуляция клеточного цикла может быть решена путем нарушения фокальных контактов клеток с субстратом и переноса клеток во взвешенную фракцию в 2D-культуре и ингибирования межклеточных контактов в щелевых соединениях в 3D-культуре [48,49,50]. Из-за наноразмеров AuNP (около 15 нм) они не могут быть центрами сцепления для клеток. В то же время, внедрение AuNP в клеточную мембрану [51], влияние на дзета-потенциал AuNP клеточной мембраны [32] и влияние на формирование межклеточных / межклеточных контактов, очевидно, могут запускать механизм некроза / апоптоза. , цитотоксический эффект и остановка клеточного цикла. Нарушение очаговых контактов клеток с субстратом и перенос клеток во взвешенную фракцию - способ регуляции клеточного цикла [48, 49]. Небольшие концентрации AuNP не оказывали статистически значимого цитотоксического действия на клетки. Однако более высокие концентрации AuNP оказывали как цитотоксическое, так и анти-когезионное действие на клетки в суспензии. Этот процесс способствовал формированию большего количества МС с уменьшенным средним объемом. Мы предполагаем, что AuNP вклинивается в когезионные контакты клеток и компрометирует их. Таким образом, наши эксперименты по влиянию AuNP на клеточные линии SPEV и HT29 подтверждают их дальнейшее применение в разработке методов лечения рака, опосредованных AuNP.
Хотя для подтверждения противоракового действия в исследованиях на животных in vivo потребуются будущие исследования. Тем не менее, мы глубоко убеждены в том, что, зная природу вещества и его возможное негативное влияние, мы сможем избежать пагубного воздействия AuNP и использовать их положительный биотехнологический потенциал. Наше исследование может быть достаточно надежно применено в эффективных материалах для противоракового лечения с максимальной пользой для медицины.
Выводы
Наши результаты подтверждают мнение о том, что AuNP вызывают дозозависимую цитотоксичность в клетках SPEV и HT29. Кроме того, этот отчет впервые демонстрирует, что 15 нм AuNP в концентрациях 6–12 мкг / мл снижают пролиферацию клеток SPEV и HT29 и увеличивают количество клеток на ранних и поздних стадиях апоптоза и некроза. Также было показано, что небольшие концентрации AuNP (1–3 мкг / мл) стимулируют образование многоклеточных сфероидов. Однако более высокие концентрации AuNP оказывали как цитотоксическое, так и анти-когезионное действие на клетки в суспензии. Большую чувствительность к действию AuNP продемонстрировала линия HT29 (6 мкг / мл) по сравнению с клетками SPEV (12 мкг / мл)
Наноматериалы
- Наночастицы золота для химиотерапевтических датчиков
- Многофункциональные наночастицы золота для улучшенных диагностических и терапевтических применений:обзор
- Наночастицы для терапии рака:текущий прогресс и проблемы
- Получение и магнитные свойства легированных кобальтом наночастиц шпинели FeMn2O4
- Получение наночастиц mPEG-ICA, нагруженных ICA, и их применение в лечении LPS-индуцированного повреждения клеток H9c2…
- Биобезопасность и антибактериальная способность графена и оксида графена in vitro и in vivo
- Синтез и эффективность in vitro покрытых полипирролом железо-платиновых наночастиц для фототермической терапии…
- Стимуляция роста клеток SH-SY5Y наночастицами золота, модифицированными 6-меркаптопурином и проникающим в нейро…
- Влияние упругой жесткости и поверхностной адгезии на отскок наночастиц
- Двухэтапная методология изучения влияния агрегации / агломерации наночастиц на модуль Юнга полимерных нанок…