


Нанотехнологии инактивируют раковые стволовые клетки
Аннотация
Одна из задач современной онкологии - идентификация раковых стволовых клеток и поиск терапевтических средств, способных к их специфическому подавлению. В статье представлены данные о фенотипических характеристиках клеток карциномы Эрлиха как удобной и понятной модели роста опухоли. Доказательства наличия раковых стволовых клеток как части карциномы Эрлиха и значения CD44 + и CD44 - были продемонстрированы субпопуляции в поддержании роста этого типа опухоли. Высокая (десятикратная) канцерогенная активность карциномы Эрлиха CD44 + клетки по сравнению с CD44 - клеток было доказано. В этой паре сравнения CD44 + клетки имели более высокий потенциал генерирования в брюшной полости CD44 high , CD44 + CD24 - , CD44 + CD24 + субпопуляции клеток, подчеркивая присутствие раковых стволовых клеток в пуле CD44 + ячеек.
В данном исследовании показана способность синтезированных гибридных нанокомплексов, содержащих наночастицы ортованадатов редкоземельных элементов GdYVO 4 :Eu 3+ и холестерин для подавления роста опухоли и увеличения выживаемости животных с опухолями. Особый вклад в опухолевый эффект вносит каждый из его компонентов. Обработка клеток карциномы Эрлиха двухкомпонентным гибридным комплексом привела к максимальному снижению концентрации наиболее канцерогенного CD44 high клеток с одновременным увеличением количества CD117 + клетки, снизившие интенсивность роста опухоли на 74,70 ± 4,38% по сравнению с контролем.
Фон
Проблема злокачественного новообразования остается одной из самых актуальных в медицине. В последние десятилетия был достигнут некоторый прогресс в разработке новых методов лечения рака. Это связано с пересмотром классической концепции рака и открытием раковых стволовых клеток (РСК), которые способны к неограниченному самообновлению и могут быть идентифицированы по ряду фенотипических маркеров. Большинство этих клеток устойчивы к радио- и химиотерапии, вызывая рецидив злокачественного роста и метастазирование. Освоены новые методы противоопухолевой терапии, а именно, позволяющие избирательно инактивировать опухолевые клетки с минимальным повреждением нормальных тканей [1].
РСК были впервые идентифицированы и описаны в 1997 г. группой М. Дика [2]. Авторы исследовали острый миелоидный лейкоз, при котором субпопуляция, составляющая 0,01–1% от общей популяции клеток, могла вызывать лейкемию при трансплантации мышам с иммунодефицитом NOD / SCID (комбинированная иммунодефицитность без ожирения и тяжелого диабета). Эти опухолевые клетки были фенотипически охарактеризованы как CD34 + CD38 - . В 2003 году М. Аль-Хадж и М.С. Wicha удалось идентифицировать РСК в твердой форме рака груди (РМЖ) человека [3]. Было обнаружено, что неразделенная популяция первичного рака молочной железы проявляла канцерогенный потенциал в 100% случаев (10/10) при введении мышам NOD / SCID в концентрации 5 × 10 4 клеток / мышь. Снижение концентрации вводимых клеток до 1 × 10 4 клетки / мышь снижали свою онкогенную активность в 4 раза (3/12) [3]. CD24 + CD44 + фракция при введении в различных дозах (2 × 10 4 до 100 клеток / мышь) не допускали роста опухоли. При этом CD44 + CD24 - / low субпопуляция обладала значительно более высокой онкогенной активностью, демонстрируя образование опухолей в 100% случаев при введении 10 3 клеток / мышь. Наиболее выраженная способность образовывать опухоли была присуща субпопуляции CD44 + CD24 - / lo ЕКА + фенотип. Введение мышам всего 200 из этих клеток привело к образованию солидных опухолей в 100% (4/4) через 5 месяцев после их инъекции [3]. Эти исследования были продолжены Ponti D. et al., Которые показали способность определенных популяций образцов биопсии рака груди образовывать маммосферы in vitro в бессывороточной культуре [4]. Большинство клеток полученных маммосфер имели CD44 + / CD24 - / low фенотип, а также повышенный канцерогенный потенциал in vivo при введении мышам SCID (тяжелый комбинированный иммунодефицит). Способность образовывать опухоль в этой субпопуляции была в 1000 раз выше, чем у традиционно пересаживаемой линии карциномы молочной железы MCF7 [4]. Однако авторы показали, что только 20% CD44 + CD24 - / low клетки обладают способностью к самообновлению. Это может быть связано с гетерогенностью данной субпопуляции, а именно с наличием дополнительных маркеров (ESA, ALDH), определяющих функцию клеток, а также может быть связано со скоростью экспрессии CD44. Статьи, опубликованные в течение последних нескольких лет, показывают, что CSC с высокой экспрессией маркера (CD44 high ) обладают наибольшей канцерогенной активностью [5, 6]. При ортотопической имплантации 5 × 10 5 CD44 высокий -RAS-преобразованный и CD44 низкий клеток к мышам NOD / SCID, было обнаружено, что субпопуляция CD44 low обладали низкой онкогенностью (опухоль образовывалась в 30% случаев), а CD44 высокой клетки были способны образовывать опухоли в 100% случаев [6].
Обобщая опубликованные данные, дифференцирующий ряд субпопуляций клеток рака груди можно представить следующим образом:
$$ \ mathrm {C} \ mathrm {D} {44} ^ {\ mathrm {high}} \ to \ mathrm {C} \ mathrm {D} {44} ^ {+} \ mathrm {C} \ mathrm { D} {24} ^ {\ hbox {-}} \ to \ kern0.5em \ mathrm {C} \ mathrm {D} {44} ^ {+} \ mathrm {C} \ mathrm {D} {24} ^ {+} \ to \ mathrm {C} \ mathrm {D} {44} ^ {\ hbox {-}} \ mathrm {C} \ mathrm {D} {24} ^ {+} $$Ряд клеток, несущих другие маркеры, в частности Sca-1 + утверждает стадию CSCs. Данные о снижении роста опухоли у мышей с нокаутом Sca-1 свидетельствуют в пользу гипотезы о инициирующей опухоль роли Sca-1 + клетки на ранней стадии онкогенеза [7]. В последнее время гораздо больше внимания исследователей привлекают не только РСК, но и клетки, составляющие их вспомогательно-регуляторное микроокружение. CD117 + Особого внимания среди них заслуживают клетки, традиционно выявляемые в пуле стволовых клеток крови [8]. Общая популяция клеток карциномы молочной железы человека включает так называемые фибробласты стромы, ассоциированные с карциномой, с CD117 + фенотип. Они поддерживают рост опухоли, способствуя ее ангиогенезу [9, 10]. Предположение о наличии в популяции карциномы Эрлиха (ЭК) стволовых клеток, исследование канцерогенного потенциала CD44 + фракция и роль CD117 + клеток, поддерживающих развитие опухоли, требует дополнительных доказательств.
Большинство экспериментов по пассированию CSC in vivo было проведено на мышах SCID или NOD / SCID. Эти мыши не реагируют иммунной реакцией на ксенотрансплантацию клеток человека. Поиск адекватных и актуальных экспериментальных моделей для изучения и оценки противоопухолевой активности различных терапевтических агентов продолжается. Одним из них является трансплантированная in vivo линия опухолевых клеток ЭК, полученная при спонтанном раке груди у мышей [11]. Однако практически отсутствуют публикации о субпопуляционном составе клеток ЭК и их фенотипических характеристиках, наличии РСК и их важности в поддержании роста этого типа опухоли. Принимая во внимание гистогенетическое сходство EC и BC, можно предположить, что в инициации и развитии моделируемой опухоли могут участвовать одни и те же гены, контролирующие пролиферацию раковых клеток, а также аналогичные биохимические пути, ведущие к экспрессии белков-маркеров опухоли. Однако предположение о наличии РСК в популяции ЭК и изучение их канцерогенного потенциала требует дополнительных доказательств, что и было одной из целей настоящего исследования.
Не менее актуальной задачей современной онкологии является поиск препаратов, не только специфически распознающих, но и инактивирующих РСК. Сама концепция понимания проблемы послужила основой, сформированной в направлении момента «тераностика» (терапия + диагностика) [12]. В рамках тераностики разработаны технологические подходы использования лекарственных средств и средства одновременной диагностики и терапии рака. Одно из направлений тераностики - нацеливание наночастиц золота на место опухоли и последующая фототермическая терапия [13]. Другой подход к идентификации опухолевых клеток - использование квантовых точек, мощных оптически контрастных агентов, позволяющих контролировать опухоль in vivo [14]. Способность квантовых точек неинвазивно визуализировать человеческие эмбриональные стволовые клетки in vivo свидетельствует в пользу их возможного биомедицинского применения [15].
В последнее время все большее внимание уделяется нанолюминофорам на основе диэлектрических материалов и широкозонных полупроводников, активированных редкоземельными элементами, а именно наночастиц (НЧ) редкоземельных металлов (в частности, ванадия и его соединений) [16]. Эти материалы обладают высокой фотостабильностью, большим стоксовым сдвигом люминесценции, отсутствием сцинтилляционного эффекта и стабильностью характерных узких полос люминесценции. При этом известно противоопухолевое действие соединений ванадия. Итак, было показано, что дихлорид ванадия может значительно ингибировать пролиферацию клеток в результате накопления в ядерном гетерохроматине с последующей индукцией митотической аберрации, временным подавлением митозов, что приводит к накоплению клеток в поздних S и G 2 фазы [17]. Перспективным для лечения злокачественных опухолей может быть использование гибридных нанокомплексов на основе НЧ РЗЭ ортованадатов GdYVO 4 :Eu 3+ и холестерин, разработанные в Институте сцинтилляционных материалов НАН Украины [18].
Целью их создания было усиление терапевтического действия противоопухолевых средств за счет наличия в составе нанокомплексов, обладающих сродством к мембранам клеток-мишеней. Один из них - холестерин, который активно «выводится» из кровотока размножающимися раковыми клетками для создания биомембран. Этому способствует наличие на поверхности большого количества опухолевых клеток SR-B1 (рецептор скавенджера, класс B тип I) и рецепторов кавеолина-1 (Cav-1), которые могут связываться со свободным холестерином кровотока [19] .
Таким образом, целью данной работы было определение субпопуляционного состава клеток ЭК, в том числе с признаками РСК, а также их онкогенной активности после предварительной обработки гибридными нанокомплексами.
Методы
Эксперименты проводились на 8-месячных самках мышей Balb / C. Мышей содержали в стандартных условиях вивария (комнатная температура 20 ± 2 ° С, относительная влажность 50–70%, цикл свет-темнота 12:12 ч). Все протоколы экспериментов одобрены Комитетом по этике животных Института проблем криобиологии и криомедицины Национальной академии наук Украины, Харьков, Украина (рекомендация № 1 от 23.01.2017) и соответствуют Европейской конвенции о защите животных. Использование экспериментальных животных (Страсбург, 1986), одобренное Первым национальным конгрессом Украины по биоэтике (Киев, 2004).
Культивирование клеток EC in vivo
Клетки карциномы Эрлиха (ЕС) пассировали в брюшной полости (ПК) мышей Balb / C. Криоконсервированные в асцитической жидкости ЕС-клетки использовали в качестве первичной культуры [20]. После оттаивания клетки ЭК были трижды повторно трансплантированы in vivo для смягчения влияния факторов замораживания-оттаивания и приобретения ими морфологических и функциональных особенностей нативных клеток [21]. «Стабилизированные» таким образом ЕС-клетки вводили внутрибрюшинно в дозе 3 × 10 6 . клеток / мышь в 0,3 мл физиологического раствора и культивировали в течение 7 дней in vivo. Через 7 дней экспериментальные животные были выведены из эксперимента под легким эфирным наркозом. Асцитную жидкость из ПК отбирали шприцем через иглу с внутренним диаметром 2,69 мм и помещали в мерную пробирку на 10 мл. Абсолютное количество клеток определяли путем увеличения объема накопленной в полости брюшины асцитической жидкости (мл) с количеством клеток ЭК, подсчитываемых в камере Горяева. Увеличение общего количества до 35,00 × 10 7 ЭК-клетки в ПК мышей к 7-м суткам были критерием развития карциномы [21]. В дальнейшем именно клетки послужили объектом изучения.
Фенотипическая оценка субпопуляций ЭК
Его проводили на проточном цитометре «FACS Calibur» («Becton Dickinson», США) с использованием моноклональных антител (US «BD Biosciences») к CD44 (FITC) (№ 553133, клон IM7), CD117 (FITC) (№ 553354, клон 2B8) и Sca-1 (FITC) № 553333, клон E13-161.7) и CD24 (PE) № 553262, клон M1 / 69) в соответствии с инструкциями производителя. В качестве контроля образцы с добавлением неиммунных FITC- и PE-меченных моноклональных антител тех же изотипов («BD Biosciences»), № 553988, клон A95-1 и № 553989, клон A95-1), в качестве антител к тестируемому маркеру. Иммунофенотипическое двойное окрашивание проводили с использованием моноклональных антител CD44 (FITC) и CD24 (PE). Клетки со средней флуоресценцией CD44-маркера выше 10 3 (по логарифмической шкале) были отнесены к CD44 high субпопуляции. Запись и анализ результатов выполнялись с помощью программы «WinMDi 2.9» (Джозеф Троттер, Ла-Хойя, США).
Разделение CD44 + Фракция клеток ЭК с использованием иммуномагнитной сортировки
Имея в виду, что ОСК с высоким уровнем экспрессии маркера CD44 (CD44 high ), состоящую из гетерогенной популяции CD44 + клеток, обладающих наивысшей онкогенной активностью, они были выделены из общей популяции ЭК с помощью магнитного сортировщика (BDTM Imagnet). Чтобы изолировать CD44 + фракции использовали первичные немеченые моноклональные антитела к маркеру CD44 (BD, 558739) и вторичные мышиные IgG1 Magnetic Particles-DM (BD, 557983) в соответствии с протоколом производителя. Чистота разделения CD44 + клеток от общей популяции ЭК составляло 90%.
Определение онкогенной активности клеток общей популяции и клеток изолированного CD44 + и CD44 - - фракции ЭК
Онкогенная способность всего населения и изолированного CD44 + и CD44 - Фракции ЭК были сравнительно проанализированы описанным выше методом культивирования in vivo. Схема экспериментальной установки представлена на рис. 1.
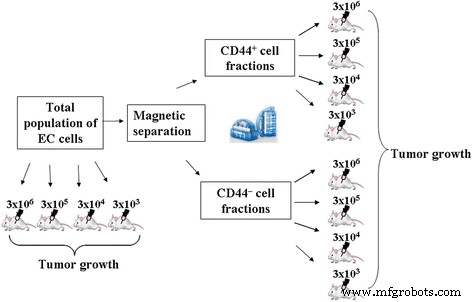
План эксперимента при сравнительном анализе канцерогенной способности всей популяции и изолированного CD44 + и CD44 - EC фракции
В первой серии экспериментов мы оценили онкогенную способность всей популяции и изолировали CD44 + и CD44 - Фракции ЭК при введении их животным в стандартной дозе, используемой для инициирования ЭК (3 × 10 6 клетки в 0,3 мл физиологического раствора).
Животные были разделены на следующие группы ( n =10):
-
Группа 1.1 - введение всей популяции клеток ЭК (3 × 10 6 клеток / животное)
-
Группа 2.1 - введение CD44 + фракция клеток ЭК (3 × 10 6 клеток / животное)
-
Группа 3.1 - введение CD44 - фракция клеток ЭК (3 × 10 6 клеток / животное)
Через 7 дней после инокуляции в каждой из экспериментальных групп подсчитывали общее количество клеток в ПК животных, оценивали фенотипические характеристики клеток (как описано выше) и уровень CD44 высокий / CD117 + соотношение клеток ЕС, определяемое как соотношение CD44 high процент к CD117 + клетки [22]. Пролиферативный потенциал клеток общей популяции ЭК и выделенного CD44 + и CD44 - фракций оценивали на основании следующих данных:коэффициент множественности (MF) избытка клеточной популяции за время культивирования, M =Н / Н 0 ; и удвоение времени (TD), TD =(log 2 2) * t / [журнал 2 ( N / N 0 )], где t время культивирования клеток (ч), N количество ячеек в t время; N 0 - начальный номер ячейки [23].
Во второй серии экспериментов была оценена минимальная доза введенных клеток всей популяции и изолированного CD44 + и CD44 - Фракции ЭК, вызывающие рост опухоли. Суммарная суспензия клеток и выделенный CD44 + и CD44 - Фракции ЭК вводили мышам внутрибрюшинно в дозах 3 × 10 6 , 3 × 10 5 , 3 × 10 4 , и 3 × 10 3 клеток на мышь в 0,3 мл физиологического раствора и культивировали в течение 7 дней на ПК.
Животные, использованные в этой серии экспериментов, были разделены на следующие группы ( n =10):
-
Группа 1.1 - введение всей популяции клеток ЭК (3 × 10 6 cellanimal)
-
Группа 1.2 - введение всей популяции клеток ЭК (3 × 10 5 клеток / животное)
-
Группа 1.3 - введение всей популяции клеток ЭК (3 × 10 4 клеток / животное)
-
Группа 1.4 - введение всей популяции клеток ЭК (3 × 10 3 клеток / животное)
-
Группа 2.1 - введение CD44 + фракция клеток ЭК (3 × 10 6 клеток / животное)
-
Группа 2.2 - введение CD44 + фракция клеток ЭК (3 × 10 5 клеток / животное)
-
Группа 2.3 - введение CD44 + доля клеток ЭК (3 × 10 4 клеток / животное)
-
Группа 2.4 - введение CD44 + фракция клеток ЭК (3 × 10 3 клеток / животное)
-
Группа 3.1 - введение CD44 - фракция клеток ЭК (3 × 10 6 клеток / животное)
-
Группа 3.2 - введение CD44 - фракция клеток ЭК (3 × 10 5 клеток / животное)
-
Группа 3.3 - введение CD44 - фракция клеток ЭК (3 × 10 4 клеток / животное)
-
Группа 3.4 - введение CD44 - фракция клеток ЭК (3 × 10 3 клеток / животное)
В каждой экспериментальной группе общее количество клеток ПК и животных с развитием асцита определяли через 7 дней после инокуляции ЭК.
Синтез нанокомплексов
В Институте сцинтилляционных материалов синтезированы гибридные нанокомплексы, содержащие сферические наночастицы (НЧ) (диаметром 2–3 нм) в концентрации 1,30 г / л и холестерин барана в концентрации 0,55 г / л («Acros Organics», Бельгия). Национальной академии наук Украины (Харьков), как сообщалось [18]. НЧ на основе ортованадатов редкоземельных элементов GdYVO 4 :Eu 3+ сферической формы с концентрацией 1,30 г / л готовили по методике [24]. Водные коллоидные растворы на основе ортованадатов очищены от примесей диализом с использованием мембран «Cellu Sep H1» 3,5 кДа.
В гибридном нанокомплексе отрицательно заряженные НЧ локализуются по периферии частиц холестерина за счет ван-дер-ваальсовых и гидрофобных взаимодействий. НЧ стабилизируют нанокомплексы за счет электростатических взаимодействий. Размеры синтезированных нанокомплексов не превышают 100 нм. Кроме того, НЧ проявляют антиоксидантные свойства и не подвергаются окислению. Этот факт способствует повышению устойчивости водной дисперсии холестерина по отношению к активным формам кислорода. Схематическая структура гибридного нанокомплекса представлена на рис. 2.
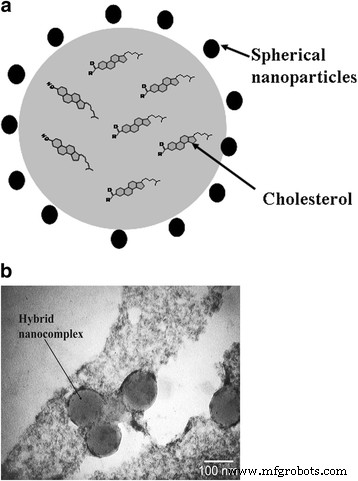
Гибридный нанокомплекс: а схематическое изображение и b Микрофотография с помощью просвечивающей электронной микроскопии гибридных нанокомплексов, полученных из водного раствора холестерина, помещенного на углеродную сетку
Для регистрации накопления гибридных нанокомплексов в клетках во время исследований in vitro в водную дисперсию холестерина можно было дополнительно ввести гидрофобный флуоресцентный краситель 1,1'-диоктадецил-3,3,3 ', 3'-тетраметилиндокарбоцианин перхлорат (DiI). позволяющий в локальной люминесцентной спектроскопии оценивать динамику интеграции комплекса в клеточную мембрану по соотношению полос люминесценции мономера –– «J-агрегат» [25]. Наши более ранние исследования показали, что гибридный нанокомплекс способен интегрироваться не более чем в 10% клеток от общей популяции ЭК и практически во все клетки изолированного CD44 + фракция, имеющая самый высокий канцерогенный потенциал. Это позволяет использовать гибридный нанокомплекс в данной модификации (НЧ + холестерин + DiI) в качестве метода выявления локального накопления нанокомплексов в раковых клетках [26, 27].
Предварительная обработка клеток ЭК наноматериалами
Тотальную суспензию клеток ЭК с гибридными нанокомплексами или НЧ инкубировали в 5% растворе глюкозы (ЗАО «Инфузия», Киев) при комнатной температуре в течение 3 ч. Такое время инкубации ранее считалось оптимальным для связывания нанокомплексов с клетками [26].
Были испытаны следующие варианты предварительной обработки клеток ЭК наноматериалами:
-
Вариант 1–900 мкл клеток ЭК (1 × 10 7 ) Добавляли 100 мкл сферических НЧ (1,3 г / л).
-
Вариант 2–900 мкл клеток ЭК (1 × 10 7 ) Добавляли 100 мкл гибридного комплекса (сферические НЧ (1,3 г / л) + холестерин (0,55 г / л)).
Контролем служили клетки общей популяции ЭК, которые инкубировали в 5% растворе глюкозы без обработки нанокомпозитами. Количество животных в каждой экспериментальной группе было не менее 20.
После инкубации клетки ЭК всех тестируемых групп трижды промывали физиологическим раствором (1:1) центрифугированием (10 мин при 300 g).
Интенсивность развития ЭК после предварительной обработки наноматериалами оценивали внутрибрюшинным введением в дозе 3 × 10 6 клетки в 0,3 мл физиологического раствора. Через 7 дней после инокуляции клеток ЭК во всех исследуемых группах были определены:
-
Общее количество (TN) клеток ЭК в брюшной полости.
-
Скорость ингибирования (Ri) роста ЭК по формуле Ri =(TN (c) - TN (e)):TN (c) × 100%, где TN (c) –– общее количество клеток EAC в ПК контрольная группа, TN (e) - общее количество клеток EAC в ПК опытной группы.
-
Скорость роста (Rg) ЭК рассчитывали по формуле Rg (e) =Rg (c) - Ri, где Rg (e) - скорость роста опухолей экспериментальной группы животных; Rg (c) - скорость роста опухоли контрольной группы, Ri - скорость ингибирования роста ЭК в опытной группе животных; Скорость ингибирования роста ЭК в контроле принималась за 100%, при этом не было ингибирования роста ЭК.
-
CD44 высокий / CD117 + соотношение (соотношение CD44 высокий процент к CD117 + ячеек).
-
Выживаемость животных оценивали к 20 дню после внутрибрюшинной инъекции необработанных и обработанных всеми типами нанокомпозитных ЕС-клеток.
Статистическая обработка проводилась с использованием непараметрического метода Манна-Уитни U Тестирование в программном обеспечении Statistica 6.0. Различия считались статистически значимыми при P <0,05.
Результаты
Полученные результаты свидетельствуют о наличии гетерогенной популяции клеток ЭК, несущих на своей поверхности маркеры CD44, CD24, Sca-1 и маркеры, которые можно отнести к вспомогательно-регуляторным элементам микроокружения (CD117). Концентрации клеток с этими характеристиками в общем пуле ЭК (группа 1.1) показаны в таблице 1 и полностью согласуются с предыдущими данными о субпопуляционном составе ЭК [28]. Идентификация структуры Sca-1 практически во всех клетках ЭК позволяет рассматривать его как универсальный маркер этого типа опухоли.
Наиболее информативной с точки зрения фенотипической идентификации ОСК является экспрессия молекулы CD44, которая сама по себе или в комбинации с другими поверхностными маркерами используется для выделения этой клеточной популяции из различных опухолей, включая ЭК. Согласно классическим представлениям, дифференцировка опухолевых клеток при развитии рака груди сопровождается снижением экспрессии рецептора CD-44 с постепенным его исчезновением и появлением клеток, экспрессирующих маркер CD24 [3].
Кандидатами на роль CSC во время ЭК могут быть клетки с высоким уровнем CD44. фенотип, являющийся частью CD44 + CD24 - - Население. Это предположение о зависимости ЭК от функциональной активности субпопуляции CD44 + клеток тестировали при оценке интенсивности роста опухоли, индуцированного CD44 + и CD44 - фракций и всего населения ЕС. Таблица 1 демонстрирует, что наибольшая опухолиндуцирующая активность была присуща клеткам CD44 + . дробная часть. Собственно, после введения 3х10 6 CD44 + клеток (группа 2.1), абсолютное количество клеток в ПК было в 23 раза выше, чем в общей популяции клеток ЭК (группа 1.1), и в 105 раз больше, чем при CD44 - фракция администрируется (группа 3.1).
При этом обнаружены изменения не только количественного, но и качественного состава развивающейся опухоли. Доля CD44 + сформировал асцит с преобладающим содержанием CD44 + клетки, т.е. CD44 high , CD44 + CD24 - , и CD44 + CD24 + клетки. Кроме того, концентрация CD44 высокая клеток была в 2 раза выше по сравнению с группой 1.1 и в 16 раз выше по сравнению с группой 3.1. Доля CD44 - , напротив, образовалась опухоль, содержащая более зрелые клетки, а именно клетки с CD44 - CD24 + фенотип. Само перераспределение субпопуляционного состава клеток в группе 3.1, по-видимому, определило минимальное абсолютное содержание клеток в ПК.
Важным является установленный факт наличия среди клеток ЭК субпопуляции с CD117 + маркер. Молекула CD117 представляет собой трансмембранный рецептор тирозинкиназы. В нормальных условиях он активируется соответствующим лигандом - фактором роста стволовых клеток (SCGF) [29]. При онкологической патологии происходит лигандон-зависимая активация рецептора c-KIT, что чаще всего (до 92% случаев) является следствием мутации онкогена c-kit или вызвано нарушением механизмов регуляции функции этого рецептора. [30].
Учитывая CD117 + клетки как клетки микроокружения опухоли установленный факт зависимости интенсивности роста опухоли от наличия или отсутствия CD117 + клетки и взаимосвязь между их концентрацией и CD44 high клетки - это логика. Как показано в таблице 1, при инициировании ЭК путем введения всей популяции клеток (группа 1.1.) В ПК было сформировано 34,80 ± 1,27 × 10 7 клетки на уровне CD44 high / CD117 + коэффициент, равный 0,02 отн. единиц.
Онкогенный потенциал CD44 - фракция была в 4 раза ниже (группа 3.1), что проявлялось снижением CD44 высоким / CD117 + в той же степени (в 4 раза) по сравнению с группой 1.1. Это изменение CD44 high / CD117 + индекс был в основном за счет снижения CD44 high концентрация (в 8,5 раза) на фоне пониженного содержания CD117 + ячеек (в 2 раза).
При оценке интенсивности роста асцита, вызванного CD44 + фракции отмечено достоверное увеличение общего количества клеток в ПК (почти в 24 раза по сравнению с группой 1.1). Также немаловажным является двукратное превышение CD44 high концентрация и недостаток CD117 + клетки. В исходном материале CD44 + фракция (до культивирования) согласно данным проточно-цитометрического анализа данных, содержание CD44 высокое клеток было в 15 раз больше, чем в общей популяции клеток ЭК (данные не представлены).
Подводя итог вышесказанному, можно утверждать, что решающую роль в инициации ЭК играют РСК с высокой скоростью экспрессии маркера CD44 (CD44 high ), а одна из важнейших функций CD117 + субпопуляция - это регуляция («сдерживание») онкогенной активности CD44 high клетки. Отсутствие CD117 + клетки (группа 2.1), по-видимому, увеличивают пролиферативный и дифференцировочный потенциал всего пула CD44 + ячеек, что приводит к значительному увеличению общего количества ячеек на ПК.
Анализ пролиферативного потенциала общего пула клеток ЭК и CD44 + фракция поддерживает эту интерпретацию. Показано, что коэффициент размножения (MF) в общей популяции, культивируемой в ПК в группе 2.1. за 7 дней увеличился почти в 24 раза по сравнению с группой 1.1. Это сопровождалось уменьшением удвоения времени клеток с 24,47 ± 2,75 ч в группе 1.1–14,70 ± 1,35 в группе 2.1, что может характеризовать популяцию асцитных клеток, выращенных из CD44 + фракция, как более активно размножающаяся (таблица 1).
Чтобы доказать особую роль CD44 + клеток в инициировании и поддержании опухоли при введении ЭК даже в минимальных дозах, представляло интерес сравнительная оценка канцерогенной способности изолированного CD44 + и CD44 - фракции при приеме внутрь в различных концентрациях. Выяснилось, что после введения 3 × 10 6 клеток от общей популяции ЭК, рост опухоли наблюдался у 100% животных (10/10) (таблица 2). Reducing 10 times the dose of cells administered (3 × 10 5 ) resulted in a proportional decrease in absolute number of cells in the PC, tumor developed only in 50% of animals (Table 2). Reducing the administered dose of total EC population of cells down to 3 × 10 4 did not lead to tumor formation in the PC.
Initiations of EC by introducing of CD44 + cells at concentrations of 3 × 10 6 and 3 × 10 5 cells per animal resulted in almost 100% tumor development for both cases. Herewith, tumorigenic potential of CD44 + fraction exceeded that of total population of EC cells administered in the same doses (in 23 and 21 times, respectively). Moreover, introduction of 3 × 10 4 cells of CD44 + fraction caused a tumor formation in 33% of animals, while total population of EC cells used in the same dose, did not cause the formation of ascites. With the introduction of 3 × 10 3 cells of CD44 + fraction, no animals with the developed EC have been identified.
Fraction of CD44 – cells just in a dose of 3 × 10 6 was capable of forming tumors in 50% of animals, the number of cells in the PC in this case was 4.5 times less than when introducing the total population and in 105.9 times less than when inducing by CD44 + fraction. Thus, the results of this part of research suggest that CSCs are mainly present in the pool of cells with CD44 + phenotype. This emphasizes the importance of this subpopulation of cells in initiation and development of EC.
As noted above, identification and inactivation of CSCs is a major theoretical and practical issue of oncology. On this basis, the next task of our study was to investigate the impact of hybrid nanocomplexes designed at the Institute for Scintillation Materials of National Academy of Sciences of Ukraine on the tumorigenic activity of EC cells.
As Table 3 demonstrates an incubation of EC cells with only NPs as a component of hybrid nanocomplexes (option 1) decreased the concentration of CD44 high virtually twice if compared to the control and 5 times the content CD44 + CD24 – cells in ascites formed in vivo. The number in it of more differentiated CD44 + CD24 + , CD44 – CD24 + cells remained practically unchanged if compared to the control. In this group, there was established reduction of CD117 + cells (35%) at a slightly changed content of Sca-1 + subpopulation. Based on the data, the inhibition rate of EC growth (59.41 ± 3.45%) in variant 1 was accompanied by a twofold decrease in the concentrations of CD44 high cells in comparison with the control that was also reflected in the reduction of CD44 high /CD117 + ratio (Table. 3).
Pretreatment of EC cells with hybrid nanocomplexes (option 2) reduced almost 10 times the concentration of CD44 high and CD44 + CD24 – cells in the developed ascites if compared to the control (Table 3). It should be noted that the concentration of more differentiated CD44 + CD24 + and CD44 – CD24 + cells after this treatment increased slightly if compared to the control. The redistribution pattern of EC subpopulation composition in this option was accompanied with a pronounced enhancement of tumor growth inhibition compared to option 1 (74.70 ± 4.38 and 59.41 ± 3.45%, respectively, P < 0.05) that underlined the importance of cholesterol as a targeted compound of antitumor therapy. Pretreatment with hybrid nanocomplexes (option 2) led to maximal reduction there was found a maximum reduction of CD44 high /CD117 + ratio (10 times) as compared with option 1, that again confirmed a specific role of ratio of these cell subpopulations in the EC growth.
For all the types of EC pretreatment, the reduction of CD44 high /CD117 + ratio was accompanied by a decrease in tumor growth rate and increased survival of animals to day 20 of EC development (Fig. 3).

Tumor growth rate of EC, survival of animals and CD44 high /CD117 + ratio after incubation with nanocomplexes. Note:differences are statistically significant as compared with administration of the control (*), option 1 (**) (P < 0.05)
Discussion
One of the tasks of current oncology is elucidation of the mechanisms of initiation and development of malignant neoplasms. Mandatory participants in these events are the CSCs and so-called accessory-regulatory cells of tumor microenvironment. The variety of functional and structural characteristics of the CSCs in the development of different types of tumors determines the need for their further study. This is facilitated by the expansion of experimental model systems. One of them is the transplantable line of tumor cells of EC.
The elucidation of the peculiarities of this experimental model development, the subpopulation composition of tumor and tumorigenic potential of individual cell populations within the general pool of the EC cells will facilitate the development of new approaches to cancer therapy.
Using the method of phenotypic evaluation of progenitor cells of various levels of differentiation in the tumor focus makes it possible the identifying the stages, dynamics of development and invasiveness of the process. The established fact of heterogeneity of the EC subpopulation composition is important and there has been emphasized the value of CD44 + subpopulation in maintaining the growth of this type of tumor.
The most important role in implementing a tumorigenesis is played by an expression rate of the molecule. Indeed, in contrast to leukocytes for adhesion of those normally a low expression rate of CD44 receptor is required, triggering and self-maintenance in CSCs are implemented its much greater density on a cell surface [31].
It is known that CD44-glycoprotein is a hyaluronic acid (HA) receptor, a main component of extracellular matrix. The emerging set of HA-CD44 activates many receptor tyrosine kinases, resulting in activation of PI3K/Akt/ mTOR way [32, 33], which plays the role of a single universal signal transmission mechanism to the translation apparatus and is responsible for the integration of proliferative stimuli.
Among two known CD44-isoforms in normal hematopoietic cells its standard isoform (CD44s) is predominantly expressed [34]. In most malignant tissues there were detected both CD44s and variable isoforms of CD44- molecule (CD44v), resulting from alternative splicing of exons 6-15. Namely alternative splicing leads to a lengthening of CD44-extracellular domain, promoting its greater interaction with HA and tumor metastasis [35]. Due to that the role of CD44 high cells in triggering and maintaining the tumorogenesis is clear. It was previously found that a minor subpopulation of CD44 high cells had a high proliferative potential and played a critical role in EC developing [20].
In this paper, a special role of CD44 + -cells of the EC in initiation and maintenance of the tumor process in the EC under administration even in minimal doses has been shown. CD44 + cells were able to form a tumor even at a cell concentration of 100 times lower (10 4 cells/ mouse) if compared with the introduction of a total EC population (10 6 cells / mouse). The belonging of tumor cells to the CD44 + fraction was also confirmed by the fact that the EC initiation by the fraction of CD44-cells even at a dose of 10 6 cells / mouse caused the formation of a tumor only in 50% of cases, with an absolute number of cells in the PC 5 times less than in under introduction of a similar amount of the total population of EC and more than 100 times less than after the introduced CD44 + -fraction.
This is in accordance with the data of Shipitsin M et al. has shown that CD44 + and CD24 + cells in breast cancer development there are cell populations with different genetic profiles [36]. The research performed by Shipitsin M CD24 + cells have been noted to be more differentiated, while more progenitor-like functions are inherent to CD44 + клетки. The research performed by Shipitsin M CD24 + cells have been noted to be more differentiated, while more progenitor-like functions are inherent to CD44 + клетки. The authors suggest that CD24 + cells can be derived from CD44 + cells [36]. Fillmore C. and Kuperwasser C. supposed that CD24 + population was mainly characterized by less differentiated basal type of breast cancer, and CD44 + cells caused the development of luminal form of breast cancer, being more differentiated type of tumor [37].
Analyzing the patterns of tumor development, the classic hypothesis of «seed and soil» looks very actual [38], which postulated that an appropriate microenvironment (soil) is required for optimal growth of tumor cells (CSCs).
Most often the carcinoma-associated fibroblasts (CAFs) act as a tumor stroma in breast cancer and pancreatic cancer [39]. It has been shown that the CAFs, derived from invasive forms of human breast carcinomas, activated much stronger the growth of human breast cancer cell line MCF-7-Ras when administered to immunodeficient mice if compared with normal fibroblasts [9]. This function is implemented by the microenvironment cells due to the secretion by them of cytokines, chemokines and growth factors [10, 40].
Although so far the phenotypic identification of the microenvironment cells for various types of tumor has remained a subject of debate, most often used for this purpose the surface markers of primitive hematopoietic and endothelial cells, including c-kit (CD 117), CD133, VE-cadherin, VEGFR-2 and endoglin are used [41]. In this experimental model the most probable candidate to the role of tumor microenvironment cells is CD 117 + .
It is known that the c-KIT receptor (CD117 + ) is highly expressed in normal epithelium of the breast and progressively decreases with the development of breast carcinoma in situ and is almost completely lost in invasive breast cancer [42, 43]. Some authors proposed this kind of change in the expression rate of this marker as a possible test to assess the effectiveness of antitumor therapy [44].
Previously, after analysis of the significance of the content ratios for different subpopulations of EC cells when maintaining tumor growth, we proposed to use the CD44 high /CD117 + ratio as a prognostic criterion of tumor development [22].
Adequacy of using this index is confirmed in this study using the applied nanocomposites as therapeutic agents when treating the EC. The inhibition rate of EC growth (59.41 ± 3.45%) when treated with spherical NPs (option 1) was accompanied by a 2-fold decrease if compared to the control in the CD44 high -cell concentration, which was reflected in the reduced CD44 high / CD117 + показатель. The maximum decrease in the CD44 high /CD117 + index (10 times if compared to option 1) was established using the hybrid nanocomplexes for a pre-treatment of EC cells. Thus, many cells of a total pool of EC, but primarily those with the phenotype CD44 high and CD117 + , can be the target of the effect of the studied nanocomplexes (both direct and indirect). A significant decrease in their concentrations in the growing pool of EC after pretreatment with hybrid nanocomplexes clearly coincides with a reduced intensity of tumor growth.
Judging by the decrease in the amount of CD44 high as the most potent CSCs forming the entire subsequent series of advanced tumor cells, the main component in manifestation of antitumor effect of the synthesized hybrid nanocomplexes is spherical NPs. Introduction of cholesterol having affinity to tumor cell membranes into composition of hybrid nanocomplexes enhanced an inhibitory activity of NPs. Similar data were obtained by Betker J.L. et al. after analysis of the structure and functioning principles of the membranes of tumor cells. The authors concluded that the incorporation of cholesterol into membranes of tumor cells could be a prerequisite for a targeted delivery of liposomes with therapeutic agents directly into a cell.
Thus, the importance of cooperative interactions of cells with different phenotypic signs in maintaining the EC growth has been proven. The cells with the CD44 high phenotype being the part of the population of CD44 + CD24 – can be considered as CSCs in this model system. The use of new forms of nanocomposites that are capable to bind to CSCs and induce tumor destruction as the EC is a promising direction the treatment of oncopathology.
Conclusions
- 1.
On the base of the findings of phenotypic assessment and functional potential studies, the Ehrlich carcinoma is a heterogeneous population of tumor cells of varying differentiation extent referred to high and less potent tumor-inducing precursors, as well as the cells composing their microenvironment.
- 2.
A high (tenfold) tumorigenic activity of the EC CD44 + cells if compared to CD44 – cells was proven. In this pair of comparison, the CD44 + cells had a higher potential of generating in PC of CD44 high , CD44 + CD24 – , CD44 + CD24 + cell subpopulations, highlighting the presence of CSCs in a pool of CD44 + cells.
- 3.
There was found an ability of the synthesized nanocomplexes based on rare earth orthovanadates and cholesterol to inhibit the growth of CD44 + cell pool (CD44 high , CD44 + CD24 – , CD44 + CD24 + ) that was accompanied by a reduced intensity of EC growth (by 75%) and increased survival of the animal with tumors (in 3.5 times) in comparison with the control.
- 4.
It has been shown that the reduction in tumor growth rate after pretreatment with hybrid nanocomplexes was accompanied with a change in the composition of EC subpopulation that was reflected in a decrease in the CD44 high /CD117 + соотношение. This ratio can be offered as one of diagnostic and prognostic tests of the severity and extent of oncology inactivation.
Сокращения
- BC:
-
Breast cancer
- CAFs:
-
Carcinoma-associated fibroblasts
- CSCs:
-
Cancer stem cells
- DiI:
-
1,1′-Dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate
- EC:
-
Ehrlich carcinoma
- HA:
-
Hyaluronic acid
- MF:
-
Multiplicity factor
- NOD/SCID mice:
-
Nonobese diabetic-severe combined immunodeficiency mice
- NPs:
-
Nanoparticles
- PC:
-
Peritoneal cavity
- Rg:
-
Growth rate of EC
- Ri:
-
Inhibition rate of EC growth
- SCID mice:
-
Severe combined immunodeficiency mice
- TD:
-
Time doubling
Наноматериалы
- Батареи специального назначения
- Искусственная кровь
- Солнечный элемент
- Нанодеревья для сенсибилизированных красителем солнечных элементов
- Высокоэффективные графеновые солнечные элементы
- Нано-гетеропереходы для солнечных элементов
- Демонстрация гибкого биосенсора на основе графена для чувствительного и быстрого обнаружения клеток рака яи…
- Наночастицы для терапии рака:текущий прогресс и проблемы
- Доставка лекарств на основе клеток для онкологических приложений
- Что такое стволовая стена?