


Ингибированный MicroRNA-301 сдерживает ангиогенез и рост клеток в плоскоклеточной карциноме пищевода путем повышения PTEN
Аннотация
Цель
Плоскоклеточный рак пищевода (ESCC) характеризуется ранним метастазированием и поздней диагностикой. Известно, что микроРНК-301 (miR-301) участвует в различных формах рака. Тем не менее, эффекты miR-301 на ESCC остаются неизученными. Таким образом, мы стремимся изучить роль miR-301 в прогрессировании ESCC.
Методы
Оценивали экспрессию miR-301 и гомолога фосфатазы и тензина (PTEN) в тканях и клеточных линиях ESCC. Затем отобранные клетки обрабатывали измененным олигонуклеотидом и плазмидой miR-301 или PTEN, а затем оценивали способность к образованию колоний, жизнеспособность клеток, миграцию, инвазию, распределение клеточного цикла и апоптоз клеток ESCC. Кроме того, также оценивались рост опухоли и плотность микрососудов (MVD), и была подтверждена целенаправленная связь между miR-301 и PTEN.
Результаты
MiR-301 был активирован, а PTEN подавлен в тканях и клетках ESCC. Клетки KYSE30 и клетки Eca109 были отобраны для функциональных анализов. В клетках KYSE30 ингибирование miR-301 или сверхэкспрессия PTEN подавляло злокачественное поведение клеток, а подавление PTEN устраняло влияние ингибирования miR-301 на прогрессирование ESCC. В клетках Eca109 сверхэкспрессия miR-301 или ингибирование PTEN способствовали злокачественному поведению клеток, а сверхэкспрессия PTEN обращала эффекты повышения miR-301 на прогрессирование ESCC. Анализ in vivo показал, что ингибирование miR-301 или сверхэкспрессия PTEN подавляли рост опухоли ESCC и MVD, а повышение miR-301 или снижение PTEN имели противоположные эффекты. Более того, мишенью для PTEN был miR-301.
Заключение
Взятые вместе, результаты нашего исследования показали, что miR-301 влияет на рост клеток, метастазирование и ангиогенез посредством регуляции экспрессии PTEN в ESCC.
Введение
Рак пищевода (ЭК), 8-е место по распространенности в мире, является критическим злокачественным новообразованием с высокой смертностью и плохим прогнозом [1]. Плоскоклеточная карцинома пищевода (ESCC), на которую приходится около 90% всех случаев ЭК, является основной формой ЭК в Китае [2]. Множественные причины, включая низкий социально-экономический статус, курение, употребление алкоголя, плохое питание, пищу, богатую нитрозаминами или микотоксинами, приводят к возникновению ESCC [3]. Несмотря на благоприятные клинические исходы, а также на назначение препарата, у пациентов с ESCC прогноз по-прежнему плохой, 5-летняя выживаемость составляет 15-25% [4]. Следовательно, крайне важно подтвердить онкогены или гены, репрессивные к опухоли, которые могут функционировать как биомаркеры в развитии ESCC, чтобы обеспечить более эффективные терапевтические методы для пациентов с ESCC.
МикроРНК (миРНК) представляют собой небольшие некодирующие РНК, играющие важную роль в модуляции экспрессии генов [5], и выяснено, что они обладают способностью влиять на прогрессирование опухоли посредством регулирования стабильности мРНК и способности мРНК [6]. Было обнаружено, что количество miRNA, таких как miR-4324 [7], miR-889-3p [8] и miR-9 [9], связано с процессом ESCC. MiR-301 является членом miRNA, которая образована транскрипционной единицей fam33a, расположенной в 17q22-23 в геноме человека. Сверхэкспрессия miR-301 была ранее идентифицирована, что свидетельствует о ее причастности к заболеваниям человека [6, 10]. Однако функциональные механизмы miR-301 не были обнаружены в ESCC. Более того, подтверждено, что фосфатаза и гомолог тензина (PTEN) часто разрушаются в опухолях и становятся мишенью мутаций зародышевой линии у онкологических больных, что играет ингибирующую роль в опухолях [11]. Было подтверждено, что нарушение регуляции PTEN коррелирует с развитием ESCC [12]. Интересно, что недавнее исследование показало, что miR-301 нацелен на PTEN при немелкоклеточном раке легкого [13]. Однако эти отношения нацеливания между miR-301 и PTEN в развитии ESCC еще предстоит раскрыть. Наше исследование было сосредоточено на эффектах miR-301 и PTEN на прогрессирование ESCC, которые остаются в значительной степени неизвестными и являются новинками. Мы сделали вывод, что miR-301 может влиять на ангиогенез и рост клеток в ESCC, модулируя экспрессию PTEN.
Материалы и методы
Этика
Письменное информированное согласие было получено от всех пациентов до начала исследования. Протоколы этого исследования были одобрены этическим комитетом Второй больницы Цзилиньского университета и основаны на этических принципах Хельсинкской декларации для медицинских исследований с участием людей в качестве субъектов. Эксперименты на животных строго соответствовали Руководству по содержанию и использованию лабораторных животных, выпущенному Национальными институтами здравоохранения. Протокол экспериментов на животных был одобрен Комитетом по уходу и использованию животных Второй больницы Цзилиньского университета.
Темы исследования
Сто десять образцов тканей ESCC и прилегающих нормальных тканей (> 5 см от опухоли) были собраны у пациентов с ESCC (78 мужчин и 32 женщины), перенесших эзофагэктомию в отделении торакальной хирургии Второй больницы Университета Цзилинь с января. 2015 г. - декабрь 2017 г. Среди 110 пациентов было 84 случая> 60 лет и 26 случаев ≤ 60 лет; размер опухоли:65 случаев ≥ 5 см и 45 случаев <5 см; 71 случай без метастазов в лимфатические узлы (LNM) и 39 случаев с LNM; стадия опухоли, узла и метастазов (TNM):60 случаев - I + II стадия, 50 - III стадия; локализация опухоли:13 случаев были верхними ESCC, а 97 случаев - средними и нижними ESCC. Всем пациентам был поставлен диагноз ESCC, и они ранее не принимали лучевую терапию или химиотерапию. Опухоли полностью иссечены, отрицательный хирургический край подтвержден патологией. Согласно критериям стадирования ESCC, предложенным Международным союзом по борьбе с раком (UICC) в 2009 году [14], послеоперационная патологическая стадия пациентов была идентифицирована как стадия pT1-4N1-2 (I-IIIb). После операции у пациентов не было серьезных осложнений, и периоперационная смерть была исключена.
Количественная полимеразная цепная реакция с обратной транскрипцией (RT-qPCR)
Суммарные РНК в тканях и клетках экстрагировали с использованием наборов Trizol (Invitrogen Inc., Карлсбад, Калифорния, США). Измеряли концентрацию и качество РНК. Праймеры РНК (таблица 1) были разработаны и синтезированы TaKaRa Biotechnology Co., Ltd. (Ляонин, Китай). РНК была обратно транскрибирована в кДНК в соответствии с инструкциями набора реагентов Takara PrimeScript ™ RT с g DNA Eraser (Takara). Мы выполнили qPCR на Light Cycler 480II (Roche) с мастер-миксом Power PCR SYBR green PCR (Takara). U6 использовали в качестве контроля загрузки miR-301, а β-актин использовали в качестве внутреннего стандарта для PTEN. Данные были проанализированы с использованием 2 - △△ Ct метод [15].
Вестерн-блот-анализ
Буфер для лизиса RIPA (Институт биотехнологии Beyotime, Шанхай, Китай) использовали для экстракции общего белка в клетках и тканях, и белок количественно определяли с помощью набора для анализа белков BCA (Beyotime). Измеряли концентрацию белка в каждом образце и проводили электрофорез в 10% -ном додецилсульфате натрия в полиакриламидном геле. Образцы переносили на нитроцеллюлозные мембраны, которые затем блокировали 5% сухим обезжиренным молоком при 4 ° C в течение ночи. После этого к мембранам добавляли первичные антитела PTEN и β-актин (оба 1:500 и от Santa Cruz Biotechnology Inc, Калифорния, США) для инкубации в течение ночи, затем добавляли соответствующие вторичные антитела и инкубировали в течение 1 часа. После погружения в усиленный хемилюминесцентный реагент (Pierce Chemical Inc., Даллас, Техас, США) на 1 мин мембраны экспонировали в темноте и проявляли с использованием мини-хемилюминесцентного устройства для визуализации LAS4000. Значения серого оценивали с помощью программного обеспечения системы визуализации с β-актином в качестве контроля; следовательно, был получен окончательный относительный экспрессированный белок. Полосы белка анализировали с помощью программного обеспечения ImageJ2x.
Анализ двойного люциферазного репортерного гена
Было предсказано, что последовательность 3'-нетранслируемой области (UTR) PTEN будет взаимодействовать с miR-301, или была синтезирована мутированная последовательность в пределах предсказанных сайтов-мишеней и вставлена в сайты XbaI и FseI репортерного вектора контрольной люциферазы pGL3 (Promega, Висконсин, США). Затем были получены векторы pGL3-PTEN-wt и pGL3-PTEN-mut. Правильно идентифицированные плазмиды-репортеры люциферазы wt и mut с имитатором miR-301 и имитатором NC были котрансфицированы в клетки KYSE30 и Eca109 в течение 48 часов. Впоследствии клетки лизировали, и активность люциферазы определяли, соответственно, с помощью наборов для обнаружения люциферазы (Promega).
Культура клеток, группировка и трансфекция
Клеточные линии ESCC (KYSE-150, KYSE-30, Eca109 и KYSE-70) были получены из Шанхайского института биохимии и клеточной биологии Китайской академии наук (Шанхай, Китай), а эпителиальные клетки пищевода человека (HEEC) были приобретены. от Mingzhou Biotechnology Co., Ltd. (Чжэцзян, Китай). Клетки культивировали в среде RPMI 1640 (Invitrogen) с добавлением 10% фетальной бычьей сыворотки (FBS, Life Technologies, США), 100 единиц / мл пенициллина G натрия (Sigma) и 100 мкг / мл сульфата стрептомицина (Sigma). Экспрессия MiR-301 и экспрессия мРНК PTEN в каждой клеточной линии были измерены с помощью RT-qPCR, и для последующих клеточных экспериментов была выбрана клеточная линия с самой высокой и самой низкой относительной экспрессией.
Клетки KYSE-30 были разделены на 7 групп и, соответственно, обработаны ингибитором miR-301, отрицательным контролем ингибитора (NC), pcDNA-PTEN (названный сверхэкспрессированным (oe) -PTEN), pcDNA-NC (названный oe-NC), Ингибитор miR-301 + малая интерферирующая РНК (si) -PTEN или ингибитор miR-301 + si-NC. Клетки Eca109 также были разделены на 7 групп и по отдельности обработаны миметиком miR-301, миметиком NC, si-PTEN, si-NC, миметиком miR-301 + oe-PTEN, миметиком miR-301 + oe-NC. ингибитор NC, ингибитор miR-301, миметик miR-301, миметик NC, si-NC и si-PTEN были приобретены у GenePharma Ltd., Company (Шанхай, Китай); pcDNA-PTEN NC и pcDNA-PTEN были получены от (Shanghai Sangon Bio-Technology Corporation (Шанхай, Китай)). Клетки были временно трансфицированы в клетки ESCC липофектамином 2000 (Invitrogen), когда слияние клеток достигло 60%.
Набор для подсчета клеток (CCK-8)
Клетки высевали в 96-луночный планшет (1 × 10 3 клеток / лунку) и инкубировали в течение разных периодов времени. После инкубирования в течение 24 ч, 48 ч, 72 ч и 96 ч в каждую лунку добавляли 10 мкл раствора CCK-8 (5 мг / мл), а затем клетки каждой группы инкубировали при 37 ° C без света. выдержка 2 ч. Значения оптической плотности (OD) при 450 нм анализировали с помощью ридера для микропланшетов (Bio-Rad Laboratories, Hercules, CA, USA).
Анализ образования колоний
Клетки высевали по 500 клеток / лунку в 6-луночные планшеты после трансфекции и культивировали в течение 14 дней. Колонии фиксировали метанолом, окрашивали 0,5% кристаллическим фиолетовым и подсчитывали под инвертированным микроскопом.
Анализ Transwell
Ячейки (5 × 10 3 ), инкубированные в среде RPMI 1640, высевали в апикальные камеры устройств Transwell с непокрытой или покрытой матригелем мембраной (Corning, NY, США). Через 24 часа клетки апикальных камер удаляли, а клетки, оставшиеся на нижней стороне, фиксировали и затем окрашивали 0,1% кристаллическим фиолетовым. Микроскоп (Olympus Corporation, Токио, Япония) использовался для подсчета в 3 случайных полях для расчета количества клеток.
Проточная цитометрия
Клеточный цикл и апоптоз оценивали с помощью проточной цитометрии. Аннексин V-флуоресцеина изотиоцианат (10 мкл) и иодид пропидия (PI; 5 мкл, Sigma) инкубировали с клетками (5 × 10 5 клеток / лунку) в темноте при 4 ° C в течение 30 мин. Процент апоптозных клеток рассчитывали с использованием проточного цитометра (BD Biosciences, Калифорния, США) с программным обеспечением FlowJo версии 10 (FlowJo LLC, OR, США).
Для оценки клеточного цикла клетки (5 × 10 5 клеток / лунку) фиксировали 75% этанолом в течение ночи при 4 ° C и окрашивали 5 мкл PI / рибонуклеазой A (Sigma) при 4 ° C в течение 30 мин в темноте. Данные анализировали с помощью проточного цитометра (BD Biosciences). Сигналы флуоресценции (14 000) каждого образца были собраны и рассчитаны с использованием программного обеспечения ModFit LT версии 3.2 (Verity Software House, Inc., ME, США).
Подкожный опухолевидный генез у голых мышей
Сорок две самки голых мышей BALB / c-nu (возраст 4 нед, вес 16-24 г) были получены из Центра экспериментальных животных Цзилиньского университета (Чанчунь, Китай). Голых мышей разделили на 14 групп ( n =3). Голым мышам семи групп, соответственно, вводили клетки KYSE-30 в соответствии с группировкой клеток, а голым мышам в остальных семи группах по отдельности вводили клетки Eca109 на основе группировки. Концентрация трансфицированных клеток KYSE-30 и Eca109 была доведена до 5 × 10 6 клеток / 100 мкл. Голых мышей фиксировали и подкожно вводили соответствующие клетки ESCC в стерильных условиях. Наибольшую длину (L) и ширину (W) опухолей измеряли каждую неделю, а объем опухоли ( V ) =1/2 × L × Вт 2 . Голых мышей умерщвляли в 5-м -м неделю инъекции с резекцией опухолей, и опухоли взвешивали и фотографировали. Скорость образования опухоли рассчитывали как количество мышей с подкожной опухолью / общее количество инъецированных голых мышей в группе × 100%. Время инъекции принимали за абсциссу, а размер опухоли - за ординату; таким образом, была построена кривая роста опухоли.
Иммуногистохимическое окрашивание
Опухолевые ткани мышей nude фиксировали 10% формальдегидом, заливали парафином и делали срезы на 4 мкм. Затем срезы сушили при 60 ° C в течение 2 часов, депарафинизировали ксилолом, дегидратировали градиентным этанолом и инкубировали с 50 мкл 3% H 2 О 2 на 10 мин. После этого срезы замачивали в 0,01 М буферном растворе лимонной кислоты, кипятили при 95 ° C в течение 20 минут, блокировали нормальным рабочим раствором козьей сыворотки при 37 ° C в течение 10 минут и добавляли CD 34 (1:100, Санта-Крус) при 4 ° C в течение ночи. После этого срезы были дополнены меченным HRP козьим анти-кроличьим / мышиным IgG-полимером (ZSGB-Bio, Пекин, Китай), окрашены гематоксилином, дегидратированы и проницаемы, а затем запечатаны нейтральным бальзамом. PBS использовали для замены первичных антител в качестве NC. Измерение плотности микрососудов (MVD):срезы наблюдали под микроскопом с малым увеличением. Эндотелиальная клетка или кластер эндотелиальных клеток окрашивались в коричневато-желтый цвет и значительно отличались от окружающих опухолевых клеток, а соединительные ткани были взяты за микрососуд. Структура ветви также принималась за сосуд, если она была разъединена, а сосуды с размером просвета> 8 эритроцитов, мышечным слоем или более толстым просветом исключались. Было записано количество микрососудов 3 высоких полей зрения, и среднее количество было MVD для каждого случая.
Статистический анализ
Все статистические анализы были выполнены с использованием программного обеспечения SPSS version SPSS 21.0 (IBM Corp. Armonk, NY, USA) и представлены с помощью Graphpad Prism Software 6.0. Данные были выражены как среднее ± стандартное отклонение. Различия между двумя независимыми группами проверяли с помощью t-критерия Стьюдента. Односторонний дисперсионный анализ был выполнен для сравнения трех или более групп. P значение <0,05 свидетельствует о статистически значимой разнице.
Результаты
MiR-301 сильно экспрессируется, тогда как PTEN плохо экспрессируется в тканях и клетках ESCC
Экспрессия MiR-301 и PTEN в тканях ESCC и прилегающих нормальных тканях оценивалась с помощью RT-qPCR и вестерн-блот-анализа, чтобы выявить их роль в ESCC, и было обнаружено, что (рис. 1a-c) miR-301 была усилена, в то время как PTEN подавлялась в тканях ESCC. Пациенты были разделены на группы с низкой и высокой экспрессией в соответствии со средним значением экспрессии miR-301 или PTEN для анализа корреляции между экспрессией miR-301 или PTEN и клинико-патологическими характеристиками пациентов с ESCC. Результаты показали, что экспрессия miR-301 / PTEN не была связана с возрастом, полом, размером опухоли, локализацией и дифференцировкой, а коррелировала со стадией TNM и LNM пациентов с ESCC (таблица 2).
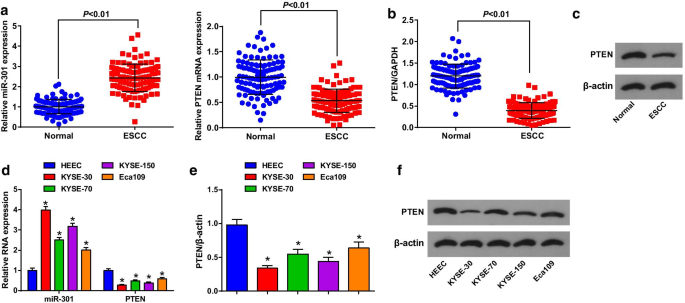
MiR-301 высоко экспрессируется, тогда как PTEN плохо экспрессируется в тканях и клетках ESCC. а Экспрессия miR-301 и экспрессия мРНК PTEN в ткани ESCC, обнаруженная с помощью RT-qPCR; б экспрессия белка PTEN в ткани ESCC, обнаруженная с помощью вестерн-блоттинга; c полосы белка PTEN в ткани ESCC при Вестерн-блоттинге; г экспрессия miR-301 и экспрессия мРНК PTEN в клеточной линии ESCC, обнаруженная с помощью RT-qPCR; е экспрессия белка PTEN в клеточной линии ESCC, обнаруженная с помощью вестерн-блоттинга; е белковые полосы PTEN в Вестерн-блоттинге. * P <0,05 по сравнению с HEEC. Данные были выражены как среднее значение ± стандартное отклонение, а t-тест был проведен для сравнения между двумя группами
Затем определяли экспрессию miR-301 и PTEN в 4 клеточных линиях ESCC и HEEC с помощью RT-qPCR и вестерн-блоттинга. Мы обнаружили, что (рис. 1d-f) miR-301 активируется, а PTEN подавляется в линиях клеток ESCC, среди которых KYSE-30 имеет самую высокую экспрессию miR-301 и самую низкую экспрессию PTEN, тогда как Eca109 имеет противоположную тенденцию. Таким образом, в клеточных экспериментах клеточная линия KYSE-30 обрабатывалась подавленным miR-301 / сверхэкспрессированным PTEN, а клеточная линия Eca109 обрабатывалась сверхэкспрессированным miR-301 / подавленным PTEN.
На PTEN нацелен miR-301
Биоинформатическое программное обеспечение (http://www.microrna.org/) предсказало, что PTEN был геном-мишенью для miR-301 (рис. 2a). Кроме того, с помощью двойного анализа репортерного гена люциферазы было подтверждено, что активность люциферазы была значительно снижена в клетках ESCC, котрансфицированных вектором PTEN-wt и миметиком miR-301, по сравнению с клетками, котрансфицированными вектором PTEN-mut и миметиком miR-301, это означает, что miR-301 может особенно связываться с PTEN (рис. 2b, c).
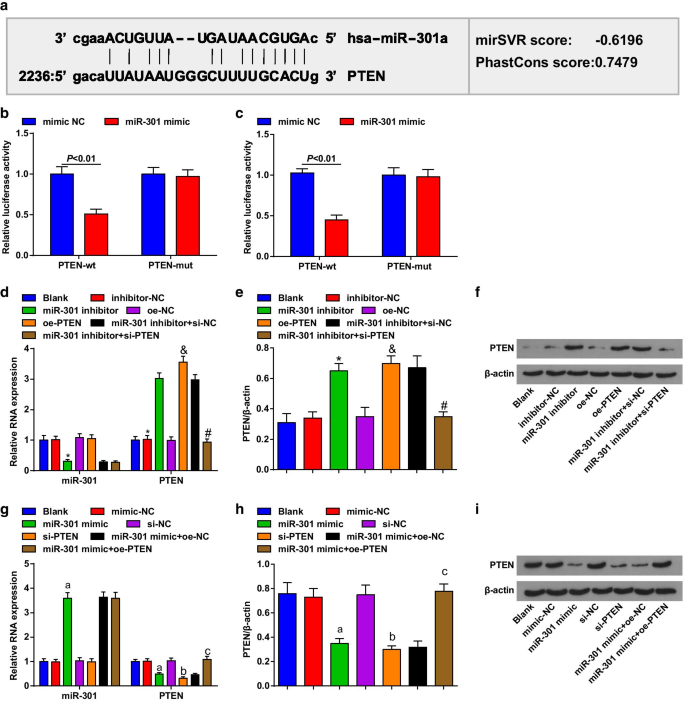
PTEN является целевым геном miR-301. а Сайты связывания miR-301 и PTEN были предсказаны с помощью программного обеспечения для онлайн-прогнозирования; б целевую связь между miR-301 и PTEN в клетках KYSE-30 оценивали с помощью анализа двойного люциферазного репортерного гена; c целевую связь между miR-301 и PTEN в клетках Eca109 оценивали с помощью анализа двойного люциферазного репортерного гена; г экспрессия miR-301 и экспрессия мРНК PTEN в клетках KYSE-30, обнаруженная с помощью RT-qPCR после подавления miR-310 или усиления PTEN; е экспрессия белка PTEN в клетках KYSE-30, обнаруженная с помощью вестерн-блоттинга после подавления miR-310 или повышения уровня PTEN; е белковые полосы PTEN в клетках KYSE-30 при Вестерн-блоттинге после подавления miR-310 или усиления PTEN; г экспрессия miR-301 и экспрессия мРНК PTEN в клетках Eca109, обнаруженная с помощью RT-qPCR после активации miR-310 или подавления PTEN; ч экспрессия белка PTEN в клетках Eca109, обнаруженная с помощью вестерн-блоттинга после активации miR-310 или подавления PTEN; я полосы белка PTEN в клетках Eca109 при Вестерн-блоттинге после активации miR-310 или подавления PTEN. * P <0,05 по сравнению с группой ингибитор-NC, & P <0,05 по сравнению с группой oe-NC, # P <0,05 по сравнению с группой ингибитор miR-301 + si-NC, a P <0,05 по сравнению с группой мимик-NC, b P <0,05 по сравнению с группой si-NC, c P <0,05 по сравнению с группой миметиков miR-301 + oe-NC; N =3. Данные были выражены как среднее значение ± стандартное отклонение, и t-тест был проведен для сравнения между двумя группами. ANOVA использовался для сравнения между несколькими группами
Для оценки экспрессии miR-301 и PTEN в трансфицированных клетках использовали RT-qPCR и вестерн-блоттинг, и было обнаружено, что в клетках KYSE-30 (рис. 2d-f) клетки, обработанные ингибитором miR-301, подавляли miR-301. , при усиленной регуляции PTEN; клетки, обработанные pcDNA-PTEN (oe-PTEN), повышали экспрессию PTEN, а si-PTEN обращал действие ингибитора miR-301 на экспрессию PTEN. В клетках Eca109 (Fig. 2g – i) клетки, обработанные miR-301, имитируют повышенную регуляцию miR-301, тогда как понижающую регуляцию PTEN; клетки, обработанные si-PTEN, снижали экспрессию PTEN, а pcDNA-PTEN (oe-PTEN) обращал ингибирующую роль миметика miR-301 в экспрессии PTEN. Эти данные свидетельствуют о том, что miR-301 нацелена на PTEN.
Ингибирование miR-301 или сверхэкспрессия PTEN ограничивает жизнеспособность клеток ESCC; Повышенный уровень miR-301 или снижение PTEN способствует жизнеспособности клеток ESCC
Жизнеспособность клеток ESCC оценивали с использованием анализов образования колоний и CCK-8. Результаты показали, что в линии клеток KYSE-30 (рис. 3a – c) трансфекция ингибитором miR-301 или oe-PTEN подавляла способность к образованию колоний и жизнеспособность клеток; трансфекция заглушенного PTEN устраняет влияние ингибитора miR-301 на жизнеспособность клеток ESCC; в клеточной линии Eca109 (рис. 3d-f) трансфекция миметика miR-301 или si-PTEN способствовала образованию колоний и жизнеспособности клеток; Сверхэкспрессия PTEN изменила промоторную роль повышения miR-301 в способности образовывать колонии и жизнеспособности клеток Eca109. Эти результаты свидетельствуют о том, что нокдаун miR-301 или сверхэкспрессия PTEN подавляют жизнеспособность клеток ESCC, чему способствует повышение miR-301 или ингибирование PTEN.
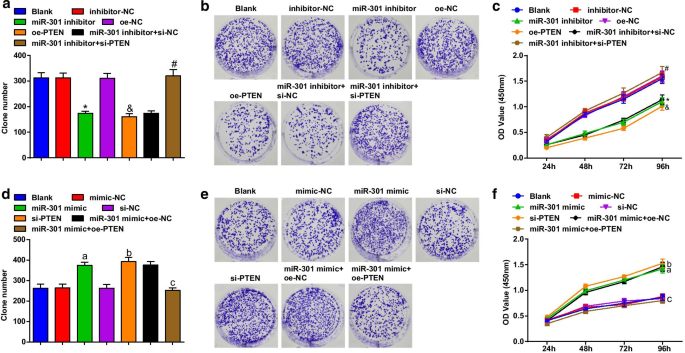
Ингибирование miR-301 или сверхэкспрессия PTEN ограничивает жизнеспособность клеток ESCC; повышенный miR-301 или пониженный PTEN способствует жизнеспособности клеток ESCC. а Количество колоний в клетках KYSE-30 после трансфекции, обнаруженное с помощью анализа образования колоний после подавления miR-310 или усиления PTEN; б способность клеток KYSE-30 образовывать колонии после трансфекции, определяемую с помощью анализа образования колоний после подавления miR-310 или усиления PTEN; c жизнеспособность клеток KYSE-30 после трансфекции, определенная с помощью анализа CCK-8, после подавления miR-310 или усиления PTEN; г количество колоний в клетках Eca109 после трансфекции, обнаруженное с помощью анализа образования колоний после активации miR-310 или подавления PTEN; е способность клеток Eca109 образовывать колонии после трансфекции, определяемую с помощью анализа образования колоний после активации miR-310 или подавления PTEN; е жизнеспособность клеток Eca109 после трансфекции, обнаруженная с помощью анализа CCK-8, после активации miR-310 или подавления PTEN; * P <0,05 по сравнению с группой ингибитор-NC; & P <0,05 по сравнению с группой oe-NC; # P <0,05 по сравнению с группой ингибитор miR-301 + si-NC; а P <0,05 по сравнению с группой мимик-NC; б P <0,05 по сравнению с группой si-NC; c P <0,05 по сравнению с группой миметиков miR-301 + oe-NC, N =3. Данные были выражены как среднее ± стандартное отклонение, и ANOVA использовался для сравнения между несколькими группами
Ингибированный miR-301 или сверхэкспрессируемый PTEN подавляет миграцию и вторжение клеток ESCC; Повышенный уровень miR-301 или снижение PTEN вызывает миграцию и инвазию клеток ESCC
Способность клеток ESCC к миграции и инвазии оценивали с помощью анализа Transwell. Результаты свидетельствуют о том, что в клеточной линии KYSE-30 (рис. 4a, b) способность к миграции и инвазии клеток ограничивается ингибированием miR-301 или сверхэкспрессией PTEN; подавляющая роль ингибитора miR-301 в способностях миграции и инвазии клеток была обращена si-PTEN. В клеточной линии Eca109 (рис. 4c, d) способность клеток к миграции и инвазии усиливалась после трансфекции миметика miR-301 или si-PTEN; сверхэкспрессия PTEN обращала влияние миметика miR-301 на клеточную миграцию и способность к инвазии. Приведенные выше данные предполагают, что миграция и инвазия клеток ESCC ингибируются репрессией miR-301 или повышением PTEN, тогда как им способствует повышающая регуляция miR-301 или понижающая регуляция PTEN.
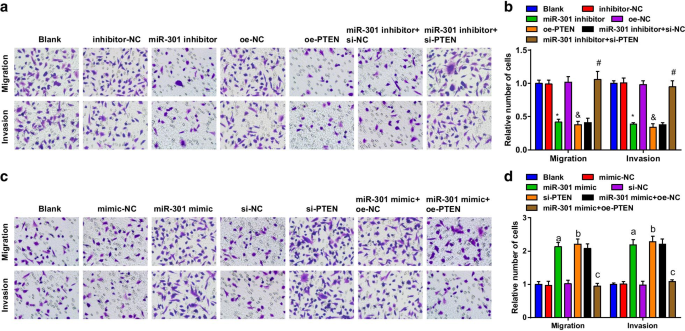
Ингибированный miR-301 или сверхэкспрессированный PTEN подавляет миграцию и инвазию клеток ESCC; повышенный miR-301 или пониженный PTEN способствует миграции и инвазии клеток ESCC. а Способность к миграции и инвазии трансфицированных клеток KYSE-30, оцениваемая с помощью анализа Transwell, после подавления miR-310 или активации PTEN; б статистические результаты мигрировавших и инвазивных клеток KYSE-30 с помощью анализа Transwell после подавления miR-310 или активации PTEN; c способность клеток Eca109 к миграции и инвазии среди групп, оцениваемых с помощью анализа Transwell, после активации miR-310 или подавления PTEN; г статистические результаты мигрировавших и инвазивных клеток Eca109 с помощью анализа Transwell с активацией miR-310 или подавлением PTEN. * P <0,05 по сравнению с группой ингибитор-NC; & P <0,05 по сравнению с группой oe-NC; # P <0,05 по сравнению с группой ингибитор miR-301 + si-NC; а P <0,05 по сравнению с группой мимик-NC; б P <0,05 по сравнению с группой si-NC; c P <0,05 по сравнению с группой миметиков miR-301 + oe-NC, N =3. Данные были выражены как среднее ± стандартное отклонение, и ANOVA использовался для сравнения между несколькими группами
Ингибирование miR-301 или сверхэкспрессия PTEN вызывает остановку клеточного цикла и апоптоз клеток ESCC; Повышенный уровень miR-301 или снижение PTEN подавляет остановку клеточного цикла и апоптоз клеток ESCC
Проточная цитометрия использовалась для обнаружения перехода клеточного цикла и апоптоза клеток после трансфекции, и результаты показали, что в клеточной линии KYSE-30 (рис. 5a-d) трансфекция ингибитором miR-301 или oe-PTEN способствовала скорости апоптоза. и увеличилось количество клеток в фазе G0 / G1, тогда как уменьшилось в фазах S и G2 / M; изменение апоптоза и остановки клеточного цикла, вызванное ингибитором miR-301, может быть обращено si-PTEN.
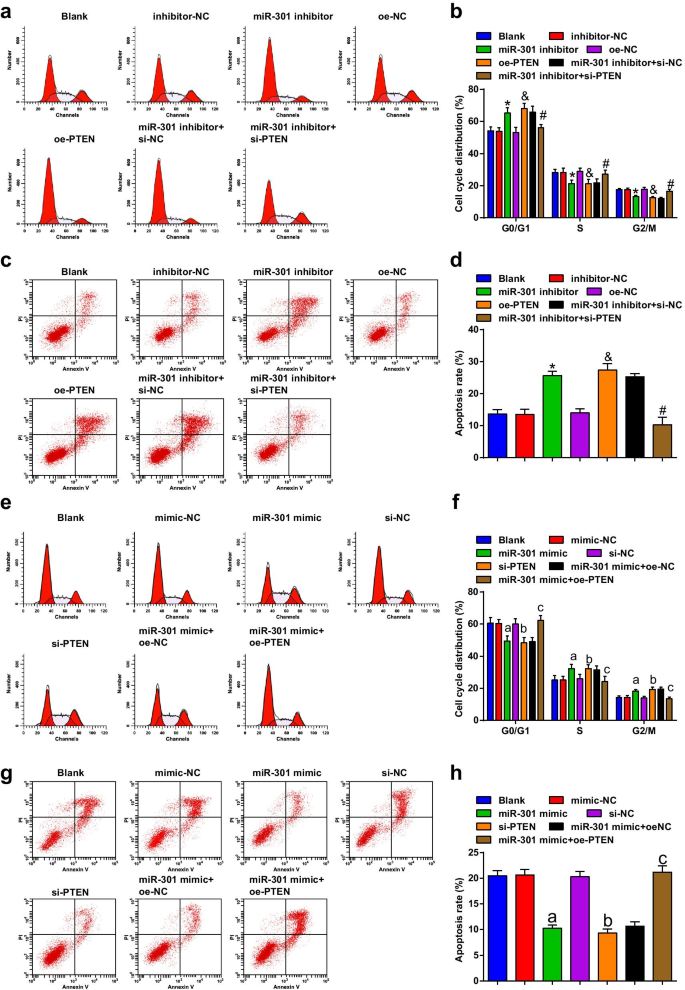
Ингибирование miR-301 или сверхэкспрессия PTEN вызывает остановку клеточного цикла и апоптоз клеток ESCC; повышенный miR-301 или пониженный PTEN подавляет остановку клеточного цикла и апоптоз клеток ESCC. а Распределение клеточного цикла клеток KYSE-30 в каждой группе определяли с помощью проточной цитометрии после подавления miR-310 или усиления PTEN; б статистические результаты процентного содержания в фазах G0 / G1, S и G2 / GM клеток KYSE-30 при проточной цитометрии после подавления miR-310 или усиления PTEN; c апоптоз клеток KYSE-30 был обнаружен с помощью проточной цитометрии после подавления miR-310 или усиления PTEN; г скорость апоптоза трансфицированных клеток KYSE-30, определенная с помощью проточной цитометрии после подавления miR-310 или усиления PTEN; е распределение клеток Eca109 в клеточном цикле в каждой группе определяли с помощью проточной цитометрии с помощью активации miR-310 или подавления PTEN; е статистические результаты процентного содержания в фазах G0 / G1, S и G2 / GM клеток Eca109 при проточной цитометрии после активации miR-310 или подавления PTEN; г апоптоз клеток Eca109 был обнаружен с помощью проточной цитометрии после активации miR-310 или подавления PTEN; ч Скорость апоптоза трансфицированных клеток Eca109, определенная с помощью проточной цитометрии после активации miR-310 или подавления PTEN. * P <0,05 по сравнению с группой ингибитор-NC; & P <0,05 по сравнению с группой oe-NC; # P <0,05 по сравнению с группой ингибитор miR-301 + si-NC; а P <0,05 по сравнению с группой мимик-NC; б P <0,05 по сравнению с группой si-NC; c P <0,05 по сравнению с группой миметиков miR-301 + oe-NC, N =3. Данные были выражены как среднее ± стандартное отклонение, и ANOVA использовался для сравнения между несколькими группами
По результатам проточной цитометрии мы обнаружили, что в клеточной линии Eca109 (рис. 5e-h) трансфекция миметика miR-301 или si-PTEN ингибировала скорость апоптоза, уменьшала количество клеток в фазе G0 / G1 и увеличивала скорость апоптоза. Фаза S и фазы G2 / M; Сверхэкспрессия PTEN обращала эффект миметика miR-301 на скорость апоптоза и остановку клеточного цикла клеток Eca109. На основании этих результатов мы пришли к выводу, что подавление miR-301 или усиление PTEN способствовало переходу клеточного цикла и апоптозу в клетках ESCC, тогда как подавление miR-301 или подавление PTEN оказывало противоположные эффекты.
Ингибирование miR-301 или сверхэкспрессия PTEN сдерживает рост опухоли и ангиогенез in vivo в ESCC; Повышенный уровень miR-301 или снижение PTEN увеличивает рост опухоли и ангиогенез in vivo в ESCC
В каждой группе наблюдали рост и изменения опухолей ESCC у мышей nude. Был оценен рост опухоли, и результаты показали, что в клеточной линии KYSE-30 (фиг. 6a-e) голые мыши, которым вводили ингибитор miR-301 или oe-PTEN, уменьшали объем и вес опухоли; Репрессивная роль ингибитора miR-301 в росте опухоли отменяется si-PTEN. В клеточной линии Eca109 (рис. 6f – j) объем опухоли и масса опухоли увеличивались у бестимусных мышей, которым вводили миметик miR-301 или si-PTEN; сверхэкспрессия PTEN обращала эффект миметика miR-301 на рост опухоли. Между тем, экспрессия CD34 в ксенотрансплантатах от мышей nude оценивалась с помощью иммуногистохимического окрашивания, и результаты показали, что (рис. 7a-d) в ксенотрансплантатах KYSE-30 MVD сдерживался после подавления miR-301 или усиления PTEN; заглушенный PTEN обращал действие ингибирования miR-301 на MVD. В ксенотрансплантатах Eca109 MVD увеличивался после активации miR-301 или подавления PTEN; усиление MVD, индуцированное повышающей регуляцией miR-301, может быть отменено сверхэкспрессией PTEN. These data indicated that miR-301 inhibition or PTEN overexpression repressed tumor growth and angiogenesis in ESCC, while miR-301 elevation or PTEN silencing had reverse effects.

Inhibited miR-301 or overexpressed PTEN restrains tumor growth in ESCC; elevated miR-301 or reduced PTEN increases tumor growth in ESCC. а Representative figures for the tumor growth observed by subcutaneous tumorigenesis in nude mice after KYSE-30 cells were transfected; b–d changes of tumor volume of each group after KYSE-30 cells were transfected; е changes of tumor weight of each group after KYSE-30 cells were transfected; е representative figures for the tumor growth observed by subcutaneous tumorigenesis in nude mice after Eca109 cells were transfected; g–i changes of tumor volume of each group after Eca109 cells were transfected; j changes of tumor weight of each group after Eca109 cells were transfected. * P < 0.05 versus the inhibitor-NC group; &P < 0.05 versus the oe-NC group; # P < 0.05 versus the miR-301 inhibitors + si-NC group; a P < 0.05 versus the mimic-NC group; b P < 0.05 versus the si-NC group; c P < 0.05 versus the miR-301 mimic + oe-NC group, n = 3 mice. Data were expressed as mean ± standard deviation, and ANOVA was used for comparisons among multiple groups

Inhibited miR-301 or overexpressed PTEN restrains angiogenesis in ESCC; elevated miR-301 or reduced PTEN increases angiogenesis in ESCC. а Representative images of tumor tissues observed by immunohistochemical staining in nude mice after KYSE-30 cells were transfected; б comparisons of MVD of KYSE-30 in tumor tissues among the groups; c representative images of tumor tissues observed by immunohistochemical staining in nude mice after Eca109 cells were transfected; г comparisons of MVD of Eca109 in tumor tissues among the groups *P < 0.05 versus the inhibitor-NC group; &P < 0.05 versus the oe-NC group; # P < 0.05 versus the miR-301 inhibitors + si-NC group; a P < 0.05 versus the mimic-NC group; b P < 0.05 versus the si-NC group; c P < 0.05 versus the miR-301 mimic + oe-NC group, n = 3 mice. Data were expressed as mean ± standard deviation, and ANOVA was used for comparisons among multiple groups
Обсуждение
EC is a kind of invasive malignancy in the gastrointestinal tract [16]. As the major type of EC, ESCC is a malignant tumor occurring in esophageal epithelial cells [17]. The miRNAs, known as small non-coding RNAs, have been demonstrated to function as a significant roles in leading molecules in the silencing of RNA [18]. Our research was designed to explore the effects of miR-301 and its target gene PTEN on ESCC progression, and we have found that the inhibited miR-301 could suppress angiogenesis and cell growth in ESCC by elevating PTEN.
MiR-301 expression was assessed, and we found that miR-301 was highly expressed in ESCC cells in comparison with HEEC, and the higher expression of miR-301 has also been found in ESCC tissues in contrast to the adjacent normal tissues. Similar to this result, Li et al. have identified that miR-301 presented high expression in myocardial infarction tissues [19]. In addition, we have elucidated that PTEN was targeted by miR-301, and the target relation has been pointed out by an extant literature [20]. We have also discovered that PTEN, which has been affirmed to be targeted by miR-301, was downregulated in both ESCC tissues and cells. Similarly, a previous research has unearthed that PTEN was poorly expressed in ESCC compared with non-tumor esophageal epithelial tissue [21]. Furthermore, Ma et al. have illuminated that PTEN expression was degraded in Eca109 cell line [22], which has also been selected for a series of experiments in this research. These studies provide evidence for the high expression of miR-301 and low expression of PTEN in ESCC.
Another important outcome in this research indicated that the inhibited miR-301 could repress the colony formation ability as well as the cell proliferation of ESCC cells via enhancing the PTEN expression, and elevated miR-301 or reduced PTEN had contrary effects. Similarly, Han et al. have elucidated that the downregulation of miR-301 mediated by luteolin has the ability to restrain the cell proliferation in prostate cancer [6]. A recent literature has revealed that the overexpression of PTEN suppresses the proliferation of pancreatic cancer cells [23], and a same result has been summarized in a study focusing on prostate cancer [24]. Besides, we have also unearthed that the downregulation of miR-301 or the elevation of PTEN could inhibit migration and invasion of ESCC cells, and elevated miR-301 or reduced PTEN exhibited the opposite trends. In accordance with this outcome, Shi et al. have supported that inhibited miR-301 attenuated migration and invasion of breast cancer cells [10], and it has been reported that the migration and invasion of ESCC cells could be repressed by the inhibition of miR-130b and the elevation of PTEN [25]. These publications helped verifying the oncogenic role of miR-301 and tumor-repressive effect of PTEN in diverse human cancers. Another result in our research was that inhibited miR-301 overexpressed PTEN to promote cell apoptosis and induce cell cycle arrest at the G0/G1 phase in ESCC cells, and elevated miR-301 or reduced PTEN had the inverse results. Similarly, it has been uncovered by a recent literature that activated PTEN induces cell cycle arrest and apoptosis in ESCC [26]. Consistently, Tian et al. have found in their study that the elevation of PTEN inhibited the angiogenesis by reducing the expression of vascular endothelial growth factor in hepatocellular carcinoma [27]. Based on the above data, the roles of miR-301 and PTEN in cell apoptosis and angiogenesis in diverse diseases were further confirmed. Consequently, we concluded that the downregulation of miR-301 could restrain the tumor growth in ESCC through the high expression of PTEN, and the similar conclusion has also been unveiled in breast cancer [10] and prostate cancer [28]. On the contrary, miR-301 elevation or PTEN reduction induced the tumor growth in ESCC. It could be concluded that miR-301 and PTEN participated in the in vivo cancer cell growth.
Conclusion
In this study, we have shown that the repression of miR-301 prohibits angiogenesis, cell proliferation, migration and invasion but promotes apoptosis in ESCC cells by upregulating PTEN. This research may further the understanding on potential molecular mechanisms of ESCC and provide novel targets for ESCC treatment.
Сокращения
- ESCC:
-
Плоскоклеточный рак пищевода
- PTEN:
-
Phosphatase and tensin homologue
- MVD:
-
Microvessel density
- EC:
-
Esophageal cancer
- miRNAs:
-
MicroRNAs
- LNM:
-
Lymph node metastasis
- UICC:
-
Union for International Cancer Control
- RT-qPCR:
-
Reverse transcription quantitative polymerase chain reaction
- 3′UTR:
-
3′-Untranslated region
- WT:
-
Wild type
- MUT:
-
Mutant type
- HRP:
-
Пероксидаза хрена
- FBS:
-
Фетальная бычья сыворотка
- OE:
-
Overexpressed
- NC:
-
Negative control
- CCK-8:
-
Cell counting kit
- DMEM:
-
Dulbecco’s modified Eagle medium
- PI:
-
Иодид пропидия
- FITC:
-
Fluorescein isothiocyanate
- MVD:
-
Microvessel density
- ANOVA:
-
Analysis of variance
Наноматериалы
- Понижающая регуляция микроРНК-342-5p или повышающая регуляция Wnt3a ингибирует ангиогенез и поддерживает стабиль…
- Получение наночастиц mPEG-ICA, нагруженных ICA, и их применение в лечении LPS-индуцированного повреждения клеток H9c2…
- Анализ организации актина и фокальной адгезии в клетках U2OS на полимерных наноструктурах
- Иерархические антибактериальные полиамидные 6-ZnO нановолокна, полученные путем осаждения атомных слоев и гид…
- Изучение in vitro влияния наночастиц Au на клеточные линии HT29 и SPEV
- Стимуляция роста клеток SH-SY5Y наночастицами золота, модифицированными 6-меркаптопурином и проникающим в нейро…
- Наночастицы маггемита действуют как нанозимы, улучшая рост и устойчивость к абиотическому стрессу у Brassica napus
- Золотые наночастицы круглой формы:влияние размера и концентрации частиц на рост корней Arabidopsis thaliana
- Поддержка новых анодных катализаторов для топливных элементов с прямым метанолом:характеристики и характер…
- 5G и проблема экспоненциального роста данных