


SOX2 регулирует ось lncRNA CCAT1 / MicroRNA-185-3p / FOXP3, чтобы влиять на пролиферацию и самообновление стволовых клеток рака шейки матки
Аннотация
Была представлена роль длинных некодирующих РНК (lncRNAs) при раке шейки матки (CC). Мы стремимся обсудить влияние определяющей пол области Y-box 2 (SOX2) / lncRNA, транскрипта-1, связанного с раком толстой кишки (CCAT1) / microRNA-185-3p (miR-185-3p) / белка 3 forkhead box (FOXP3). ) на пролиферацию и способность к самообновлению стволовых клеток CC. Экспрессию MiR-185-3p, SOX2, CCAT1 и FOXP3 тестировали в тканях и клетках CC. Была подтверждена взаимосвязь между экспрессией SOX2 / CCAT1 и клинико-патологическими особенностями у пациентов с РС. Исследования потери и увеличения функции были проведены в CD44 + Клетки HeLa для обсуждения биологических функций и способности к самообновлению. Наконец, были подтверждены отношения между SOX2, CCAT1, FOXP3 и miR-185-3p. Экспрессия miR-185-3p была снижена, тогда как экспрессия SOX2, CCAT1 и FOXP3 была увеличена в тканях и клетках CC. Экспрессия SOX2 и CCAT1 была связана с размером опухоли, метастазами в лимфатические узлы и стадией CC Международной федерации гинекологии и акушерства. Подавляющая регуляция SOX2 или CCAT1 и повышающая регуляция miR-185-3p приводили к ингибированию пролиферации, инвазии, миграции и числа клеточных сфер, а также к ускорению апоптоза CD44 + Клетки HeLa. SOX2 может связываться с CCAT1, что влияет на экспрессию miR-185-3p, а на FOXP3 нацелен miR-185-3p.
Введение
Рак шейки матки (РШМ) является четвертой по значимости причиной смертности среди женщин:в 2018 г. во всем мире было зарегистрировано 570 000 случаев заболевания и 311 000 смертей [1]. В этом сложном заболевании участвует множество факторов, включая генетические эффекты и вирусную инфекцию [2]. С развитием совместного тестирования на вирус папилломы человека и вакцинации против вируса папилломы человека ранние диагностические процедуры дисплазии шейки матки и рака приводят к снижению заболеваемости, заболеваемости и смертности от CC [3]. Пациентам на ранней стадии ХК рекомендуется хирургическое вмешательство, такое как операция по сохранению фертильности, конусная биопсия, радикальная трахелэктомия, диссекция тазовых лимфатических узлов, лучевая терапия и брахитерапия [4]. Прогноз по-прежнему плохой из-за метастазирования или рецидива у пациентов с далеко зашедшим ХК [5]. Следовательно, по-прежнему актуальна идентификация новых и эффективных прогностических маркеров и терапевтических стратегий для улучшения лечения CC.
Определяющая пол область Y-box 2 (SOX2) является критическим членом семейства транскрипционных факторов SOX и в основном проявляется в эмбриональных и взрослых стволовых клетках, а также экспрессируется в стволовых клетках опухолей [6]. Было обнаружено, что SOX2 модулирует радиорезистентность в CC посредством сигнального пути hedgehog [7]. Другое исследование продемонстрировало, что SOX2 имеет решающее значение для поддержания субпопуляции раковых стволовых клеток в линиях клеток CC [8]. Длинные некодирующие РНК (днРНК) представляют собой класс молекул РНК с длиной 200 нуклеотидов [9]. Транскрипт-1, связанный с раком толстой кишки, LncRNA (CCAT1) расположен на хромосоме 8q24.21 человека и рассматривается как «горячая точка», которая приводит к генетическим мутациям при раке толстой кишки [10]. В исследовании сообщается, что CCAT1 ускоряет пролиферацию клеток и инвазию CC [11]. Согласно Jia et al., CCAT1 резко усиливает пролиферацию, миграцию и инвазию CC клеток [12]. Более того, другое исследование показало, что CCAT1 увеличивает степень злокачественности воспалительного заболевания кишечника за счет разрушения кишечного барьера за счет уменьшения количества микроРНК-185-3p (miR-185-3p) [13]. MiRNA может обратно контролировать экспрессию гена за счет уменьшения мРНК и подавления трансляции [14]. Эксперименты in vitro в предыдущем исследовании показали, что miR-185-3p модулирует радиорезистентность карциномы носоглотки [15]. Другое исследование показало, что miR-185 участвует в устойчивости рака яичников к цисплатину in vivo и in vitro [16]. Forkhead box protein 3 (FOXP3) представляет собой фактор транскрипции, принадлежащий к семейству белков FOX, который впервые обнаружен в регуляторных T (Treg) -клетках и играет жизненно важную роль в поддержании и функционировании Treg-клеток [17]. В исследовании сообщается, что FOXP3 связан с лимфангиогенезом CC [18]. Другое исследование показывает, что уровень FOXP3 напрямую связан со стадией акушерства и гинекологии (стадия FIGO) и размером опухоли CC [19]. Поэтому в этом исследовании мы изучили влияние оси SOX2 / CCAT1 / miR-185-3p / FOXP3 на пролиферацию и способность к самообновлению стволовых клеток CC.
Материалы и методы
Утверждение этических норм и согласие на участие
Эксперименты с участием людей проводились в соответствии с принципами, изложенными в Хельсинкской декларации. Исследование было одобрено институциональным наблюдательным советом Первой больницы Цзилиньского университета. Все участники подписали документ об информированном согласии.
Темы исследования
С декабря 2016 года по декабрь 2018 года у пациентов с РС были взяты 39 образцов тканей CC и соответствующих прилегающих нормальных тканей, которые сохранялись в жидком азоте. Критерии включения были следующими:(1) Пациенты были подтверждены как CC по патологии биопсии трансцервикального и цервикального каналов, цитологии цервикального мазка, цервикальному йодному тесту, вагинальному зеркалу и резекции шейного позвонка. (2) Пациенты не получали лучевую терапию и химиотерапию за 2 недели до операции. Критерии исключения были следующими:(1) пациенты, проходящие любую лучевую терапию или химиотерапию, (2) пациенты, не согласные с забором образца и (3) пациенты с нарушениями иммунной системы.
Выбор клеток и культура
Линии клеток СС (SiHa, HeLa, CaSki, HCC94 и C33A) и линия иммортализованных клеток шейного эпителия человека H8 были приобретены у Shanghai Bioleaf Biotech Co., Ltd. (Шанхай, Китай). Клетки СС SiHa, HeLa и HCC94 культивировали в среде Игла с высоким содержанием глюкозы, модифицированной Дульбекко (DMEM), содержащей 10% фетальной телячьей сыворотки (FBS), тогда как клетки CaSki, C33A и H8 в среде Roswell Park Memorial Institute (RPMI) -1640, содержащей 10% % FBS (37 ℃ и 5% CO 2 ). Клетки отделяли и пересевали каждые 2 дня.
Сортировка стволовых клеток CC (CD44 + HeLa Cells)
Стволовые клетки СС отделяли от линии клеток СС HeLa культивированием клеточной суспензии. CC-клетки HeLa культивировали бессывороточной модифицированной средой Дульбекко Искова (IMDM) в чашке Петри со сверхнизкой адгезией в течение 21 дня с полуколичественной заменой среды каждые 3-5 дней. Некоторые клетки суспендировали в сферы, и получали клетки, образующие сферы HeLa (SFC). Выявлено и проанализировано свойство сферических клеток. Клетки были отделены трипсином и доведены до 1 × 10 6 клеток / мл. К клеткам добавляли антитело против CD44 и сортировали методом проточной цитометрии. Клетки HeLa с положительным CD44 были опухолевыми стволовыми клетками HeLa, а с отрицательным CD44 - не стволовыми клетками HeLa. Стволовые клетки CC культивировали в среде DMEM / F12 и добавляли 20 нг / мл основного фактора роста фибробластов (bFGF), 20 нг / мл эпидермального фактора роста (EGF) и B27. Среда содержала 1% пенициллина и стрептомицина [20].
Обработка клеток
CD44 + Клетки HeLa трансфицировали sh-SOX2, sh-SOX2 отрицательным контролем (NC), sh-CCAT1, sh-CCAT1 NC, миметиком miR-185-3p, миметиком NC, sh-CCAT1 и ингибитором miR-185-3p, а также ингибитором miR-185-3p. sh-CCAT1 и ингибитор NC. Все олигонуклеотидные последовательности были предоставлены GenePharma (Шанхай, Китай). Отделенные трипсином клетки высевали в 6-луночный планшет с 3 × 10 6 ячеек / лунка. При достижении 60% конфлюэнтности клетки заменяли на бессывороточную среду и инкубировали в течение 1 часа. Трансфекцию способствовал реагент для трансфекции Lipofectamine 2000 (Invitrogen, Carlsbad, California, USA).
Количественная полимеразная цепная реакция с обратной транскрипцией (RT-qPCR)
РНК в тканях и клетках экстрагировали Trizol (Invitrogen). РНК (1 мкг) превращали в кДНК с помощью набора RTase вируса мышиного лейкоза Молони (Invitrogen). кДНК добавляли в систему ПЦР в реальном времени. Праймеры были разработаны Shanghai Sangon Biotechnology Co., Ltd. (Шанхай, Китай) (таблица 1). U6 был контролем загрузки miR-185-3p, тогда как глицеральдегидфосфатдегидрогеназа (GAPDH) SOX2, CCAT1 и FOXP3. Результаты были проанализированы с помощью 2 −ΔΔCt метод.
Вестерн-блоттинг
Общий белок в клетках и тканях был экстрагирован. Концентрацию белка определяли с помощью наборов бицинхониновой кислоты (AmyJet Scientific, Ухань, Хубэй, Китай). Белок смешивали с загрузочным буфером и кипятили в течение 5 минут с последующей ледяной баней и центрифугированием. Белок обрабатывали электрофорезом в 10% додецилсульфат натрия в полиакриламидном геле и переносили на мембрану. Мембрану блокировали 5% обезжиренным молоком в течение 1 ч, исследовали первичными антителами SOX2 (1:1000, Jiangsu Rui sitan Co., Ltd., Цзянсу, Китай), FOXP3 (1:1000, Abcam Inc., Кембридж, Массачусетс). , США), GAPDH (1:1000, Cell Signaling Technology, Беверли, Массачусетс, США) и повторно зондировали вторичным антителом, меченным пероксидазой хрена, покрытым консервирующей пленкой и экспонированным. В разработку был принят имидж-сканер Bio-rad Gel Doc EZ (Bio-rad, Калифорния, США). Изображения белков анализировали с помощью программного обеспечения ImageJ2x.
Набор для подсчета клеток (CCK) -8 Assay
Анализ CCK-8 проводили с наборами (Beyotime, Шанхай, Китай). Ячейки (1 × 10 4 ) высевали в 96-луночный планшет и инкубировали. Культивировали в течение 0, 24, 48 и 72 часов, в клетки добавляли 10 мкл / лунку раствора CCK-8 и вылупляли в течение 1 часа. Значение оптической плотности определяли с помощью полноволнового ридера для микропланшетов Multiskan Spectrum при 450 нм. Для подсчета среднего значения было взято шесть лунок. Кривую роста клеток строили в зависимости от времени по оси ординат и относительной жизнеспособности клеток по оси ординат. Значение оптической плотности отражает пролиферацию клеток.
Проточная цитометрия
Ячейки (1 × 10 6 ) центрифугировали при 1500 об / мин, суспендировали с 200 мкл связывающего буфера, инкубировали с 5 мкл йодида пропидия (PI) и 5 мкл аннексина V-флуоресцеина изотиоцианата (FITC) по очереди и добавляли 400 мкл буфера. Скорость апоптоза клеток проверяли с помощью проточного цитометра (BD Biosciences, Нью-Джерси, США).
Скретч-тест
Трансфицированные клетки отделяли и готовили суспензию клеток. Суспензии клеток (1 × 10 6 клеток / мл) высевали в 6-луночный планшет и культивировали до 80–90% конфлюэнтности. Поперечные и продольные царапины на 6-луночном планшете были равномерно нанесены кончиком асептической пипетки на 10 мкл вдоль линейки. Исходную среду заменяли полной средой, и клетки непрерывно культивировали. Миграцию клеток наблюдали под микроскопом через 0 и 72 часа и отображали в том же месте.
Анализ Transwell
Трансфицированные клетки трипсинизировали, центрифугировали при 1000 g и промывали DMEM. Полностью гидратированную камеру Transwell помещали в 24-луночный планшет, содержащий 10% FBS-DMEM (600 мл / лунку). Клеточная суспензия (5 × 10 5 клеток / мл, 200 мкл) добавляли в верхнюю камеру Transwell (покрытую матригелем 1:8 80 мкл), а 500 мкл 20% FBS-DMEM - в нижнюю камеру. Клетки непрерывно культивировали в течение 24 ч, фиксировали 500 мкл 4% параформальдегида (PFA) и окрашивали 0,1% раствором для окрашивания кристаллическим фиолетовым. Затем клетки на поверхности и краю верхней камеры протирали ватным тампоном. Случайным образом было выбрано пять полей, и клетки были подсчитаны под микроскопом Nikon Eclipse TE2000-S (Nikon, Япония).
Анализ сферообразования
Клетки высевали в 6-луночный планшет с низкой адсорбцией, содержащий бессывороточную суспензионную среду из расчета 200 клеток / лунку. Через 2 недели скорость образования сфер из клеток наблюдали под микроскопом Nikon Eclipse TE2000-S (Nikon), и скорость образования сфер рассчитывалась как среднее количество сфер / количество засеянных клеток × 100%.
Анализ иммунопреципитации хроматина (ChIP)
ChIP-анализ проводили с помощью набора ChIP (Upstate, NY, USA). SOX2 (1:1000, Re-stem Biotech, Цзянсу, Китай) и нормальные кроличьи IgG (12–370, Millipore, США) использовали для иммунопреципитации сшитого комплекса белок-ДНК. CD44 + Клетки HeLa фиксировали 1% PFA и инкубировали для образования перекрестных связей ДНК-белок. Затем ДНК разрезали на фрагмент хроматина размером 200–300 п.н. с помощью ультразвука. Осажденную ДНК хроматина выделяли и анализировали с помощью RT-qPCR.
Анализ РНК с понижением давления
Меченные биотином плазмиды miR-185-3p дикого типа (WT) и мутантного типа (MUT) (50 нМ) трансфицировали в CD44 + Клетки HeLa соответственно. Клетки вылупляли специфическим клеточным лизатом (Ambion, Остин, Техас, США) через 48 часов трансфекции. Лизат клеток (50 мл) был упакован. Из остаточного лизата наносили гранулы стрептавидина М-280 (Sigma, Сент-Луис, Миссури, США), предварительно покрытые не содержащей РНКаз и дрожжевой тРНК (Sigma). После этого клетки дважды очищали холодным лизатом, трижды - буфером с низким содержанием соли и один раз - буфером с высоким содержанием соли. Антагонистический зонд miR-185-3p был настроен как NC. Общую РНК экстрагировали с помощью Trizol, а уровень CCAT1 проверяли с помощью RT-qPCR.
Анализ двойного люциферазного репортерного гена
Возможные сайты связывания E1 и E2 SOX2 на промоторной области CCAT1 были предсказаны https://jaspar.genereg.net/. Была синтезирована промоторная последовательность CCAT1, содержащая SOX2 и сайт связывания CCAT1 E1, и были сформированы векторы CCAT1 3′UTR WT (E1-WT) и CCAT1 3′UTR MUT (E1-MUT). Векторы клонировали в pmirGLO (Beyotime). После этого CCAT1-WT / pmirGLO или CCAT1-MUT / pmirGLO котрансфицировали с sh-SOX2 или sh-SOX2 NC в CD44 + Клетки HeLa соответственно в течение 2 дней, а затем лизируются. Активность люциферазы проверяли с помощью системы обнаружения люциферазы (Такара, Далянь, Китай).
Веб-сайт биоинформатики использовался для прогнозирования и анализа сайтов связывания CCAT1 и miR-185-3p. Сайты связывания CCAT1 и miR-185-3p проверяли с помощью анализа двойного люциферазного репортерного гена. Был составлен CCAT1 3'UTR, содержащий сайт связывания miR-185-3p. CCAT1 3′UTR WT и CCAT1 3′UTR MUT были сконструированы и котрансфицированы миметиками NC и miR-185-3p в CD44 + Клетки HeLa на 2 дня. Затем клетки лизировали и проверяли активность люциферазы с помощью системы обнаружения люциферазы (Takara). Тот же метод был применен для проверки связи нацеливания между miR-185-3p и FOXP3.
Статистический анализ
Все данные были оценены с помощью программного обеспечения SPSS 21.0 (IBM Corp. Armonk, NY, США). Данные измерений были указаны как среднее ± стандартное отклонение. Тест Стьюдента применялся для несоответствия между двумя группами и однофакторного дисперсионного анализа (ANOVA) с последующим тестом множественных сравнений Тьюки на несоответствие между группами. Классификационная переменная оценивалась точным критерием Фишера. А п значение менее 0,05 считалось значимым.
Результаты
Экспрессия miR-185-3p снижается, в то время как экспрессия SOX2, CCAT1 и FOXP3 увеличивается в тканях CC, а экспрессия SOX2 и CCAT1 связана с размером опухоли, лимфатическим узлом Метастазирование (LNM) и стадия FIGO
При обнаружении роли оси SOX2 / CCAT1 / miR-185-3p / FOXP3 в пролиферации и способности к самообновлению стволовых клеток CC, экспрессия miR-185-3p, SOX2, CCAT1 и FOXP3 в тканях CC и прилегающих нормальных тканях была протестировано с помощью RT-qPCR и вестерн-блоттинга. Было показано, что (рис. 1a – c) экспрессия miR-185-3p была снижена, в то время как экспрессия SOX2, CCAT1 и FOXP3 была увеличена в тканях CC.
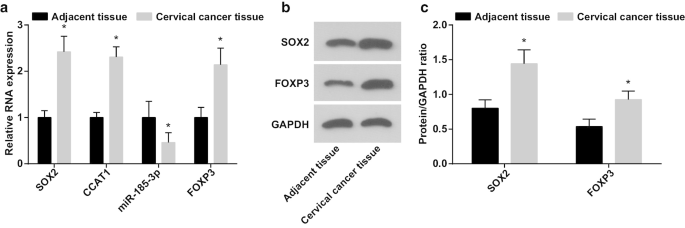
Экспрессия miR-185-3p снижается, тогда как экспрессия SOX2, CCAT1 и FOXP3 увеличивается в тканях CC. а Сравнение экспрессии мРНК SOX2, CCAT1, miR-185-3p и FOXP3 в CC и прилегающих нормальных тканях. б Белковые полосы экспрессии белков SOX2 и FOXP3 в CC и прилегающих нормальных тканях. c Сравнение экспрессии белков SOX2 и FOXP3 в CC и прилегающих нормальных тканях. * p <0,05 по сравнению с соседними нормальными тканями. Данные измерений были представлены как среднее значение ± стандартное отклонение, а сравнения между двумя группами оценивались с помощью t-критерия
Была проанализирована взаимосвязь между экспрессией SOX2 / CCAT1 и клинико-патологическими особенностями CC (Таблица 2). Сверхэкспрессия SOX2 и CCAT1 в CC были связаны с размером опухоли, LNM и стадией FIGO, что указывает на то, что экспрессия SOX2 и CCAT1 была выше у пациентов с большим размером опухоли, LNM и продвинутой стадией FIGO у пациентов с CC.
Экспрессия miR-185-3p уменьшается, тогда как экспрессия SOX2, CCAT1 и FOXP3 увеличивается в CD44 + Клетки HeLa
Затем были протестированы экспрессии miR-185-3p, SOX2, CCAT1 и FOXP3 в иммортализованных клетках шейного эпителия человека H8 и клеточных линиях CC SiHa, CasKi, HeLa, HCC94 и C33A. Было высказано предположение, что (Fig. 2a-c) экспрессия miR-185-3p деградировала, тогда как SOX2, CCAT1 и FOXP3 были повышены в линиях клеток CC. Среди них SOX2 / CCAT1 / miR-185-3p / FOXP3 в клетках HeLa имел наибольшее отличие экспрессии от клеток H8; таким образом, клетки HeLa были отсортированы; HeLa-SFC были получены с помощью культур суспензий клеток, а экспрессия CD44 на поверхности клеток до и после сортировки была идентифицирована с помощью проточной цитометрии. Результаты подчеркнули, что количество положительных CD44 в HeLa-SFC было заметно выше, чем до сортировки, что свидетельствует об успешной сортировке стволовых клеток CC (рис. 2d). Отсортированные стволовые клетки были названы CD44 + . Клетки HeLa. Затем экспрессии miR-185-3p, SOX2, CCAT1 и FOXP3 были протестированы в HeLa и CD44 + Клетки HeLa (рис. 2д – ж). Было продемонстрировано, что экспрессия SOX2, CCAT1 и FOXP3 активизировалась, а miR-185-3p подавлялась в CD44 + Клетки HeLa.
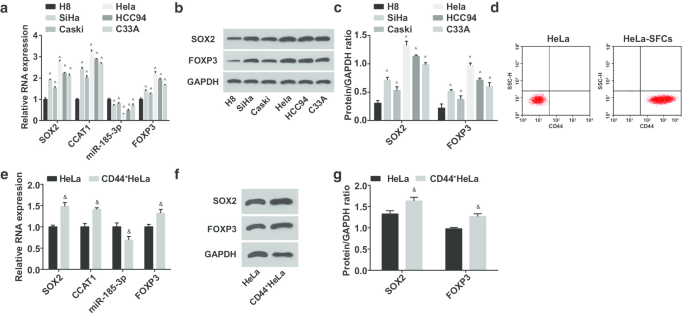
Экспрессия miR-185-3p снижается, в то время как экспрессия SOX2, CCAT1 и FOXP3 повышается в CD44 + Клетки HeLa. а Сравнение экспрессии мРНК SOX2, CCAT1, miR-185-3p и FOXP3 в линиях клеток H8 и CC. б Белковые полосы экспрессии белков SOX2 и FOXP3 в клеточных линиях H8 и CC. c Сравнение экспрессии белков SOX2 и FOXP3 в клеточных линиях H8 и CC. г Определение скорости экспрессии CD44 в клетках HeLa до и после сортировки с помощью проточной цитометрии. е Сравнение экспрессии мРНК SOX2, CCAT1 и miR-185-3p между клетками HeLa и CD44 + Клетки HeLa. е Белковая полоса экспрессии белка SOX2 в клетках HeLa и CD44 + Клетки HeLa. г Сравнение экспрессии белка SOX2 между клетками HeLa и CD44 + Клетки HeLa. ^ p <0,05 по сравнению с клетками H8. & p <0,05 по сравнению с клетками HeLa. Данные измерений были представлены как среднее значение ± стандартное отклонение, сравнения между двумя группами оценивались с помощью t-критерия, а сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа с последующим апостериорным тестом Тьюки
Понижающее регулирование SOX2 и понижающее регулирование CCAT1 снижают скорость распространения, миграции, вторжения и образования сфер, а также увеличивают апоптоз CD44 + Клетки HeLa
Впоследствии SOX2 и CCAT1 были заглушены в CD44 + Клетки HeLa, чтобы изучить их влияние на пролиферацию и самообновление стволовых клеток CC. Обнаруженные с помощью анализа CCK-8, проточной цитометрии, скретч-теста, анализа Transwell и эксперимента по формированию сфер, пролиферация, миграция, инвазия и скорость образования сфер были подавлены, а апоптоз CD44 + Клеткам HeLa способствовало ингибирование SOX2 и CCAT1 (рис. 3a – i). Это свидетельствует о том, что подавление SOX2 или CCAT1 ингибирует пролиферацию и самообновление стволовых клеток CC.
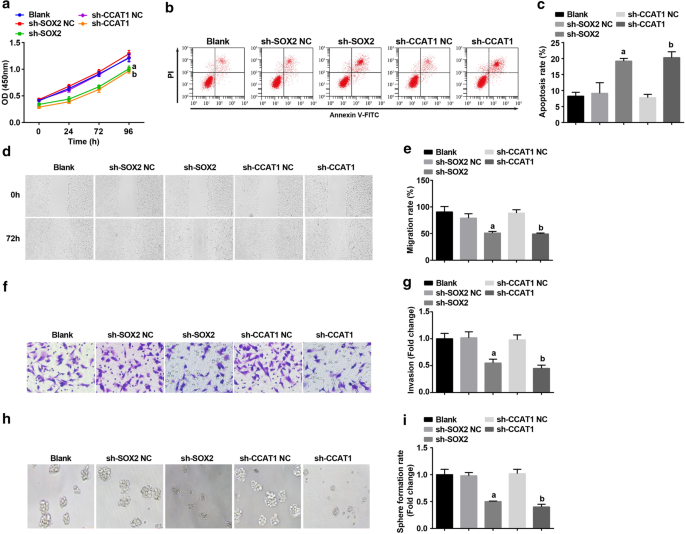
Подавление SOX2 и подавление CCAT1 снижает скорость пролиферации, миграции, инвазии и образования сфер, а также увеличивает апоптоз CD44 + Клетки HeLa. а Анализ CCK-8 проверял кривую роста клеток в клетках, обработанных sh-CCAT1 или sh-SOX2. б Проточная цитометрия выявила апоптоз клеток, обработанных sh-CCAT1 или sh-SOX2. c Сравнение скорости апоптоза клеток, обработанных sh-CCAT1 или sh-SOX2. г Миграция клеток в клетках, обработанных sh-CCAT1 или sh-SOX2, проверена методом царапания. е Сравнение миграции клеток в клетках, обработанных sh-CCAT1 или sh-SOX2. е Определение способности клеток к инвазии в клетках, обработанных sh-CCAT1 или sh-SOX2, с помощью анализа Transwell. г Сравнение способности клеток к инвазии, обработанных sh-CCAT1 или sh-SOX2. ч В эксперименте по формированию сфер проверялась способность к самообновлению клеток, обработанных sh-CCAT1 или sh-SOX2. я Сравнение скорости образования сфер в клетках, обработанных sh-CCAT1 или sh-SOX2. а p <0,05 по сравнению с группой sh-SOX2 NC. б p <0,05 по сравнению с группой sh-CCAT1 NC. Данные измерений были представлены в виде среднего значения ± стандартное отклонение, а сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа с последующим апостериорным тестом Тьюки
Истощенный miR-185-3p меняет роль понижающей регуляции CCAT1 в CD44 + Клетки HeLa
Затем мы исследовали, участвует ли miR-185-3p в процессе CCAT1, регулирующем пролиферацию и самообновление стволовых клеток CC. CD44 + HeLa трансфицировали миметиком miR-185-3p или котрансфицировали sh-CCAT1 и ингибитором miR-185-3p. Результаты показали, что повышающая регуляция miR-185-3p заметно снижает скорость пролиферации, миграции, инвазии и образования сфер, в то же время повышая скорость апоптоза CD44 + Клетки HeLa. Клетки, обработанные ингибитором miR-185-3p, могут изменить роль подавляемого CCAT1 в пролиферации, миграции, инвазии, апоптозе и образовании клеточных сфер CD44 + Клетки HeLa (рис. 4a – i).
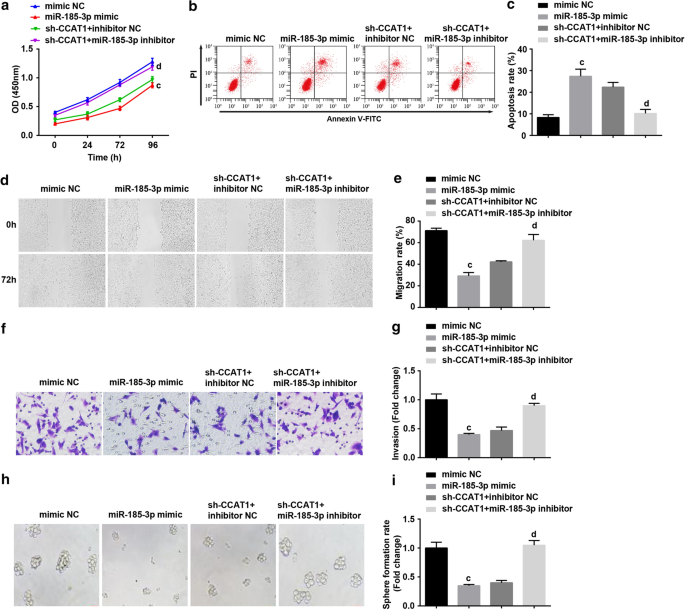
Сверхэкспрессия miR-185-3p подавляет скорость пролиферации, миграции, инвазии и образования сфер, а также увеличивает апоптоз CD44 + Клетки HeLa. а Анализ CCK-8 тестировал кривую роста клеток в клетках, обработанных миметиком miR-185-3p. б Проточная цитометрия выявила апоптоз клеток, обработанных миметиком miR-185-3p. c Сравнение скорости апоптоза клеток, обработанных миметиком miR-185-3p. г Миграцию клеток в клетках, обработанных миметиком miR-185-3p, проверяли с помощью скретч-теста. е Сравнение миграции клеток в клетках, обработанных миметиком miR-185-3p. е Обнаружение способности к инвазии клеток, обработанных миметиком miR-185-3p, с помощью анализа Transwell. г Сравнение инвазионной способности клеток, обработанных миметиком miR-185-3p. ч В эксперименте по формированию сфер проверялась способность к самообновлению клеток, обработанных миметиком miR-185-3p. я Сравнение скорости образования сфер в клетках, обработанных миметиком miR-185-3p. а п <0,05 по сравнению с группой sh-SOX2 NC. б р <0,05 по сравнению с группой sh-CCAT1 NC. c p <0,05 по сравнению с группой мимических NC. d p <0,05 по сравнению с группой NC с ингибитором sh-CCAT1 +. Данные измерений были представлены в виде среднего значения ± стандартное отклонение, а сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа с последующим апостериорным тестом Тьюки
Слабоэкспрессированный SOX2 и слабоэкспрессированный CCAT1 снижают экспрессию FOXP3 и увеличивают экспрессию miR-185-3p в CD44 + Клетки HeLa
После этого мы исследовали экспрессию SOX2 / CCAT1 / miR-185-3p / FOXP3 в CD44 + Клетки HeLa после трансфекции sh-SOX2, sh-CCAT1, miR-185-3p имитируют и котрансфекцию sh-CCAT1 и ингибитора miR-185-3p. Экспрессия SOX2, CCAT1 и FOXP3 была снижена, тогда как экспрессия miR-185-3p была повышена в клетках, обработанных sh-SOX2. Экспрессия CCAT1 и FOXP3 была снижена, а экспрессия miR-185-3p увеличена в клетках, обработанных sh-CCAT1. Экспрессия miR-185-3p была повышена, а экспрессия FOXP3 снижена в клетках, введенных с миметиком miR-185-3p. Экспрессия FOXP3 была повышена, а экспрессия miR-185-3p снижена в клетках, последовательно трансфицированных sh-CCAT1 и ингибитором miR-185-3p (рис. 5a – d).
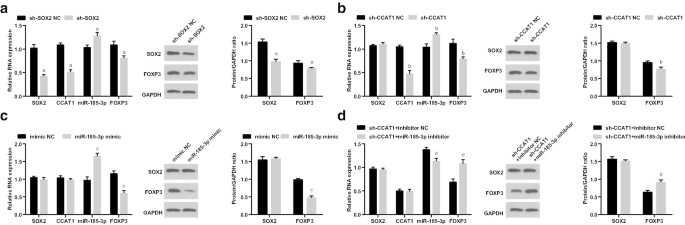
Низко экспрессируемый SOX2 и низко экспрессируемый CCAT1 снижают экспрессию FOXP3 и увеличивают экспрессию miR-185-3p в CD44 + Клетки HeLa. а Экспрессия SOX2 / CCAT1 / miR-185-3p / FOXP3 в группах sh-SOX2 NC и sh-SOX2. б Экспрессия SOX2 / CCAT1 / miR-185-3p / FOXP3 в группах sh-CCAT1 NC и sh-CCAT1. c Экспрессия SOX2 / CCAT1 / miR-185-3p / FOXP3 в мимических группах NC и miR-185-3p. г Экспрессия SOX2 / CCAT1 / miR-185-3p / FOXP3 в группах ингибитора sh-CCAT1 + NC и ингибитора sh-CCAT1 + miR-185-3p. а п <0,05 по сравнению с группой sh-SOX2 NC. б р <0,05 по сравнению с группой sh-CCAT1 NC. c p <0,05 по сравнению с группой мимических NC. d p <0,05 по сравнению с группой NC с ингибитором sh-CCAT1 +. Данные измерений были представлены в виде среднего значения ± стандартное отклонение, а сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа с последующим апостериорным тестом Тьюки
SOX2 связывается с CCAT1, который влияет на экспрессию miR-185-3p, а FOXP3 является целевым геном miR-185-3p
Потенциальные сайты связывания фактора транскрипции промоторной области CCAT1 были предсказаны и проанализированы на сайте https://jaspar.genereg.net/, и он показал, что SOX2 и CCAT1 имеют потенциальные сайты связывания в промоторной области CCAT1 (рис. 6a). ChIP-qPCR сообщил, что (рис. 6b):в отличие от группы IgG, больше фрагментов промотора CCAT1 было обогащено группой SOX2 в сайте связывания E1, что доказывает, что SOX2 был связан с промотором CCAT1 в сайте E1, а SOX2 был связан с промотором CCAT1. участвует в регулировании CCAT1. Анализ двойного люциферазного репортерного гена показал, что (рис. 6c):активность люциферазы подавлялась в клетках, котрансфицированных sh-SOX2 и E1-WT, что указывает на то, что SOX2 может связываться с CCAT1.
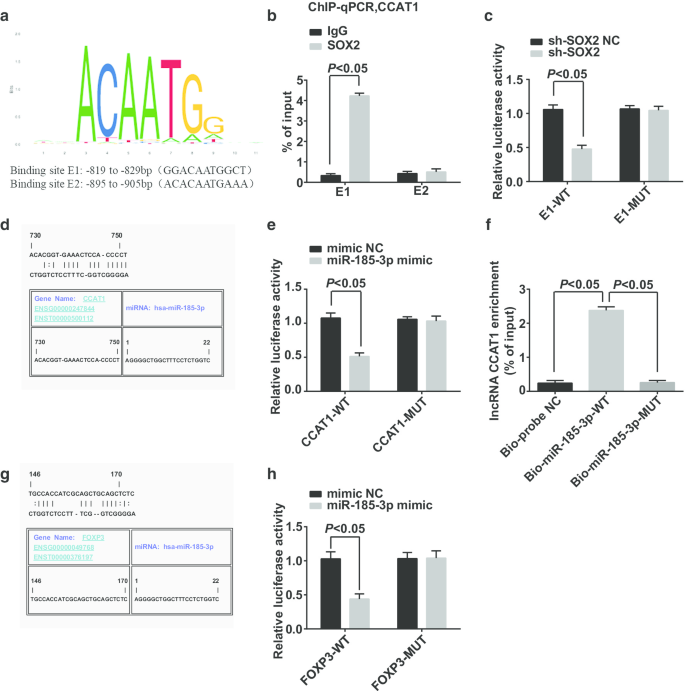
SOX2 может связываться с CCAT1, что влияет на экспрессию miR-185-3p, а FOXP3 является геном-мишенью для miR-185-3p. а Прогнозирование сайтов связывания в промоторных областях SOX2 и CCAT1 по сайтам биоинформатики. б ChIP-qPCR эксперимент подтвердил взаимосвязь связывания между SOX2 и CCAT1. c Сайт связывания SOX2 и CCAT1 подтверждается двойным анализом репортерного гена люциферазы. г Предсказание сайтов связывания в CCAT1 и miR-185-3p по сайтам биоинформатики. е Проверка связывания CCAT1 и miR-185-3p с помощью двойного анализа репортерного гена люциферазы. е Связь связывания между CCAT1 и miR-185-3p в клетках подтверждается анализом РНК с понижением. г Прогнозирование целевого отношения между miR-185-3p и FOXP3 на веб-сайте биоинформатики. ч Идентификация целевых отношений между miR-185-3p и FOXP3 с помощью двойного анализа репортерного гена люциферазы. Данные измерений были представлены как среднее значение ± стандартное отклонение, а сравнения между двумя группами оценивались с помощью t-критерия
Сайт Jefferson предсказал, что CCAT1 может связываться с miR-185-3p (рис. 6d). Анализ двойного люциферазного репортерного гена сообщил, что (фиг. 6e) активность люциферазы была снижена в клетках, введенных с миметиком miR-185-3p и CCAT1-WT, предполагая, что miR-185-3p может связываться с CCAT1. Для проверки того, может ли CCAT1 связываться с miR-185-3p, использовали анализ «вытягивания вниз». Результаты показали, что (фиг. 6f) уровень обогащения CCAT1 в клетках, обработанных Bio-miR-185-3p-WT, заметно увеличился, в то время как уровень обогащения CCAT1 в клетках, обработанных Bio-miR-185-3p-MUT, показал нет существенной разницы. Этот результат продемонстрировал, что CCAT1 может адсорбировать miR-185-3p, тем самым влияя на экспрессию miR-185-3p.
Целевые отношения между miR-185-3p и FOXP3 были предсказаны веб-сайтом Jefferson (рис. 6g). Анализ двойного люциферазного репортерного гена подтвердил, что (рис. 6h) относительная активность люциферазы клеток резко снижается после того, как миметик FOXP3-WT и miR-185-3p котрансфицирован в CD44 + Клетки HeLa, в то время как FOXP3-MUT, котрансфицированные миметиком miR-185-3p, не влияли на относительную люциферазную активность клеток, что позволяет предположить, что miR-185-3p нацелена на FOXP3.
Обсуждение
РС является четвертым по частоте злокачественным новообразованием у женщин в мире, за ним следуют рак груди, толстой кишки и легких [3]. Сообщается, что клетки CC с положительной экспрессией SOX2 проявляют характеристики раковых стволовых клеток [21]. В исследовании сообщается, что CCAT1 является важной онкогенной днРНК, связанной с CC, и играет вспомогательную роль в росте и инвазии CC клеток [11]. Другое исследование показывает, что miR-185-3p может предсказывать радиочувствительность карциномы носоглотки и регулировать рост и апоптоз раковых клеток [22]. Сообщается, что циркулирующие аутоантитела к FOXP3 отражают непрерывное развитие поражения шейки матки и могут быть потенциальным биомаркером для раннего прогноза CC [23]. Настоящее исследование было разработано для изучения того, как ось SOX2 / CCAT1 / miR-185-3p / FOXP3 влияет на пролиферацию и способность к самообновлению стволовых клеток CC.
Based on our findings, SOX2 and CCAT1 expressions were increased in CC tissues and cells that were linked to tumor size, LNM and advanced FIGO. Functionally proved, down-regulating SOX2 and CCAT1 declined proliferation, migration, invasion and sphere cells number and increased apoptosis of CC stem cells. Similar to our study, SOX2 expression trends to increase in CC [24, 25]. Moreover, SOX2 expression is also up-regulated in CC cells derived from cancer stem cells [26]. Overexpressed SOX2 was suggested to link with clinicopathological characteristics of patients with several types of cancer, not limited to CC. For example, it was suggested that up-regulated SOX2 shows in cervical squamous cell carcinoma patients staged in FIGO I-II [27]. Moreover, SOX2 expression is linked to LNM in oral squamous cell carcinoma [28]. When it comes to the molecular function of SOX2 for cancer progression, there is an observational work presenting that down-regulated SOX-2 suppresses cell migration and invasion of cervical squamous cell carcinoma [29]. Meanwhile, another research has offered a proof that up-regulated SOX2 enhances CC cell clonogenicity, proliferation and tumorigenicity in vitro and in vivo than control cells [30].
Concerning to the regulatory relation between SOX2 and CCAT1, an existed study has presented that silencing SOX2 markedly reduces CCAT1 mRNA level [31]. As to the role of CCAT1 in cancers, a study has showed that CCAT1 expression is markedly elevated in CC tissues versus in the adjacent normal tissues [11, 12]. Of note, CCAT1 overexpression in CC is positively related to the tumor size [12]. In terms of the role of CCAT1 in cancer cell activity, there is a research highlighting that overexpressed CCAT1 accelerates CC cell proliferation, colony formation and invasion [11]. Interestingly, a previous research has demonstrated that the cell viability, invasive and migratory abilities are declined via knocking down CCAT1 [12]. Anyway, the functional effect of SOX2 and CCAT1 in other cancers was similar to that in CC.
Afterward, our research revealed that CCAT1 could bind to miR-185-3p, the down-regulated CCAT1 in CC and overexpressing miR-185-3p suppressed the proliferation and self-renewal abilities of CC stem cells. It is reported that CCAT1 and miR-185-3p are negatively correlated [13]. Furthermore, a result reported that a reduction is seen in miR-185-3p expression in radioresistant nasopharyngeal carcinoma cases [22]. Regarding to the suppressive function of miR-185-3p in cancer cell aggressiveness, a study has revealed that up-regulation of miR-185-3p suppresses the invasive and metastatic properties of nasopharyngeal carcinoma cells [32]. Furthermore, Zou et al. have suggested that restored miR-185 represses breast cancer cell growth and invasion [33]. There is a article finding that up-regulation of miR-185 declines the proliferation, invasion and colony formation capacities of non-small cell lung cancer cells in vitro [34]. It is presented that in vitro cell proliferation, invasion and migration as well as in vivo tumor growth are suppressed via miR-185-overexpressing in non-small cell lung cancer cells [35]. From those studies, the anti-tumor role of miR-185-3p in the present study was consistent with previous researches.
To proceed, we unveiled that miR-185-3p targeted FOXP3, the overexpressed gene in CC to regulate CC stem cell activities. In fact, FOXP3, the regulator of SOX2 cancer stem-like cell marker in colon cancer [36], has been investigated in CC, showing an up-regulation in CC cells [19] [37]. It was evidenced that elevating FOXP3 promotes the formation of tumor spheres and stimulates the stemness of non-small cell lung cancer cells [38].
Conclusion
Collectively, we explored for the first time that SOX2 transcription could activate CCAT1, thereby inhibiting miR-185-3p and regulating FOXP3 to promote the proliferation and self-renewal of CC stem cells, which is a potential avenue to treat CC. Additionally, however, limitations in this present study still exist in the relatively small trial size in the designed experiment. Thus, clinical researches might be further carried out to detect the efficacy for the treatment of CC.
Доступность данных и материалов
Не применимо.
Наноматериалы
- Демонстрация гибкого биосенсора на основе графена для чувствительного и быстрого обнаружения клеток рака яи…
- Наночастицы для терапии рака:текущий прогресс и проблемы
- Доставка лекарств на основе клеток для онкологических приложений
- Нанотехнологии инактивируют раковые стволовые клетки
- Влияние распределения наночастиц золота в TiO2 на оптические и электрические характеристики сенсибилизирован…
- Каркасы nHAC / PLGA, гибридизированные с оксидом графена, способствуют пролиферации клеток MC3T3-E1
- Исследования TEM и STEM морфологии поперечного сечения двух- / трехслойных широкополосных антиотражающих пленок…
- Изготовление и характеристика ZnO Nano-Clips с помощью процесса, опосредованного полиолом
- Частотно-модулированный волновой диэлектрофорез везикул и клеток:периодические развороты на частоте кроссо…
- Факторы, влияющие на успех и безопасность процесса штамповки