


Исследования по созданию и применению точного индикатора pH лизосом на основе саморазлагающихся наночастиц
Аннотация
Кислый pH лизосом тесно связан с аутофагией; таким образом, хорошо известные лизосомы, изменения pH дадут больше информации о процессе и статусе аутофагии. Однако до сих пор можно было указать только изменения pH в относительно широком диапазоне, точное определение pH лизосом так и не было достигнуто. В нашем исследовании мы установили индикатор pH эндо / лизосом на основе саморазлагающегося SiO 2 система наночастиц с определенными параметрами синтеза. Центральный концентрированный метиленовый синий (МБ) в центральных полых структурных наночастицах демонстрирует чувствительное высвобождение как функцию значений pH от pH 4,0–4,8, что в точности соответствует диапазону pH лизосом. Построена линейная корреляция значений оптической плотности (OD) и значений pH, которая была использована для определения pH лизосом в 6 различных линиях клеток. Более того, с помощью этой системы нам удалось точно определить изменения среднего pH лизосом до и после эндоцитоза NP черного мезопористого кремния (BPSi), прояснив механизм прекращения аутофагии после эндоцитоза BPSi. Таким образом, индикатор pH просвета на основе саморазлагающихся наночастиц может предоставить новую методологию и стратегию, позволяющую лучше узнать pH лизосомы, а затем указать более подробную информацию о процессе аутофагии или других важных сигналах о метаболизме.
Введение
Лизосомы служат конечным местом назначения для макромолекул, где эти макромолекулы расщепляются гидролитическими ферментами, активируемыми при низком pH [1]. Кислый pH лизосом, поддерживаемый H + -АТФазой вакуолярного типа (v-ATPase) [2], которая перекачивает протоны из цитоплазмы в просвет лизосом, должен был поддерживать активность ~ 60 типов гидролитических ферментов [3]. Более того, недавние литературные сообщения показали, что кислый pH лизосом тесно связан с аутофагией [4], так что хорошо известные точные изменения pH лизосом дадут больше информации о процессе и статусе аутофагии. Основываясь на наших исследованиях и обзоре литературы, эндоцитоз аминоположительных заряженных наночастиц, вероятно, увеличит изменение pH в эндо / лизосомах, таких как наночастицы, украшенные первичным и вторичным амином PEG, или некоторые гидрофильные украшения на поверхности частиц [5, 6].
Повышение pH, вызванное эндоцитозом наночастиц амина, резко увеличит ядерную локализацию транскрипционного фактора EB (TFEB) [7], что приводит не только к усилению транскрипции пути, но также вызывает дисфункцию лизосом, что в конечном итоге приводит к блокированию аутофагического потока [7]. 7,8,9]. Поскольку TFEB регулирует аутофагию и, следовательно, его сверхэкспрессия приводит к значительному увеличению продукции аутофагосом в культивируемых клетках.
Таким образом, для прогнозирования процесса аутофагии и деталей аутофагии очень важны точный pH лизосомы и измерение его изменения. До сих пор, исходя из значений pH эндо / лизосом, указывающих на обзоры литературы [10], и коммерческих продуктов для обнаружения значений pH эндо / лизосом, можно было указать только изменения pH в относительно широком диапазоне, а точное определение pH лизосом так и не было достигнуто. . Таким образом, для понимания деталей аутофагии важным подходом является создание точного метода определения изменения pH в просвете.
Основываясь на нашем предыдущем опыте с саморазлагаемым SiO 2 наночастиц, в этом исследовании мы установили точный индикатор pH, который может определять изменения pH в просвете. SiO 2 наночастицы обладают хорошими преимуществами в регулируемом размере и биосовместимости [11]. Установив определенные параметры синтеза, установленный саморазлагаемый SiO 2 Индикатор pH может чутко высвобождать полезный метиленовый синий (МБ) при pH 4,0–4,8, что в точности соответствует диапазону pH лизосом. Более того, высвобождение МБ имеет линейную корреляцию с изменением значения pH (схема 1). Затем мы проверили возможность использования индикатора pH на клеточных уровнях, введя 6 различных клеточных линий, удалось определить средние изменения pH лизосом до и после эндоцитоза НЧ черного мезопористого кремния (BPSi), прояснив механизм прекращения аутофагии после BPSi. эндоцитоз. Таким образом, индикатор pH просвета на основе саморазлагающихся наночастиц может предоставить новую методологию и стратегию, позволяющую лучше узнать pH лизосомы, а затем указать более подробную информацию о процессе аутофагии или других важных сигналах, касающихся метаболизма.
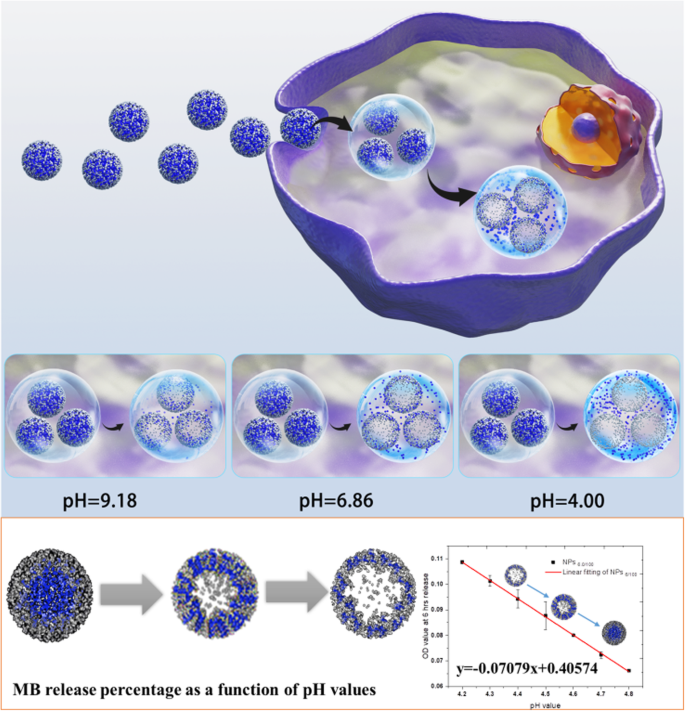
Схематическое изображение MB @ SiO 2 измерение pH лизосом в живых клетках
Материалы и методы
Раздел материалов
Силицид натрия (NaSi) и пластины Si (диаметр 20 см, p + (100), 0,01–0,02 Ом см) были предоставлены SiGNa Chemistry Inc. и Ocmetic Inc. соответственно. Бромид аммония (NH 4 Br, 99%), бромид натрия (NaBr, 99%), толуол (безводный, 99,8%), соляная кислота (HCl, 37%), МБ и тетраэтилортосиликат (TEOS) были закуплены у Sigma-Aldrich. Метокси-ПЭГ-силан 0,5 кДа и метокси-ПЭГ-силан 2 кДа были приобретены отдельно у Fluorochem Ltd. и Laysan Bio Inc. Среда RPMI 1640 была предоставлена Life Technologies. Фетальная бычья сыворотка (FBS) была куплена в компании TianHang Biological Technology. Бикарбонат натрия, сульфат стрептомицина, пенициллин G, HEPES, раствор лизоцима, CellLight Early-endosomes-GFP, LysoTracker ™ Red DND-99, набор для анализа белка Pierce® BCA, усиленная хемилюминесценция, конъюгат pHrodo ™ Red Transferrin, раствор для визуализации живых клеток и Реагент Trizol был куплен у Thermo Fisher Scientific. Этанол и аммиак-вода были предоставлены Sinopharm. Набор для синтеза первой цепи кДНК BioRT Master HiSensi был куплен у Hangzhou Bioer Technology Co., Ltd. Лизат RIPA был куплен у Heart Biological Technology Co., Ltd. P62, TFEB и антитело против β-актина были куплены у Proteintech Group , Inc. Антитело LC 3B было предоставлено Abcam. 2- (4-Пиридил) -5 - ((4- (2-диметиламиноэтиламинокарбамоил) метокси) фенил) оксазол (PDMPO) предоставлен Yeasen Biotech Co., Ltd.
Цель, дизайн и постановка исследования
Целью данного исследования является (1) изучить влияние наночастиц BPSi на аутофагию в клетках HepG2, (2) выяснить скрытый механизм изменений значения pH лизосом, влияющих на аутофагию, (3) установить точный индикатор pH лизосом, который может измерять точное значение pH лизосом и, наконец, (4) указывает на влияние колебаний pH на аутофагию. Чтобы реализовать вышеуказанную цель исследования, мы использовали эксперимент по секвенированию транскриптома для изучения изменений генов транскриптома в клетках HepG2 после подачи наночастиц BPSi и подтвердили это с помощью RT-qPCR и Western-экспериментов. Флуоресцентные красители, такие как PDMPO, использовали для измерения изменения лизосомного pH в клетках HepG2 после кормления BPSi. Чтобы точно измерить pH лизосом, мы разработали MB @ SiO 2 наночастиц с 10 параметрами и протестировали характеристики этих 10 видов наночастиц с помощью таких экспериментов, как DLS и HR-TEM. Эффективность загрузки MB и исследование кинетики высвобождения 10 серий саморазлагающихся систем наночастиц были протестированы в растворах с различным pH и клетках HepG2. Чтобы проверить внутриклеточное расположение наночастиц после попадания в клетку, мы провели эксперимент TEM на клетках и конфокальную микроскопию живых клеток. Наконец, мы измерили изменения лизосомного pH в 6 типах клеток после кормления BPSi, чтобы проверить универсальность MB @ SiO 2 наночастицы для измерения изменений pH лизосом.
Синтез наночастиц BPSi
Наночастицы BPSi были приготовлены нашим предыдущим методом [12] и предоставлены нашим сотрудником (Wujun Xu, факультет прикладной физики Университета Восточной Финляндии). Получение BPSi, NaSi, аммониевой соли и NaBr (NaSi:NH 4 Br:NaBr 1:4:4, мас. / Мас.) Измельчали в перчаточном ящике с атмосферой Ar. Им позволили прореагировать в трубчатой печи под N 2 атмосферу при 240 ° C в течение 5 часов (уравнение 1). После охлаждения до температуры окружающей среды полученные микрочастицы очищали промывкой отдельно 0,5 М HCl и 1,0 М растворами HF. Микрочастицы измельчали на шаровой мельнице в этаноле при 1000 об / мин в течение 15 минут и собирали наночастицы BPSi желаемого диаметра, регулируя скорость центрифугирования.
$$ \ mathrm {NaSi} + {\ mathrm {NH}} _ 4 \ mathrm {Br} \ to \ mathrm {NaBr} + {\ mathrm {NH}} _ 3+ \ mathrm {Si} / \ mathrm {H} + { \ mathrm {H}} _ 2 $$ (1)С помощью эксперимента динамического рассеяния света были изучены распределение диаметров и поверхностный заряд наночастиц. Все НЧ были диспергированы в среде после стерилизации небольшим ультразвуковым воздействием (5 с для их равномерного диспергирования в растворе, ультразвуковой очиститель SB-5200DT, Ningbo Scientz Biotechnology Co., Ltd.) непосредственно перед их введением в клетки.
Создание системы саморазлагаемых наночастиц серии 10
10 серий саморазлагающихся наночастиц были синтезированы по методикам, о которых мы сообщали ранее [13,14,15,16], с измененными параметрами. В типичной процедуре определенное количество МБ сначала добавлялось к смеси этанола (75 мл) с водным раствором аммиака (25%, 3,4 мл), после чего добавлялось определенное количество TEOS. Серия саморазлагаемых MB @ SiO 2 НЧ получали после перемешивания в течение 24 ч и промывали 3 раза перед сушкой. Количества МБ и TEOS, добавленные в протоколы, были такими, как описано в таблице 1. Значение 1,0 / 100 в NP 1,0 / 100 представляли инвентарь МБ и TEOS, когда мы синтезировали наночастицы, с 1,0 мг МБ и 100 мкл TEOS. И значения 1,5 / 100 в НП 1,5 / 100 и другие соответствуют 1.0 / 100.
Культура клеток
Чтобы проверить эффективность и универсальность индикатора pH, основанного на саморазлагающихся наночастицах, мы попытались протестировать его на конкретных линиях опухолевых клеток. Таким образом, в качестве объектов исследования мы выбрали линии клеток рака печени, легких, рака толстой кишки и меланоцитомы. Клеточные линии клеток рака толстой кишки человека HCT116, HCT8 и HCT15; клетки рака печени человека HepG-2; клетки рака легких человека A549; и клетки меланомы мыши B16 поддерживали в среде RPMI 1640 (Life Technologies) с добавлением 10% инактивированного нагреванием FBS, 2,0 г / л бикарбоната натрия, 0,1 г / л сульфата стрептомицина, 0,06 г / л пенициллина G и 5,958 г / л. ГЕПЕС. Клетки поддерживали в стандартном инкубаторе для клеточных культур при 37 ° C в увлажненной атмосфере с 5% CO 2 .
Характеристики саморазлагаемых систем наночастиц серии 10
Морфология наночастиц всех серий была охарактеризована с помощью HR-TEM с режимом STEM, а отображение Si было изучено с помощью сопоставления элементов EDS. Анализ распределения наночастиц по размерам был выполнен с помощью программного обеспечения ImageJ путем расчета диаметров наночастиц на случайно выбранных изображениях STEM. Дзета-потенциал наночастиц и индекс полидисперсности (PDI) были измерены с помощью исследования динамического светорассеяния (DLS) в серии буферов с определенными значениями pH. Данные были проанализированы с помощью SPSS15.0, и статистические результаты были представлены как среднее ± стандартное отклонение
Исследование эффективности загрузки МБ и кинетики высвобождения саморазлагаемых систем наночастиц серии 10
Для изучения эффективности загрузки МБ и кинетики высвобождения сначала была построена стандартная кривая для МБ в сериях концентраций. Поглощение МБ осуществляли по спектру УФ – видимой области с поглощением при 660 нм, что является λ max мономера MB. Эффективность загрузки МБ рассчитывалась по приведенному ниже уравнению:эффективность загрузки МБ (%) =количество инкапсулированных МБ / (общий объем введенных МБ).
Высвобождение МБ из наночастиц 10 серии изучали в чистой воде и буферах pH с разными значениями pH (pH 4,0, pH 6,86 и pH 9,18) и растворе лизоцима (Thermo Scientific ™ # 90082). Кроме того, также была исследована кинетика высвобождения МБ после определенного периода времени в буферах с различным pH. Затем были изучены значения OD при 660 нм и процент высвобождения МБ в зависимости от времени.
Более подробно исследования высвобождения МБ были выполнены по протоколам, приведенным ниже; растворил наночастицы серии 10 в 15 мл стандартного буфера с pH 4,0, 6,86 и 9,18 с раствором лизосом, соответственно; и выполнили высвобождение МБ в смесителе Hula при 37 ° C. В течение следующих 15 дней собирали 1 мл каждого образца, затем центрифугировали при 12000 об / мин в течение 10 минут. Спектры поглощения супернатанта и осадка измеряли при 200–800 нм.
Кроме того, высвобождение МБ в буферах с точным pH с раствором лизоцима в диапазоне pH от 4,1 до 5,5 также исследовали с использованием тех же протоколов, описанных выше. Определенные промежутки времени (6 ч, 12 ч и 24 ч) были помещены в качестве точек времени наблюдения. Поглощение при 660 нм регистрировали в каждом образце. Линейная зависимость поглощения каждого раствора pH и сумма квадратов остатков подсчитывались в каждый момент времени соответственно.
Чтобы определить профили высвобождения МБ в клетках, клетки HepG-2 культивировали в 75 см 2 в колбу для культивирования и в нее вводили НЧ (300 мкг / мл), когда клетки размножались до 70% культуральной колбы. Каждые 30 минут клетки собирали. Клетки многократно замораживали и оттаивали для полного высвобождения МБ в клетках. Лизаты клеток центрифугировали при 12000 об / мин в течение 10 мин. Получали супернатант и измеряли его оптическую плотность при 660 нм для расчета общего количества высвободившегося МБ. В этом исследовании в качестве объектов исследования были выбраны клетки HepG2 из-за их быстрой клеточной пролиферации, которая могла минимизировать различия между всеми 10 тестируемыми группами.
Совместная клеточная локализация наночастиц серии 10 и эффективность высвобождения в 6 различных клеточных линиях
Клеточный TEM использовался для изучения совместной локализации наночастиц в эндо / лизосомах в соответствии со стандартными протоколами клеточного TEM. Клетки высевали с интенсивностью 1 × 10 6 клеток / колбу и инкубируют в течение 24 ч, позволяя клеткам прикрепиться. Десять серий наночастиц в среде с той же концентрацией (100 мкг / мл) инкубировали с клетками еще 12 и 24 ч соответственно. Затем клетки промывали 3 раза PBS для удаления избыточных наночастиц, затем фиксировали в 2,5% растворе глутарового альдегида более 1 дня. Затем фиксированные клетки промывали и окрашивали тетроксидом осмия, 1% в деионизированной воде в течение 1 ч, с последующей промывкой 3 раза PBS и 2 раза деионизированной водой. Затем был проведен классический протокол TEM клеток [17, 18], и срезы толщиной 90 нм были собраны для наблюдения TEM. Высвобождение МБ в зависимости от значений pH изучали на 6 линиях клеток с обоими НЧ 6/100 и НП 7.5 / 80 . Кроме того, для анализа данных записывались значения OD от выпуска MB и процент выпуска MB.
Исследование внутриклеточного поглощения MB @ SiO 2 Наночастицы
Конфокальная микроскопия живых клеток была использована для оценки клеточного поглощения и внутриклеточной судьбы MB @ SiO 2 наночастицы. Ранние эндосомы клеток HepG-2 окрашивали (CellLight Early-endosomes-GFP, BacMam 2.0 ThermoFisher Scientific C10586, с возбуждением / испусканием ~ 488/510 нм) в течение 16 часов. А затем клетки инкубировали с НЧ 6/100 (Возбуждение / испускание МБ:640 / 650-700 нм) при концентрации наночастиц 100 мкг / мл через определенные интервалы времени (2 ч, 2,5 ч, 3 ч, 5 ч и 6 ч). Перед получением изображений лизотрекер окрашивали LysoTracker ™ Red DND-99 (Thermo Fisher Scientific L7528, возбуждение / испускание:577/590 нм) в течение 40 минут. После этого удалите раствор красителя и 2–3 раза промойте клетки в PBS. Изображения были получены с помощью конфокального микроскопа Nikon A1R.
Секвенирование транскриптома для оценки изменения экспрессии гена после кормления BPSi
Экстракцию тотальной РНК контрольной группы и группы, обработанной BPSi, проводили с использованием реагента Trizol в соответствии со стандартными рабочими процедурами. Качество исходного образца тотальной РНК для эксперимента по секвенированию определяли с помощью спектрофотометра NanoDrop ND-2000. Тотальную РНК, прошедшую контроль качества, использовали в последующих экспериментах по секвенированию. Сравнение экспрессии генов было выполнено секвенированием следующего поколения. Все программы секвенирования были выполнены BGI-Shenzhen Corporation (Шэньчжэнь, Китай) с использованием платформы BGISEQ-500. Необработанные данные, полученные путем секвенирования, используются для контроля качества, чтобы определить, подходят ли данные секвенирования для последующего анализа. В случае успешного прохождения проведите количественный анализ генов на основе уровней экспрессии генов и проведите значительный обогащающий анализ функций онтологии генов (GO) для дифференциально экспрессируемых генов в отобранных выборках.
Анализ количественной полимеразной цепной реакции с обратной транскрипцией (RT-qPCR) для подтверждения активации генной сети TFEB-CLEAR
Суммарную РНК экстрагировали из культивированных клеток HepG-2 контрольной группы и группы, обработанной BPSi, с использованием реагента Trizol и подвергали обратной транскрипции в кДНК с помощью набора для синтеза первой цепи кДНК BioRT Master HiSensi (Hangzhou Bioer Technology Co., Ltd. .) со случайными праймерами. кДНК использовали для амплификации генной сети TFEB-CLEAR с помощью количественной ПЦР с системой ПЦР в реальном времени Applied Biosystems ™ 7500 (Applied Biosystems, Life Technologies, Карлсбад, Калифорния) с актином в качестве эталонного контроля. Праймеры, использованные для количественной ОТ-ПЦР, перечислены в таблице S4.
Вестерн-блоттинг для подтверждения активации аутофагии после кормления BPSi
Клеточные белки контрольной группы и групп, обработанных BPSi с различной концентрацией, экстрагировали лизатом RIPA (Heart Biological Technology Co., Ltd.). Ингибитор протеазы добавляли к лизату RIPA и предварительно охлаждали на льду. Промыли клетки 3 раза предварительно охлажденным PBS. Полностью слили жидкость и поместили посуду во лед на 2 мин. Четыреста микролитров лизата RIPA добавляли на поверхность всей чашки, несколько раз пипетировали пипеткой и инкубировали на льду в течение 30 минут, в течение которых чашку встряхивали несколько раз для полного лизиса клеток. Жидкость лизированных клеток переносили в пробирки Эппендорфа на 1 мл и центрифугировали при 13000 об / мин в течение 10 мин при 4 ° C. Полученный супернатант кипятили в воде в течение 10 мин и помещали в - 20 ° C для дальнейшего использования. Концентрацию белка определяли количественно с помощью набора Pierce® BCA Protein Assay Kit (Thermo Scientific).
Экстракты клеток, содержащие 25 мкг общего белка, непосредственно подвергали SDS-PAGE и переносили. Мембраны были заблокированы 5% обезжиренным молоком и исследованы первичными антителами, распознающими P62 (Proteintech # 18420–1-AP), TFEB (Proteintech # 13372–1-AP), LC 3B (Abcam # ab192890) и β-актин (Proteintech # 20536–1-АП). Вторичные антитела были выбраны в соответствии с видом происхождения первичных антител и обнаружены с помощью усиленной хемилюминесценции (Pierce) или с помощью системы визуализации гелей Bio-Rad ChemiDoc XRS + (Bio-Rad, США). Нормализованная интенсивность полосы P62, TFEB и LC 3B относительно β-актина была количественно определена денситометрией с использованием программного обеспечения ImageJ в группах BPSi, и данные представляют собой среднее значение ± стандартное отклонение. из трех независимых экспериментов.
Измерение pH клеток с помощью PDMPO и конъюгата pHrodo ™ Red с помощью конфокальной микроскопии
Исследование PDMPO
1 × 10 5 Клетки HepG-2 культивировали на стерильных конфокальных планшетах в течение ночи, а наночастицы BPSi подавали с концентрацией 100 мкг / мл. На следующий день перед иммунофлуоресцентным окрашиванием слайды трижды промывали 0,01 М фосфатно-солевым буфером (PBS), pH 7,4, затем добавляли 1 мкМ красителя PDMPO (Ex / Em =329/440). После трехкратной промывки PBS клетки инкубировали со свежей культуральной средой RPMI-1640 и наблюдали под флуоресцентным микроскопом (Nikon A1R, Япония) с камерой CCD и делали снимки в течение 5 минут, а соотношение интенсивности синей и зеленой флуоресценции в лизосомах затем рассчитывали согласно процедуре Chen et al. [19].
Исследование конъюгата pHrodo ™ с красным трансферрином
Клетки HepG-2 помещали в конфокальные планшеты таким же образом для прикрепления клеток в течение 24 ч, затем держали планшеты на льду в течение 10 мин. Промытые клетки холодным раствором для визуализации живых клеток, содержащим 20 мМ глюкозы и 1% BSA. Добавляли конъюгат pHrodo ™ Red Transferrin (Ex / Em =560/585 нм) в концентрации 25 мкг / мл в раствор для визуализации живых клеток и инкубировали при 37 ° C в течение 20 мин, затем промывали клетки в растворе для визуализации живых клеток. Наблюдение также проводилось с помощью конфокальной микроскопии. Количественный анализ интенсивности микроскопических изображений был выполнен с помощью программного обеспечения ImageJ.
Определение pH лизосом клеток
Клетки A549, HepG-2, HCT8, HCT15, HCT116 и B16 культивировали в 75 см 2 во флакон для культивирования и в него вводили НЧ, когда клетки размножались до 70% культурального флакона, а через 6 часов клетки собирали. Клетки многократно замораживали и оттаивали для полного высвобождения МБ в клетках. Лизаты клеток центрифугировали при 12000 об / мин в течение 10 мин. Получали супернатант и измеряли его оптическую плотность при 660 нм для расчета общего количества высвободившегося МБ. Поглощение НЧ 6.0 / 100 при стандартном pH сравнивали, чтобы получить значение pH каждой ячейки.
Статистический анализ
Статистический анализ был выполнен с помощью программного обеспечения SPSS15.0 с использованием двустороннего дисперсионного анализа (ANOVA) для независимых групп и с использованием метода Tukey HSD для теста множественных сравнений. Статистическая значимость основана на значении P . <0,05.
Результаты и обсуждение
Мы впервые обнаружили дифференциальную экспрессию генов, когда клетки питались двойными функциональными наночастицами черного пористого кремнезема (BPSi NP), которые были предложены нашей лабораторией [12]. Изменение дзета-потенциала с -18,5 до +2,8 мВ также указывало на то, что поверхностное двойное ПЭГилирование было успешным (рис. S1a). Средний диаметр наночастиц BPSi составлял 156 нм (рис. S1b). На основе кластерной тепловой карты дифференциальной экспрессии генов (рис. S2) мы выбрали более чем 2-кратную дифференциальную экспрессию генов для дальнейшего исследования. Го и KEGG были введены для анализа дифференциальных генов. Из пузырьковых карт обогащения Go (рис. 1а) для дальнейшего анализа были выбраны метаболические и связанные с лизосомами гены, включая сборку фаголизосом, фагоцитоз и метаболический процесс ксенобиотиков. Примечательно, что экспрессия генов, ассоциированных с TFEB-CLEAR [17], была значительно увеличена. Результаты ОТ-ПЦР (рис. 1b) также подтвердили результаты секвенирования генов, гены на TFEB - скоординированной лизосомной экспрессии и пути регуляции (CLEAR) значительно увеличились, такие как CTSD, CTSF, TFEB, MFN1, LAMP2 и TPP1. . Эти гены были помечены на пути лизосом, как показано на рис. S3. Их экспрессия выше, чем в контрольной группе, и имеет статистическую значимость ( P BPSi VS Control <0,05). А TFEB положительно регулирует экспрессию лизосомных генов, контролирует популяцию лизосом и способствует клеточной деградации лизосомных субстратов.
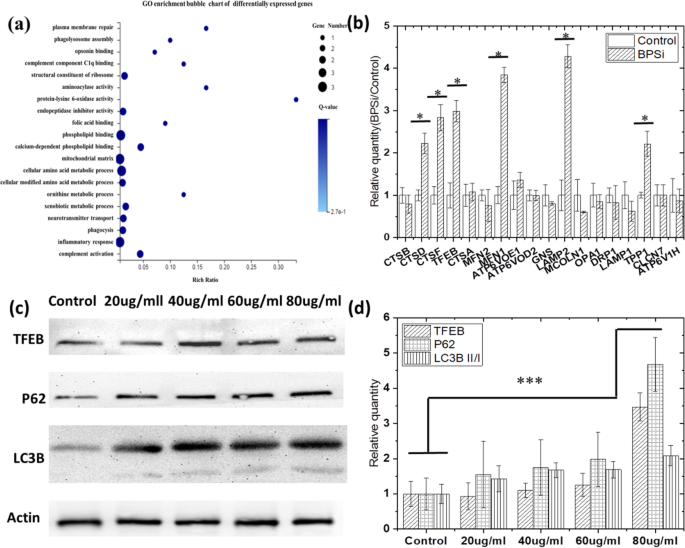
Аутофагия активируется в клетках HepG-2 после кормления BPSi. а Обогащенная пузырьковая карта GO дифференциально экспрессируемых генов, обнаруженных с помощью секвенирования транскриптома. б Проверка изменений генов в генной сети TFEB-CLEAR после обработки BPSi с помощью экспериментов RT-qPCT. c Экспрессия белков P62, TFEB, LC3B II / I после обработки BPSi. г Нормализованная интенсивность серого цвета полосы белка P62, TFEB, LC3B II / I в группах, получавших BPSi, в соответствии с контрольной группой. Данные представлены как среднее ± стандартное отклонение
Кроме того, TFEB регулирует аутофагию, и его сверхэкспрессия приводит к значительному увеличению продукции аутофагосом в культивируемых клетках из-за того, что основная функция гена TFEB состоит в том, чтобы индуцировать биосинтез лизосом и способствовать возникновению аутофагии [20]. Вестерн-блоттинг также использовался, чтобы подтвердить, происходила ли аутофагия, когда клетки питались НЧ BPSi. Целью Вестерн-теста было дополнительное подтверждение того, что экспрессия TFEB была увеличена и возникновение клеточной аутофагии после кормления BPSi. Белки LC3B и P62 являются маркерами аутофагии. Когда происходит аутофагия, экспрессия ассоциированного с микротрубочками белка 1A / 1B-легкой цепи 3B (LC3B) II / I увеличивается. p62 является рецептором для везикул, которые будут разлагаться аутофагией, а также рецептором для убиквитинированных белковых агрегатов, которые должны быть очищены, и его экспрессия снижается при возникновении аутофагии. Поэтому мы измеряем экспрессию этих белков в западном эксперименте.
Из результатов вестерн-блоттинга, показанных на фиг. 1c, d, TFEB ( P 80 мкг / мл VS контроль =0,000008), LC3B II / I ( P 80 мкг / мл VS контроль =0,000297) и p62 ( P 80 мкг / мл VS контроль =0,000016) все белки значительно активированы. Поскольку повышающая регуляция белков TFEB и LC3B II / I указывает на активацию аутофагии [18], мы подозревали, что эндоцитоз BPSi способствует возникновению аутофагии. Однако предполагается, что белок p62 подвергается подавлению во время процесса аутофагии из-за природы белка-носителя, который переносит эндосомы в лизосомы и, наконец, разрушается. В нашем исследовании значительная повышающая регуляция p62 указывает на прекращение деградации во время процесса слияния эндолизосом [21], что, вероятно, вызвано повышением pH в везикулах эндолизосом. Таким образом, эндоцитоз BPSi может сначала вызвать возникновение аутофагии, а затем ингибировать процесс аутофагии за счет увеличения значений pH эндо / лизосомы из-за его амидной щелочности.
Чтобы подтвердить характеристики повышения pH в эндо / лизосомах за счет эндоцитоза BPSi, в нашем исследовании были использованы два коммерческих флуоресцентных зонда pHrodo ™ Red Transferrin Conjugate (Thermo Fisher # P35376) и RatioWorks ™ PDMPO.
pHrodo ™ Red в качестве коммерческого индикатора внутриклеточного pH обычно имеет слабую флуоресценцию при нейтральном pH, но усиливается при падении pH. Предполагалось, что для количественной оценки клеточного цитозольного pH в диапазоне 9–4 с pKa ~ 6.5 с возбуждением / испусканием 560/585 нм. На основании 6 определений клеточных линий мы смогли получить качественный вывод анализа о том, что эндоцитоз НЧ BPSi обладает способностью увеличивать значения pH в эндо / лизосомах из-за ослабления красных флуоресцентных сигналов (рис. S4 и S5). Однако после повторения экспериментов несколько раз в соответствии с протоколами работы с продуктом мы с трудом количественно проанализировали точное значение pH, которое снизилось среди различных клеточных линий до или после кормления НЧ BPSi, из-за отсутствия корреляции между интенсивностью и установленными значениями pH. P>
PDMPO затем использовался как лучший раствор для индикации изменений значения pH после эндоцитоза BPSi, что позволяет использовать методы визуализации соотношения при количественном измерении pH. PDMPO [2- (4-пиридил) -5 - ((4- (2-диметиламиноэтиламинокарбамоил) метокси) фенил) оксазол] характеризуется как ацидотропный зонд с двойным возбуждением и двойной эмиссией. Он излучает интенсивную зеленую флуоресценцию при более низком pH и дает интенсивную синюю флуоресценцию при более высоком pH. Эта уникальная pH-зависимая флуоресценция делает PDMPO идеальным датчиком pH для кислых органелл с pKa =4,47. PDMPO избирательно маркирует кислые органеллы (например, лизосомы) живых клеток, и два различных пика эмиссии можно использовать для мониторинга колебаний pH живых клеток при измерениях соотношения. Однако нам все еще не удалось измерить значения pH в 6 клеточных линиях до и после кормления BPSi. Как показывают результаты на фиг. S6, не наблюдали значительных различий во всех 6 клеточных линиях до и после кормления BPSi. Хотя была установлена корреляция между отношением синий / зеленый и значениями pH (рис. S7), нелинейная корреляция от pH 4–5 делает метод PDMPO неэффективным при количественном анализе эндо / лизосом до и после кормления BPSi.
Из данных двух коммерческих индикаторов pH, приведенных выше, мы сначала продемонстрировали наше подозрение, что декорированные ПЭГ наночастицы с амидом на цепи ПЭГ могут увеличивать рН эндо / лизосомы из-за щелочной природы амида. Однако без количественного анализа точных изменений pH (диапазон pH 0,1) мы по-прежнему не можем установить корреляции между статусом аутофагии и значениями pH эндо / лизосом, что не позволяет прогнозировать аутофагию.
Основываясь на нашем предыдущем исследовании саморазлагающихся наночастиц [13, 14] [15, 22], мы сохранили ту же концентрацию гидроксида аммония в 75% этаноле, но скорректировали концентрации MB и TEOS. Две серии количества TEOS были установлены как 100 мкл и 80 мкл, чтобы получить разную толщину оболочки и размер пор. Было установлено десять серий количества МБ для получения различных размеров полой структуры и эффективности загрузки МБ.
Количества МБ и TEOS, добавленные в протоколы, были такими, как описано в таблице 1 ниже.
Как показано на рис. 2a, b, размер наночастиц увеличивался с увеличением количества МБ в обеих концентрациях TEOS (100 мкл и 80 мкл). При той же концентрации МБ размер частиц увеличивался с увеличением количества ТЭОС. Moreover, with the increase of the TEOS amount, the shell thickness grew up, which has been proved by the element mapping (shown as Fig. 2c). The polydispersity index (PDI) and surface charge of the nanoparticles are shown in Fig. S8 and Table S1. The morphology studies predicted that with the increase of MB amount, the loading efficiency will grow up, leading to the faster release profile, while with the increase of TEOS amount, the release will slow down. And we need to find out the appropriate MB and TEOS concentration, with which we could obtain the optimized nanoparticle systems, that we may be able to make the MB release profile linear correlated with the pH changes.

Morphology characterization of 10 different self-decomposable nanoparticles with specific MB or TEOS amount. а STEM figures. б Nanoparticle size distribution analysis. c Si mapping of 10 self-decomposable nanoparticles. Scale bars in all figures are 100 nm. The size distribution analysis was performed by randomly chosen 100 nanoparticles from STEM figures and measured by ImageJ software. Data was presented as mean ± S.D.
The MB loading efficiency was determined by UV-Vis spectrum. The standard curve (Fig. S9) of MB was firstly drawn using series concentrations of MB solution (from 6.25 to 46.88 μg/mL), with the equation as y = 67.63x + 0.10919, R 2 = 0.9987. As calculated with the equation above, we obtain MB loading efficiency of 10 self-decomposable nanoparticles with specific parameters, detailed data shown in Fig. S10.
Before the study of the MB release profiles in different pH solutions, the release profiles in pure water have been studied. As shown in Fig. S11 and Fig. S12, all the nanoparticles with TEOS amount of 80 μL presented increased MB release along with the duration increase, which was reflected by the UV–Vis absorption. Moreover, with the MB encapsulated amount increase, the growth trend of MB release becomes more significant. Also, the release velocity grows faster. However, as the TEOS amount increase to 100 μL, the particle surface became more densed and the release becomes slower when the MB amount below 3.0 mg; almost no increase trend could be observed in the MB release in water during 14 days of release. As long as the MB amount increases to above 4.0 mg, an obvious increase trend of MB release could be observed. One thing to be noticed is that the nanoparticle parameter of both NPs7.5/80 and NPs6.0/100 presented solid growth as the time prolongs, almost showed a linear increase trend during the first 7 days, and then reached the platform.
Then, we focused on the MB release behavior in different pH buffers to figure out whether self-decomposable nanoparticles with specific parameters could have the linear pH-dependent MB release.
Firstly, we carried out the MB release experiments at pH 4.0 buffer solution. From Fig. S13, we could easily reach the conclusion that with the same TEOS amount of 100 μL, the MB release velocity presented a similar trend was observed in the 5 nanoparticle systems of TEOS at 80 μL (Fig. S14), the center positive correlation with the MB encapsulated amount.
Concentrated MB diffuses into the surrounding solution via diffusion due to concentration difference. The bigger concentration gradient makes the faster MB release. Compared with the MB release in pure water, we found that the acidic environment speeded up the release of MB (Fig. S13 and S14 compared with Fig. S11 and S12), indicating that the MB release is not only driven by diffusion; however, in acidic solutions, electrostatic repulsion is also an important driven force due to the positive charge nature of MB. We then calculated the release percentage of each nanoparticle parameter according to the MB loading efficiency, MB standard curve, and the dilution ratio at measurements. The release percentage reflected the release speed of MB in pH 4.0 acidic solution, and the results (Fig. 3) showed that only the release percentage of NPs7.5/80 presented linear release in pH 4.0 solution. Other nanoparticle systems with specific MB and TEOS parameters showed similar release trends, and the release percentage did not have a linear growth. One exception is NPs6/100 , and the MB release reached the platform in only 72 h; thus, it was hard to tell whether the MB release could grow linear before that duration at this stage.

MB release percentage of 10 series self-decomposable nanoparticles after a specific duration in pH 4.0 buffer. All experiments were triple repeated, and the data were shown as mean ± S.D.
Meanwhile, we tested the MB release profiles in near-neutral and alkali buffers (pH 6.86 and pH 9.18). The results in both Fig. S15, S16, Fig. S17, and S18 demonstrated that the MB release slowed down with the solution pH increase to 6.86; moreover, with the central MB concentration increased, the MB release percentage decreased. At pH 9.18, all nanoparticles with 10 specific parameters presented a very slow MB release (Fig. S19 and S20); no matter in UV–Vis absorption or the release percentage, the trend was similar with the one in pH 6.86 buffer, but with even lower release percentage. So, it was clear that the self-decomposable nanoparticles only presented MB release linear growth in acidic solutions. We thought back to the endo/lysosomes pH, from 4 to 5, which is exactly the pH range of MB linear growth as a function of time in a specific MB/TEOS parameter. Thus, we get more confident that the self-decomposable nanoparticle system may be an accurate measuring tool for quantitative determining the endo/lysosome average pH, then provide evidence on the exact pH value of autophagy status.
The precondition of using the specific self-decomposable nanoparticles as an endo/lysosome pH indicator is that the nanoparticles stay stable in the endo/lysosome during the whole measurement process. Secondly, the MB release in endo/lysosome should occur smoothly when the measurement carried out.
The colocalization of the nanoparticle in the endo/lysosomes by cell TEM study and the MB release in 6 different cell lines were studied. From the cell TEM results, all of the 10 series nanoparticles stayed in the endo/lysosomes without escaping, after 24-h incubation with the HepG-2 cells (Fig. 4). Since the diameter of HepG2 cells used in Fig. 4 is about 10–20 μm and the diameter of MB@SiO2 nanoparticles is between 75 and 200 nm, it will be very difficult to clarify the nanoparticle morphologies using the images with low magnification (as shown in Fig. S21). We also investigated the intracellular location of the nanoparticles in the other 5 cell lines, and 4 nanoparticles were randomly selected to demonstrate the nanoparticles were trapped in the endo/lysosomes (Fig. S22). The nanoparticles with all parameters showed a central hollow structure in all other 5 cell lines after 24-h incubation, indicating the MB release. Moreover, under more precise observation, we noticed the MB release may be different due to different hollow sizes, pointing to the fact that (1) the endo/lysosome pH in different cells is different and (2) the MB release from the nanoparticles is very sensitive to the endo/lysosomes pH, especially for the NPs6/100 and NPs7.5/80 . From Fig. S23, we can find that nanoparticles have already realized the endocytosis and stayed in the vesicles 2 h after nanoparticle feeding, then nanoparticles gradually accumulated in lysosomes.

The colocalization of the nanoparticle in the endo/lysosomes by cell TEM study after 12 and 24 h incubation with the 10 series nanoparticles. The scale bar is 200 nm in the TEM images
We then evaluated the correlation between the pH values and the OD values in pH 4.0–4.8. From the results in Fig. 5 and Fig. S24, for NPs6/100 and NPs7.5/80 nanoparticle systems, the MB release presented a linear decrease as a function of pH in the pH range from 4.0 to 4.8.

MB release as a function of pH values in NPs6/100 and NPs7.5/80 after specific incubation duration, 6 h, 12 h, and 24 h. The linear correlation equations were also calculated for 6 h and 12 h for MB release from both NPs6/100 and NPs 7.5/80 as a function of pH values. All experiments were carried out triplicated, and the data were shown as mean ± S.D.
We then converted the OD value to the MB release percentage according to the MB loading efficiency and feeding amount. As shown in Fig. S25, in the first 6 h, the MB release percentage in NPs6/100 and NPs7.5/80 nanoparticle systems also presented as a function of pH values. We then calculated the residual sum of squares and Pearson’s related coefficient at 6 h and 12 h release duration, respectively, as the residual sum of squares present a negative correlation with closeness of linear fitting, while the closer the absolute value of Pearson’s related coefficient to 1, the more linear it is. As shown in Table S2 and S3, the highest degree of linearity is the fitting of NPs6.0/100 nanoparticle systems, followed by the one of NPs7.5/80 at 6 h release.
Till then, we were so excited by the results that the method for precisely monitoring the pH values has been established, especially with the accuracy less than or equal to 0.1 pH value interval. That means, we have great possibilities to figure out the correlation between endo/lysosome pH values and the autophagy status, which is of great significance for better studying the autophagy mechanism and predicting the autophagy process. As we can see in Fig. S26, the MB release in HepG-2 cells have already reached the plateau after incubation for 4 h. Thus, we chose 6 h as the observation time point.
We then carefully investigated the MB release of NPs6.0/100 in 6 cell lines in the nanoparticle cell interaction duration of 6 h, including liver cancer HepG-2 cell line, colon cancer HCT8, HCT 15, and HCT 116 cell lines, lung cancer A549 cell line, and myomelanocytic cancer B16 cell line.
As shown in Table 2, we clearly differentiate the endo/lysosomes in 6 cancer cell lines, with the accuracy at 0.01 pH values, which is impossible to be done with the commercial intraocular pH indicator kits.
Moreover, we re-evaluated pH values in endo/lysosomes of the HepG2 cells before and after cultured with BPSi nanoparticles. We reached the conclusion that BPSi uptake significantly increases the endo/lysosome pH values, from 4.70 ± 0.09 to 5.59 ± 0.05, perfectly illustrating the reason for BPsi uptaken induced the autophagy initially then terminated the autophagy flux. The intracellular uptake of BPSi makes the quantities of the endo/lysosomes increased, which was consistent with the results of gene sequencing, that autophagy-related genes (TFEB-CLEAR) were activated. Meanwhile, the autophagy termination by the increased pH values in endo/lysosomes also coincide with the results of p62 proteins upregulation in Western blot study.
Обсуждение
Nanoparticles can generally cause autophagy in cells [23], and studies have shown that the autophagic response to nanoparticles presenting a neutral or anionic surface involves enhanced clearance of autophagic cargo. Cell exposure to nanoparticles presenting a cationic surface, on the other hand, results in transcriptional upregulation of the TFEB pathway, but also causes lysosomal dysfunction, ultimately resulting in blockage of autophagic flux [7]. And our results are in consistent with these previous conclusions. In our study, we found that the expression of autophagy-related genes and proteins in HepG2 cells has been increased after feeding of BPSi nanoparticles through transcriptome sequencing, RT-qPCR, and Western experiments. However, the expression level of autophagy-related P62 protein does not decrease as the autophagy is activated. We suspect that the PEG-amine on the surface of BPSi nanoparticles raises the pH value of the lysosome, resulting in inhibition of P62 degradation. Existing lysosomal pH indicators cannot verify our guess. To accurately measure the lysosomal pH of living cells, we established a new method for endo/lysosomes pH qualitative determination based on self-decomposable nanoparticle systems. Ten nanoparticle systems with specific MB/TEOS parameters were employed for obtaining optimized pH sensitively responsive measurement method. The radial MB concentration gradient from inner out served as a major driving force for MB release. The drug release proceeded with simultaneously carrier decomposition, which was driven by a diffusion-controlled mechanism. Moreover, as the pH value decreases, the hydrogen ion concentration increases, and the enhanced electrostatic interaction promotes inner MB to release faster than in neutral solution [24]. The optimized central hollow nanoparticle system could release the central concentrated MB as a linear function of precise pH values in the range of pH 4.0–4.8, which is exactly the pH of lysosomes. Finally, by this qualitative pH indicator based on self-decomposable nanoparticles, we have succeeded in the detection of the average pH values of lysosomes in 6 cell lines. Moreover, by this system, we can qualitatively differentiate the pH changes of lysosomes before and after BPSi nanoparticle endocytosis by HepG-2 cells, clarifying the mechanism of the autophagy occurrence and then termination after BPSi endocytosis. The self-decomposable nanoparticle systems pave a brand new way for studying the luminal pH values, providing new tools to know better of the cell signaling and metabolism, and then providing new ways and methods for the treatment of cancer [25, 26].
Заключение
In this study, we found that BPSi can promote cell autophagy through transcriptome sequencing, but the amino groups on the surface of the nanoparticles can increase the pH of the lysosome and inhibit the degradation of autophagic flow. Thus, the lysosome pH significantly influences the autophagy stages. And precisely acquiring the information of lysosome pH will promote the perceiving of autophagy. However, the existing fluorescent lysosomal pH indicators could only determine a wide range of lysosomal pH; thus, we established a precise lysosomal pH indicator based on the self-dissociation system. By adjusting the synthesis parameters of MB@SiO2 , the release of MB loaded on the nanoparticles was linearly and negatively correlated with pH. And the nanoparticles mainly stay in the lysosome after entering the cell. By measuring the amount of MB released in the cells, the pH value of the lysosome can be calculated exactly according to the linear function. The established precise pH indicator provided a brand new tool and methodology to precisely study the lysosome pH values and further acquire more information on autophagy.
Доступность данных и материалов
Все данные, полученные или проанализированные в ходе этого исследования, включены в эту опубликованную статью и файлы с дополнительной информацией к ней.
Сокращения
- BPSi:
-
Black mesoporous silicon
- FBS:
-
Фетальная бычья сыворотка
- GO:
-
Gene ontology
- HCl:
-
Hydrochloric acid
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- МБ:
-
Methylene blue
- NaBr:
-
Sodium bromide
- NaSi:
-
Sodium silicide
- NH4Br:
-
Ammonium bromide
- НП:
-
Наночастицы
- OD:
-
Оптическая плотность
- PBS:
-
Физиологический раствор с фосфатным буфером
- PDMPO:
-
2-(4-Pyridyl)-5-((4-(2-dimethylaminoethy- laminocarbamoyl) methoxy) phenyl) oxazole
- RT-qPCR:
-
Reverse transcription quantitative polymerase chain reaction
- TEOS:
-
Тетраэтилортосиликат
Наноматериалы
- Мониторинг приложений SaaS:вид на лес и деревья
- Разработка и применение стекловолокна
- Получение наночастиц mPEG-ICA, нагруженных ICA, и их применение в лечении LPS-индуцированного повреждения клеток H9c2…
- Ультратонкий идеальный поглотитель и его применение в качестве плазмонного датчика в видимой области
- Потенциальная токсичность наночастиц диоксида титана для печени, мозга и эмбрионов у мышей
- Формирование и люминесцентные свойства нанокомпозитов Al2O3:SiOC на основе наночастиц оксида алюминия, модифици…
- Получение наноструктуры Au @ TiO2 - оболочка и ее применение для разложения и обнаружения метиленового синего
- Зеленая способность к синтезу и стабилизации наночастиц меди:каталитическая, антибактериальная, цитотоксич…
- Исследования TEM и STEM морфологии поперечного сечения двух- / трехслойных широкополосных антиотражающих пленок…
- Пленочный конденсатор:свойства, конструкция и применение