


Флуоресцентные нанокластеры неогликопротеина золота:синтез и применение в зондировании лектина растений и визуализации клеток
Аннотация
Взаимодействие углеводов и белков опосредует фундаментальные биологические процессы, такие как оплодотворение, передача сигналов между клетками или связь между хозяином и патогеном. Однако из-за огромной сложности событий распознавания гликанов в последние годы появляются новые инструменты, позволяющие их анализировать или применять. Здесь мы описываем первый препарат функционализированных неогликопротеином флуоресцентных нанокластеров золота, содержащих двухантенный N-гликан G0 в качестве целевой молекулы, овальбумин в качестве носителя / модельного антигена и флуоресцентное золотое ядро в качестве зонда для визуализации (G0-OVA-AuNCs). Впоследствии мы демонстрируем полезность созданных G0-OVA-AuNC для специфического восприятия лектинов растений и получения изображений дендритных клеток in vitro.
Введение
Золотые нанокластеры (AuNC), образованные от десяти до сотен атомов золота, привлекли внимание научного сообщества благодаря своим привлекательным химическим и физическим свойствам [1]. Менее 3 нм, нанокластеры золота приближаются к длине волны Ферми электронов, вызывая зависящее от размера флуоресцентное излучение и предлагая возможности в качестве зондов для зондирования и визуализации для приложений in vitro и in vivo [2,3,4]. Современные флуоресцентные анализы в основном используют органические красители, такие как родамин или флуоресцеин, или, реже, квантовые точки [5,6,7]. Однако из-за низкой фотохимической стабильности, pH-чувствительной флуоресценции или плохой растворимости в воде некоторых органических красителей и токсичности квантовых точек их использование может быть скомпрометировано [8]. В этом контексте нанокластеры золота можно рассматривать как альтернативные ультрамалые флуорофоры, лишенные упомянутых выше ограничений. Кроме того, AuNC характеризуются большим стоксовым сдвигом и длиной волны излучения флуоресценции от красного видимого до ближнего инфракрасного (ИК) диапазона, что очень удобно при биоимиджинге, поскольку перекрывает окно прозрачности ткани [8,9,10]. P>
Синтез нанокластеров золота с участием белков был впервые описан в 2009 году с использованием бычьего сывороточного альбумина (БСА) [11], и с тех пор водорастворимые защищенные белком нанокластеры стали новой тенденцией в нанонауке [9, 12, 13]. Намного меньше внимания уделялось гликопротеинам [14] для получения AuNCs, и нам не известны какие-либо сообщения, описывающие образование AuNC из синтетических неогликопротеинов в качестве каркасов. В общем, гликозилирование модулирует физико-химические свойства гликопротеинов, например фолдинг, время жизни в кровотоке или стабильность. Он также влияет на важные биологические функции белка, такие как распознавание рецептора-лиганда. Таким образом, создание синтетических неогликопротеинов из белков путем химического присоединения тщательно разработанных и охарактеризованных углеводов [15] может наделить их новыми функциональными свойствами для биологических применений. Поскольку углеводы участвуют в большом количестве различных биологических процессов через взаимодействие с углеводсвязывающими белками [16, 17], мы рассматриваем нанокластеры золота неогликопротеинов как новые зонды для изучения и использования событий распознавания углеводов как in vitro, так и in vivo [18]. В связи с этим наличие нескольких копий гликанов на поверхности белка обеспечивает поливалентное представление углеводов с последующим усилением аффинности связывания.
Здесь мы исследуем использование самофлуоресцентных неогликопротеиновых функционализированных AuNC в качестве датчиков для определения лектинов растений и в качестве реагентов для нацеливания на лектиновые рецепторы для визуализации дендритных клеток (DC) in vitro. Лектины - это белки, связывающие углеводы, которые помогают в различных явлениях биологического распознавания. У высших растений, например, лектины предотвращают появление растительноядных организмов путем распознавания и агглютинации чужеродных гликопротеинов, подавляя их рост и размножение [19]. В DCs млекопитающих, с другой стороны, рецепторы лектина C-типа (CLR), экспрессирующиеся на поверхности клетки, играют основную роль в распознавании патогенов [20]. Декорированные гликанами антигены распознаются специфическими CLR, которые затем подвергаются эндоцитозу, процессингу и, в конечном итоге, представляются Т-клеткам, вызывая специфические иммунные ответы. В нашем предыдущем исследовании мы показали [21], что функционализация модельного антигена, овальбумина (OVA), с помощью синтетического двухантенарного GlcNAc, завершающего N-гликан G0, усиливает нацеливание на DC и последующее поглощение и презентацию антигена. Мы предположили, что вышеупомянутый феномен инициируется взаимодействием гликана G0 и эндоцитарных рецепторов лектина С-типа, экспрессируемых на поверхности ДК. Следовательно, мы считаем, что флуоресцентные и поливалентные нанокластеры золота G0-OVA могут стать альтернативой флуоресцентно меченым G0-OVA, применявшемуся в нашем предыдущем исследовании, и могут быть использованы в качестве нового инструмента для визуализации DC. Более того, опосредованное гликаном нацеливание на DC, которое позволяет инициировать сильные Т-клеточные иммунные ответы, можно использовать для повышения эффективности вакцин-кандидатов [22,23,24]. В этом первоначальном исследовании мы представим синтез нанокластеров золота, начиная с неогликопротеинов, и оценку функциональности и доступности гликанов G0 с использованием экспериментов по агглютинации лектина. Наконец, мы продемонстрируем потенциал флуоресцентных нанокластеров золота для визуализации дендритных клеток.
Результаты и обсуждение
Мы оптимизировали синтез золотых нанокластеров овальбумина, украшенных G0-гликаном (G0-OVA-AuNCs), на основании предыдущих отчетов с использованием неконъюгированного белка OVA [25, 26]. AuNC, защищенные белком, получали добавлением тетрахлорозавр (III) кислоты золота (HAuCl 4 . 3H 2 О) до белкового раствора, а затем 1 М водный раствор гидроксида натрия. Повышение pH реакции увеличивает потенциал восстановления остатков триптофана и тирозина, присутствующих в OVA (рис. 1а) [14]. Белковый каркас действует как восстанавливающий и стабилизирующий реагент, захватывая небольшой кластер золота внутри структуры белка и изолируя его от окружающей среды. Предыдущий протокол синтеза флуоресцентных OVA-AuNC требовал очень высококонцентрированного раствора OVA (до 65 мг / мл -1 ) [25]. Чтобы ограничить использование ценных неогликоконъюгатов OVA для получения AuNC, мы определили минимальное количество неконъюгированных OVA, которые могут эффективно продуцировать OVA-AuNC. Мы обнаружили, что при концентрации OVA 15 мг / мл -1 было достаточно для получения кластеров с сильным флуоресцентным излучением (дополнительный файл 1:Рисунок S1). Образование OVA-AuNC значительно ускорялось микроволновым облучением, что сокращало время реакции с 18 ч до 6 мин [25]. Кроме того, микроволновое облучение обеспечивало однородный нагрев раствора, способствуя образованию однородных и монодисперсных кластеров [1]. OVA-AuNC, полученные таким образом, имеют бледно-коричневый цвет и излучают сильную красную флуоресценцию при освещении УФ-светом (рис. 1b). В УФ-видимом спектре OVA-AuNC отсутствует оптическая плотность, соответствующая полосе локализованного поверхностного плазмонного резонанса, что указывает на отсутствие наночастиц золота размером более 5 нм [27], что было дополнительно подтверждено просвечивающей электронной микроскопией (ПЭМ). Мы измерили средний диаметр золотого ядра OVA-AuNCs в диапазоне 1,9 ± 0,7 нм (дополнительный файл 1:Рисунок S2). Наконец, средний гидродинамический размер OVA-AuNC, равный 8,7 нм ± 2,5 нм, был определен с помощью динамического светорассеяния (дополнительный файл 1:Рисунок S2). Его размер, подобный OVA (диаметр 6,5–7 нм [28]), предполагает присутствие одной молекулы белка на золотое ядро. Дальнейший анализ OVA-AuNC с помощью электрофореза в агарозном геле показал сходную подвижность OVA и OVA-AuNC по направлению к положительному электроду, что дополнительно подтверждает одинаковый размер для обоих видов и их отрицательный заряд при нейтральном pH (рис. 1c). Спектр испускания флуоресценции OVA-AuNC показывает максимум испускания при λ 670 нм при возбуждении на длине волны 350 нм (рис. 1d). Спектры возбуждения AuNC широкие, а стоксов сдвиг большой (более 200 нм), что идеально подходит для спектрального многоцветного детектирования в присутствии другого флуоресцентного зонда (мультиплексирование), характеризующегося аналогичной длиной волны возбуждения, такого как синий излучающий краситель Alexa Fluor® 405. [26]. Расчетный квантовый выход (QY) OVA-AuNCs составил ≈ 4% при использовании флуоресцеина в 0,1 М NaOH (QY =≈ 92%) в качестве эталона [29]. Степень окисления золотого ядра была приписана смеси частиц Au (I) и Au (0) на основании измерений рентгеновской фотоэлектронной спектроскопии (XPS) (дополнительный файл 1:Рисунок S3) [11, 30]. Изучение кругового дихроизма вторичной структуры белка (CD) выявило случайную организацию клубков, предполагающую потерю естественной складки для OVA в AuNCs, вероятно, из-за жестких щелочных условий во время синтеза (дополнительный файл 1:Рисунок S4) [31]. Наконец, мы также наблюдали необычайную стабильность OVA-AuNC в широком диапазоне pH (3–11), а также в растворе, содержащем фетальную бычью сыворотку (FBS), наиболее часто используемую сывороточную добавку для культивирования клеток in vitro. (Дополнительный файл 1:Рисунок S5). Эта особенность нанокластеров золота OVA подчеркивает их полезность для биоанализов in vitro и in vivo и открывает новые захватывающие возможности. Например, нечувствительное к pH флуоресцентное излучение OVA-AuNCs могло бы обеспечить эффективное отслеживание частиц внутри клетки без потери сигнала даже внутри эндосомных компартментов, характеризующихся слегка кислым pH [32, 33]. Напротив, в этих условиях использование определенных конъюгатов флуоресцеина может быть затруднено, поскольку максимальное излучение флуоресценции этих красителей достигается при основном pH. OVA-AuNC также показали отличную растворимость как в воде, так и в среде для роста клеток. Полная солюбилизация OVA-AuNC наблюдалась до концентрации до 40 мг / мл как в воде, так и в среде для роста клеток. (Дополнительный файл 1:Рисунок S6). Спектры испускания флуоресценции при различных разведениях OVA-AuNC были измерены и исследованы на предмет их стабильности при инкубации при 37 ° C до 24 часов.
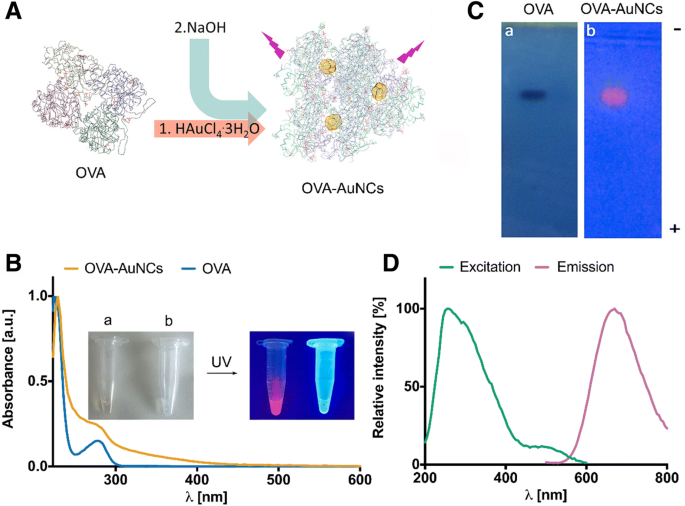
а Схематическое изображение синтеза OVA-AuNCs. б УФ-видимые спектры OVA (синяя линия) и OVA-AuNC (оранжевая линия). Вставка:изображения OVA-AuNC ( a ) и OVA ( b ) при освещении видимым светом (слева) и при освещении УФ-светом (365 нм) (справа). c Электрофорез в агарозном геле OVA (окрашенного кумасси синим G-250) и OVA-AuNC (при освещении УФ-светом). г Спектры возбуждения (зеленая линия) и испускания (розовая линия) флуоресценции OVA-AuNC
Для синтеза AuNC, защищенных неогликопротеином G0-OVA, мы первоначально получили функционализированные G0 неогликопротеины OVA. Двуантенный N-гликан G0, снабженный амино-линкером C5, был синтезирован, как описано ранее [34], и конъюгация с OVA была достигнута с использованием сложного эфира дисукцинимидилсуберата (DSS) в качестве сшивающего реагента (рис. 2a) [21, 35]. Вкратце, конъюгация выполняется в виде двухэтапной реакции:N-гликан G0 функционализируется 13-кратным избытком линкера DSS и затем связывается со свободными аминогруппами на OVA. Контролируя соотношение гликанов / белков, мы могли эффективно регулировать степень замещения OVA (см. SI). Успешное образование неогликопротеинов было подтверждено появлением диффузной электрофоретически более медленной мигрирующей полосы на геле SDS-PAGE (рис. 2b), тогда как среднее количество введенных гликанов было дополнительно определено с помощью масс-спектрометрии MALDI-TOF [21] (рис. . 2в). Следуя этой стратегии, мы получили неогликоконъюгаты OVA, отображающие 2–3, 3–4 и 5–6 копий N-гликана G0 на белок.
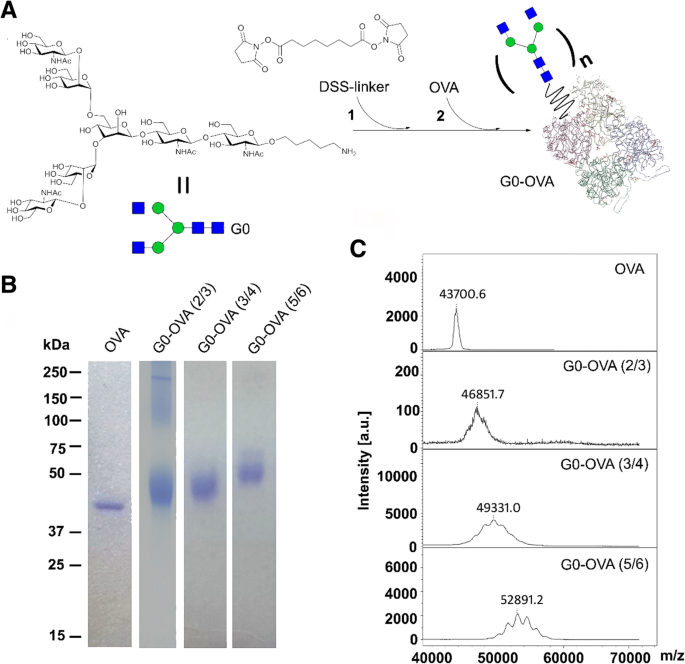
а Синтез неогликоконъюгатов G0-OVA ( n =валентность). б Гель-электрофорез в SDS-PAGE неогликопротеинов OVA и G0-OVA, содержащих разную валентность N-гликана G0. c MALDI-TOF масс-спектры неогликопротеинов OVA и G0-OVA, содержащих разную валентность N-гликана G0
Затем мы использовали синтетические неогликопротеины для получения AuNC (рис. 3а) в ранее оптимизированных условиях и исследовали влияние функционализации гликана и его валентности на физические и оптические свойства кластеров. Как показано на фиг. 3, поглощение в УФ-видимой области и флуоресцентное излучение G0-OVA-AuNC были идентичны OVA-AuNC с характеристическим поглощением при 278 нм и эмиссией красной флуоресценции с максимумом около 670 нм (рис. 3b, c).
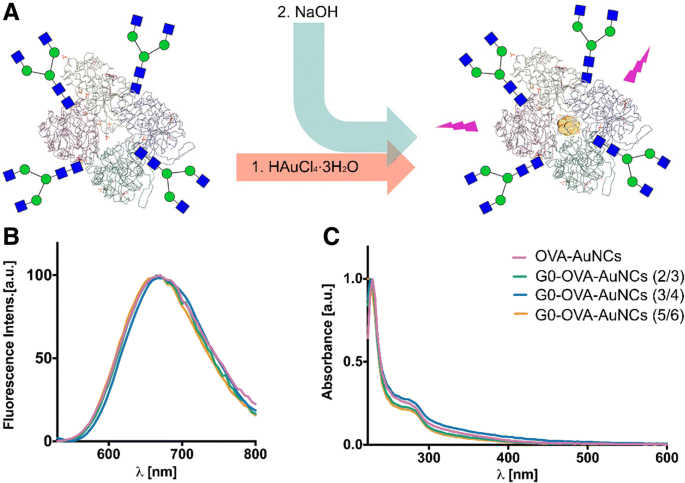
а Схематическое изображение синтеза OVA-AuNC на основе G0-гликана. б Спектры флуоресценции G0-OVA-AuNC (синие, оранжевые и зеленые линии) по сравнению с OVA-AuNC (розовая линия). c УФ-видимые спектры G0-OVA-AuNC (синие, оранжевые и зеленые линии) и OVA-AuNC (розовая линия)
Кроме того, с помощью измерений ПЭМ мы установили средний диаметр 1,6 ± 0,5 нм (рис. 4) для ядра G0-OVA-AuNC, что сопоставимо с размером OVA-AuNC (1,9 ± 0,7 нм). Таким образом, основываясь на наших результатах, мы думаем, что гликаны, конъюгированные с OVA через остатки лизина или N-конец, не влияют на общий восстановительный потенциал гликопротеинов и дальнейшее образование кластеров при проверенных валентностях [36]. Кроме того, кажется, что сахара не препятствуют образованию кластер-стабилизирующих тиолатных полимеров Au (I) [37, 38] между цистеиновыми группами белка и золотым ядром, даже если они присутствуют в большем количестве (5–6 копий).
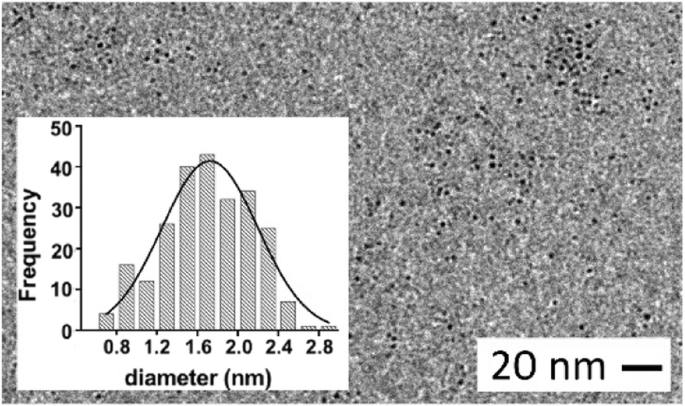
ПЭМ-изображение G0-OVA-AuNC (3/4). Вставка:распределение диаметров золотого сердечника G0-OVA-AuNC по размерам (3/4)
Наконец, аналогично OVA-AuNCs, вторичная структура белка G0-OVA-AuNCs (3/4) была назначена CD и выявила случайную организацию клубков (дополнительный файл 1:Рисунок S4). Однако, прикрепляя гликаны к белковой структуре AuNC, мы вводим нацеленные молекулы, обеспечивающие взаимодействие с углеводсвязывающими белками, и в то же время белок OVA остается только в качестве носителя, конформационные изменения которого не изменят распознавание лектином целая система. Описана антигенная способность денатурированного OVA к продукции антител у мышей [39] и к активации Т-клеток. Активация Т-клеток не зависит от вторичной структуры антигена, поскольку опосредуется узнаванием определенных аминокислотных последовательностей, которые представлены как в нативном, так и в денатурированном OVA [40]. Фактически, пептид антигена OVA 323-339 отвечает за основной специфический ответ Т-клеток на OVA, и этот пептид был использован в наночастицах для индукции иммунных ответов [41].
Чтобы исследовать функциональность вновь введенных гликановых цепей на нанокластеры золота, защищенные неогликопротеином, мы исследовали их взаимодействие с различными растительными лектинами в экспериментах по агглютинации. Мы провели инкубацию G0-OVA-AuNC и OVA-AuNC с двумя разными растительными лектинами:лектином-II Bandeiraea simplicifolia (BSL-II), специфичным для концевых фрагментов GlcNAc, и Solanum tuberosum лектин (STL), распознающий хитобиозу ядра, присутствующую в N-гликанах [34, 42]. Кроме того, чтобы исключить любые неспецифические взаимодействия между G0-OVA-AuNC и лектинами, Aleuria aurantia лектин (AAL), специфичный для L-фукозы, был включен в качестве контроля. Мы добавляли возрастающие концентрации лектинов BSL-II, STL и AAL в раствор G0-OVA-AuNC (5/6). После инкубации в темноте растворы центрифугировали и измеряли флуоресценцию супернатанта. Как показано на рис. 5, мы могли наблюдать уменьшение интенсивности флуоресценции в зависимости от концентрации после инкубации с BSL-II и STL (рис. 5a – d), в то время как интенсивность флуоресценции оставалась неизменной в присутствии AAL (рис. . 5e). После инкубации G0-OVA-AuNC (5/6) с STL наблюдался видимый осадок при облучении УФ-лампой, тогда как при инкубации с AAL осадок не появлялся (рис. 5f). Связь начальных значений флуоресценции (F0) с конечными значениями (F) после инкубации лектинов была представлена в зависимости от возрастающих концентраций лектинов (рис. 5b, d). Взаимодействие STL с G0-OVA-AuNC (5/6) показало линейность от 0 до 7,5 мкМ, и предел обнаружения был рассчитан на основе уравнения SD * 3 / S (где SD - стандартное отклонение калибровочной кривой и S - значение наклона) [12, 13], равное 2,35 мкМ ( y =1,95 x + 0,4266, R 2 =0,97). Таким же образом взаимодействие BSL-II с G0-OVA-AuNC (5/6) показало линейность от 0 до 10 мкМ, а предел обнаружения (LOD) был рассчитан как 2,83 мкМ ( y =0,5687 x + 0,5838, R 2 =0,965). Интересно, что кластеры, содержащие меньшее количество конъюгированных сахаров, таких как G0-OVA-AuNC (2/3) (дополнительный файл 1:рисунок S7, AB), не показали какого-либо значительного изменения интенсивности флуоресценции при добавлении BSL-II и STL, что указывает на отсутствие агглютинации. Такое поведение подчеркивает важное влияние поливалентного представления гликанов на сшивающую активность лектинов, даже когда применяются лишь небольшие различия в плотности гликанов. Важно отметить, что никаких изменений в интенсивности излучения флуоресценции неконъюгированных OVA-AuNC, инкубированных с лектинами, не наблюдалось (дополнительный файл 1:рисунок S7, C-D). Несмотря на то, что нативный куриный OVA содержит единственный N-связанный сайт гликозилирования (Asn-292), преимущественно замещенный высоким содержанием маннозы или менее распространенными гибридными и сложными N-гликанами [43], эта модификация сахара не распознается ни STL, ни BSL-II. , вероятно, из-за моновалентного предлежания. Это подтвердило наличие специфического углевод-лектинового взаимодействия между химически введенным G0 на G0-OVA-AuNC и STL / BSL-II и исключило неспецифическое связывание OVA-AuNC с лектинами. Мы предполагаем возможное применение этой системы для обнаружения биомаркеров углеводсвязывающих белков. Тем не менее, получение неогликопротеинов, демонстрирующих большое количество копий гликана, повысило бы привязанность конструкции к желаемому лектину, улучшив предел обнаружения [44].
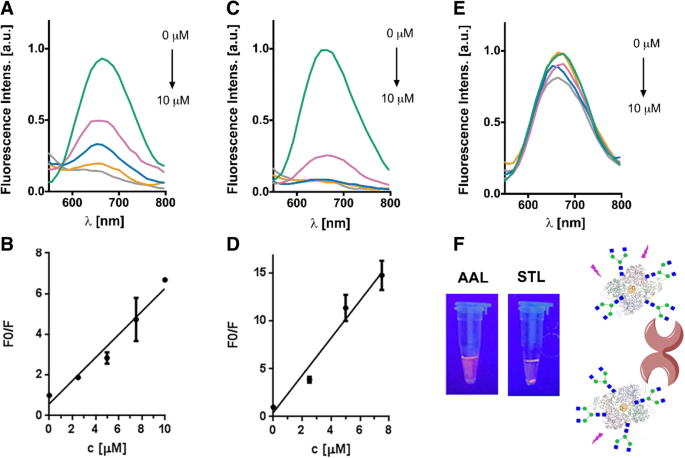
Анализ лектиновой агглютинации G0-OVA-AuNC (5/6). а Типичные спектры флуоресценции супернатантов растворов OVA-G0 (5/6) -AuNCs после инкубации с BSL-II. б Представление F0 / F против возрастающих концентраций BSL-II. c Типичные спектры флуоресценции супернатантов растворов OVA-G0 (5/6) -AuNCs после инкубации с STL. г Представление F0 / F против возрастающих концентраций STL. е Типичные спектры флуоресценции супернатантов OVA-G0 (5/6) -AuNCs после инкубации с AAL. е Изображение, соответствующее инкубации G0-OVA-AuNC (5/6) с AAL и STL при освещении УФ-светом (365 нм). После инкубации с STL образовался видимый осадок. Справа:схематическая иллюстрация связывания между G0-OVA-AuNC и растительным лектином
Мы также изучили взаимодействие G0-OVA-AuNC (5/6) с STL в присутствии среды для роста клеток, чтобы различить возможное влияние компонентов среды на взаимодействия углеводов и белков. G0-OVA-AuNC (5/6) растворяли в полной модифицированной Iscove среде Дульбекко (IMDM) с добавлением фетальной телячьей сыворотки и инкубировали с возрастающими количествами STL; полученные растворы анализировали электрофорезом в агарозном геле. (Дополнительный файл 1:Рисунок S8). Электрофоретическая подвижность G0-OVA-AuNC (5/6) сохраняется как в воде, так и в комплексной среде. Тем не менее, в присутствии возрастающих количеств STL наблюдалось дозозависимое смещение G0-OVA-AuNC (5/6) к отрицательному полюсу, подчеркивая взаимодействие белков и углеводов даже в присутствии сложных клеточных сред. Это указывает на то, что взаимодействию белков и углеводов с G0-OVA-AuNC (5/6) не препятствует присутствие компонентов среды.
Потенциал кластеров для флуоресцентной визуализации и предыдущий успех с нацеливанием на DCs с помощью неогликопротеинов G0-OVA [21] побудили нас изучить G0-OVA-AuNCs в захвате мышиными DCs in vitro. Мы использовали конфокальную флуоресцентную микроскопию для визуализации интернализации самофлуоресцентных G0-OVA-AuNC (3/4). ДК селезенки выделяли от мышей C57BL / 6J и CD11c + Популяцию очищали с помощью сортировки магнитно-активированных клеток (MACS) (дополнительный файл 1:фиг. S9) и высевали на стеклянные покровные стекла, покрытые поли-d-лизином, в течение ночи. В качестве доказательства концепции очищенные DC инкубировали с G0-OVA-AuNC (3/4), промывали для удаления несвязанных материалов и фиксировали. После 40 мин инкубации были получены изображения конфокальной флуоресцентной микроскопии (рис. 6), и мы наблюдали сильную флуоресценцию дендритных клеток, инкубированных с G0-OVA-AuNC (3/4), демонстрируя их эффективную интернализацию (рис. 6a) и отсутствие флуоресценции. в отрицательном контроле (нестимулированный CD11 + клетки; Дополнительный файл 1:Рисунок S9). Интернализация нанокластеров была дополнительно подтверждена наложением отдельных изображений постоянного тока, сделанных вдоль их Z -оси (z-стек) и путем реконструкции трехмерного изображения одного постоянного тока для визуализации флуоресцентного излучения, исходящего изнутри клетки (рис. 6b и дополнительный файл 1:рис. S10).
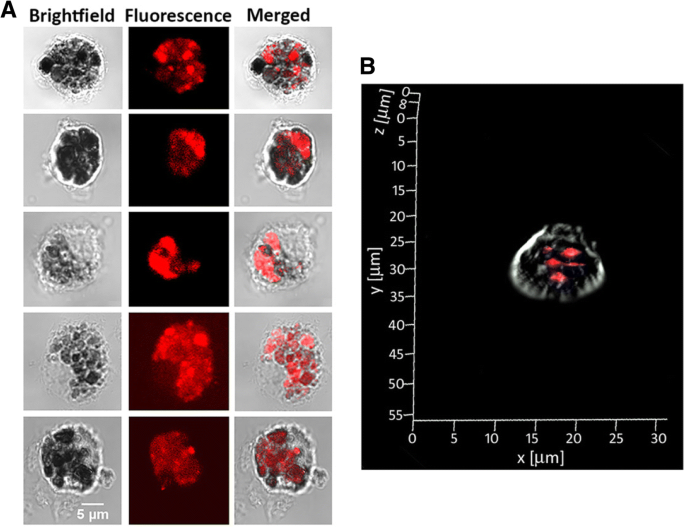
Поглощение G0-OVA-AuNC (3/4) мышиными DC, измеренное с помощью конфокальной микроскопии. а Типичные изображения CD11c + клетки после инкубации с G0-OVA-AuNC (3/4). б Изображение Z-стека одной дендритной клетки, представляющее поглощение G0-OVA-AuNC внутри клетки
Таким образом, мы подготовили и охарактеризовали защищенные неогликопротеином нанокластеры золота, которые были использованы в экспериментах по агглютинации и в анализе поглощения DC. На примере определения специфического лектина растений мы продемонстрировали полезность G0-OVA-AuNC для анализа углеводно-белковых взаимодействий. Мы также показали in vitro визуализацию дендритных клеток с помощью флуоресцентной микроскопии. Однако необходимо провести дальнейшие эксперименты, чтобы, во-первых, лучше понять роль гликана в поглощении нанокластеров DC (например, путем отслеживания локализации нанокластеров в клеточных компартментах), а во-вторых, изучить их био- и цитосовместимость. Основываясь на наших предыдущих результатах, описывающих свойства нацеливания гликана G0 и увеличение поглощения G0-OVA DC по сравнению с неконъюгированными OVA, мы считаем, что G0-OVA-AuNC могут стать привлекательной альтернативой флуоресцентно меченным неогликопротеинам.
Выводы
В заключение, в этом первоначальном исследовании мы представляем первый синтез нанокластеров золота, защищенных неогликопротеином, и оценку их физических и оптических свойств по сравнению с кластерами неконъюгированных белков. Мы подтвердили доступность сахара G0 на AuNC в анализах агглютинации путем специфических взаимодействий с растительными лектинами, что дополнительно подчеркнуло важность минимальной плотности гликанов для эффективной сшивающей активности лектинов. В качестве доказательства концепции мы также продемонстрировали пригодность G0-OVA-AuNC для визуализации модельных мышиных DC.
Мы полагаем, что самофлуоресцентные AuNC, функционализированные неогликопротеином, могут стать привлекательными инструментами, позволяющими анализировать и применять углеводно-белковые взаимодействия. Благодаря своим уникальным физическим и химическим свойствам, таким как большие сдвиги Стокса, растворимость в воде или стабильность pH, G0-OVA-AuNC могут стать альтернативой меткам органических красителей и использоваться для визуализации DC, исследований поглощения углеводов и растений. лектиновое зондирование. Наконец, используя иммуногенный носитель, такой как OVA, для прикрепления гликанов, функционализированные неогликопротеином нанокластеры золота могут не только позволить провести углубленные исследования поглощения, процессинга и презентации антигена, но и в будущем могут найти применение в качестве флуоресцентных терапевтических средств или адъювантные молекулы.
Методы / экспериментальные
Материалы
Все растворы были приготовлены в наночистой воде (18 МОм · см) из системы очистки воды Diamond UV (Branstead International, IA, Мадрид, Испания). Вся стеклянная посуда, используемая для приготовления AuNC, была предварительно промыта раствором царской водки (HNO 3 :HCl, 1:3, v / v ) и тщательно промыть водой с наночастицами. Тригидрат хлорида золота (III), гидроксид натрия и триэтиламин были приобретены у Sigma Aldrich. Овальбумин, не содержащий ЛПС, был приобретен у Hyglos (Бернрид, Германия). Суберат дисукцинимидила (DSS) и ДМСО были приобретены в Thermo Fisher Scientific. N-гликан G0 был химически синтезирован, как описано ранее [21].
Синтез AuNC. Общая процедура
К перемешиваемому раствору OVA или неогликопротеина (15 мг мл -1 ) в наночистой воде водный 0,1 М раствор HAuCl 4 · 3H 2 По каплям добавляли О (4,2 мМ, конечная концентрация). Полученную смесь перемешивали при комнатной температуре в течение 5 минут и по каплям добавляли водный 1 М раствор NaOH (конечная концентрация 150 мМ) для увеличения pH смеси. Полученные растворы инкубировали при 100 ° C в течение 6 мин в микроволновом реакторе Biotage® Initiator. Диализ AuNC проводили на диализных кассетах Slide-Z-lyser TM (10 K MWCO) от Thermo Fisher Scientific над наночистой водой.
Спектры флуоресценции синтезированных OVA и AuNC, защищенных неогликопротеином, измеряли с использованием 96-луночных планшетов из полистирола Nunc ™ на ридере Varioskan Flashmicroplate (Thermo Scientific) с длиной волны возбуждения при λ 350 нм, работающей с программным обеспечением ScanIt.
Электрофорез OVA-AuNC и белка OVA в агарозном геле проводили в 0,75% -ном агарозном геле при 80 В в течение 30 мин. Визуализацию OVA-защищенных AuNC проводили при облучении УФ-светом (365 нм), и белок OVA окрашивали кумасси синим G-250.
Получение изображений ПЭМ проводили на автоэмиссионном ПЭМ JOEL JEM-2100F с ускоряющим напряжением 200 кВ. Растворы AuNC наносили методом капельного литья на медные сетки ПЭМ, покрытые ультратонкой углеродной подложкой (Ted Pella, Redding, США). Средний диаметр золотого нанокластера был определен количественно с помощью ImageJ (Java).
Инкубация G0-OVA-AuNC (5/6) с лектинами растений
Десять микролитров G0-OVA-AuNC (5/6) [0,2 мг мл -1 ] в буфере TSM (20 мМ Трис · HCl, 150 мМ NaCl, 2 мМ CaCl 2 , 2 мМ MgCl 2 , pH =7,4) помещали на 384-луночный планшет из полистирола с черным покрытием Nunc ™. Впоследствии 10 мкл Aleuria aurantia лектин (AAL), Solanum tuberosum лектин (STL) и лектин-II Bandeiraea simplicifolia (BSL-II) в буфере TSM добавляли, в результате чего конечные концентрации белка составляли 0, 2,5, 5, 7,5, 10 мкМ. Соответствующие растворы инкубировали в течение ночи в темноте при осторожном встряхивании. Растворы белков центрифугировали при 11000 g в течение 1 ч и спектры флуоресцентного излучения супернатантов записывали с помощью считывающего устройства для микропланшетов Varioskan Flash (Thermo Scientific) с длиной волны возбуждения λ 350 нм. Для наблюдения за осаждением при образовании комплексов AuNCs-лектин был проведен контрольный эксперимент. Затем 10 мкл G0-OVA-AuNC (5/6) [0,2 мг мл -1 ] инкубировали в течение 1 часа с 10 мкл AAL и STL [40 мкМ] с последующим центрифугированием при 11000 g (1 час). Изображения были получены при облучении УФ-светом (365 нм).
Выделение дендритных клеток селезенки мыши
Дендритные клетки мышей, использованные во всех экспериментах, были выделены от мышей C57BL / 6J, выведенных Charles River (CIC biomaGUNE, Сан-Себастьян, Испания). Протоколы экспериментов на животных были одобрены комитетом по этике животных CIC biomaGUNE и проводились в соответствии с руководящими принципами ARRIVE и Директивами Европейского Союза по этике и благополучию животных. Мышей содержали в обычных условиях содержания (22 ± 2 ° C, влажность 55 ± 10% и 12-часовой цикл день / ночь) и кормили стандартной диетой ad libitum . Мышей анестезировали 2,5% изофлураном в 100% O 2 и умерщвлен шейным вывихом.
Селезенки были получены от мышей C57BL / 6J ( n =3, женщина, 27–28 недель). Для выделения спленоцитов селезенки промывали модифицированной средой Дульбекко Искова (IMDM) с добавлением 2 мМ l-глутамина, 100 Ед / мл пенициллина, 100 мкг / мл стрептомицина и 10% фетальной телячьей сыворотки (FCS; PAN Biotech). Суспензию клеток хранили холодной и фильтровали через сетчатый фильтр для клеток 40 мкм для удаления агрегатов клеток. После центрифугирования (300 g, 5 мин, 4 ° C) осадки клеток ресуспендировали в 5 мл свежеприготовленного буфера для лизиса эритроцитов (10% 100 мМ Трис pH 7,5 + 90% 160 мМ NH 4 Cl), осторожно перемешивают и инкубируют при комнатной температуре в течение 2 мин. Клетки дважды промывали полной средой IMDM и центрифугировали перед ресуспендированием в буфере MACS (PBS, 0,5% BSA, 2 мМ EDTA). Дендритные клетки (CD11c + клетки) были выделены из суспензии клеток селезенки мышей C57BL / 6 методом магнитно-активированной сортировки клеток с использованием CD11c + MicroBeads (Miltenyi). Клетки, инкубированные с магнитными микрошариками, загружали на колонку MACS, помещенную в магнитное поле. Несвязанные ячейки прошли через столбец, оставаясь CD11c + клетки промывали буфером MACS и элюировали из колонки. Для повышения чистоты постоянного тока CD11c + очистку клеток повторяли. Суспензию клеток центрифугировали, ресуспендировали в полной среде IMDM и подсчитывали.
Использование контроллеров домена G0-OVA-AuNC (3/4)
CD11c + клетки мышей C57BL / 6J (2 × 10 6 клетки) высевали на покровные стекла, покрытые поли-d-лизином, в течение ночи. G0-OVA-AuNCs (3/4) were added to cells (150 μg mL −1 ) and incubated for 40 min at 37 °C. After incubation, cells were carefully washed with cooled PBS and fixed with 3% paraformaldehyde at RT for 20 min. After washing with PBS and water, the cover-slips were mounted on slides using Vectashield® mounting medium. Fluorescent images were taken using the Zeiss LSM 510 laser scanning confocal microscope (Carl Zeiss) equipped with a UV laser (365 nm) and × 63 oil immersion objective.
Сокращения
- AAL:
-
Aleuria aurantia lectin
- AuNCs:
-
Gold nanoclusters
- BSA:
-
Bovine serum albumin
- BSL-II:
-
Bandeiraea simplicifolia lectin-II
- CD:
-
Circular dichroism
- CLRs:
-
C-type lectin receptors
- DCs:
-
Dendritic cells
- DMSO:
-
Диметилсульфоксид
- DSS:
-
Disuccinimidyl suberate ester
- FBS:
-
Fetal bovine serum
- G0:
-
Biantennary N-glycan
- G0-OVA-AuNCs:
-
Neoglycoprotein functionalized gold nanoclusters
- HAuCl4 . 3 H 2 О:
-
Gold tetrachloroauric (III) acid
- IMDM:
-
Iscove’s modified Dulbecco’s medium
- IR:
-
Инфракрасный
- LOD:
-
Limit of detection
- MACS:
-
Magnetic-activated cell separation
- OVA:
-
Ovalbumin
- КГ:
-
Quantum yield
- STL:
-
Solanum tuberosum lectin
- ТЕМ:
-
Просвечивающая электронная микроскопия
- XPS:
-
Рентгеновская фотоэлектронная спектроскопия
Наноматериалы
- Взгляд на новейшие приложения в области зрения и визуализации
- Создание и визуализация Cyclocarbon
- Многофункциональные наночастицы золота для улучшенных диагностических и терапевтических применений:обзор
- Достижения и проблемы флуоресцентных наноматериалов для синтеза и биомедицинских приложений
- Синтез композита из электропроводных кремнеземных нановолокон / наночастиц золота с помощью лазерных импуль…
- Синтез и активность окисления CO 1D смешанного бинарного оксида CeO2-LaO x, нанесенного золотыми катализаторами
- Синтез и эффективность in vitro покрытых полипирролом железо-платиновых наночастиц для фототермической терапии…
- Синтез композита поли (3,4-этилендиокситиофен) / золото / графен в твердом состоянии и его применение для амперо…
- Стимуляция роста клеток SH-SY5Y наночастицами золота, модифицированными 6-меркаптопурином и проникающим в нейро…
- Сапонины Platycodon из Platycodi Radix (Platycodon grandiflorum) для зеленого синтеза наночастиц золота и серебра