


Экзосомная микроРНК-133a, полученная из мезенхимальных стволовых клеток костного мозга, сдерживает фиброз миокарда и эпителиально-мезенхимальный переход у крыс с вирусным миокардитом посредством подавления MAML1
Аннотация
Миокардит - заболевание, характеризующееся локальным или диффузным воспалением миокарда без эффективного лечения. В этом исследовании изучался регуляторный механизм микроРНК-133 (miR-133), секретируемой из экзосомы, полученной из мезенхимальных стволовых клеток костного мозга (BMSC-Exo), на фиброз миокарда и эпителиально-мезенхимальный переход (EMT) у крыс с вирусным миокардитом (VMC) посредством регулирования подобный вдохновителю 1 (MAML1). BMSC у крыс были изолированы и культивированы для определения их иммунного фенотипа, а также остеогенной и адипогенной способности, а BMSC-Exo были извлечены и идентифицированы. Экзосомы получали ультрацентрифугированием, которые идентифицировали с помощью просвечивающего электронного микроскопа и вестерн-блоттинга. Крысам вводили вирус Коксаки B3 для приготовления модели VMC, и кардиомиоциты выделяли, культивировали и группировали так же, как в экспериментах на животных (NC Exo , Ad-miR-133a Exo , Адас-miR-133a Exo ). Были проведены эксперименты in vivo и in vitro, чтобы выяснить роль экзосомальных miR-133a и MAML1 в воспалении, апоптозе, EMT, фиброзе и жизнеспособности клеток. Связь нацеливания между miR-133a и MAML1 была подтверждена двойным анализом репортерного гена люциферазы. BMSC-Exo повышал экспрессию miR-133a у крыс VMC и эффективно улучшал сердечную функцию крыс VMC и фиброз миокарда, увеличивал жизнеспособность кардиомиоцитов и подавлял процесс EMT. Повышенный уровень miR-133a в экзосомах усиливал улучшения. Заглушенная miR-133a эффективно обращала действие BMSC-Exo на крыс VMC. miR-133a нацелена на MAML1. Ингибирование MAML1 улучшает сердечную функцию и фиброз миокарда у крыс VMC и может обратить действие экзосом с молчанием miR-133a на крыс VMC. Наше исследование предполагает, что повышенный уровень miR-133a в экзосоме подавляет фиброз миокарда и ЭМП у крыс с VMC посредством подавления MAML1, тем самым подавляя прогрессирование миокардита.
Введение
Миокардит считается воспалительным заболеванием клеток сердечной мышцы [1]. Миокардит, очевидно, более распространен у мужчин, чем у женщин [2]. Вирусный миокардит (ВМК) является основным фактором, приводящим к дилатационной кардиомиопатии (ДКМП) и внезапной смерти у молодых людей [3]. Клинические проявления миокардита различны:от бессимптомных состояний с неоднозначными признаками и симптомами до серьезного разрушения миокарда вирусами и иммунными клетками, страдающими кардиогенным шоком и аритмиями [1]. Миокардит может быть вызван множеством инфекционных элементов, в том числе вирусами, бактериями, хламидиями, риккетсиями, грибами и простейшими, а также неинфекционными индукторами. Среди них вирусная инфекция была наиболее частой причиной, особенно у детей [4]. Вирус Коксаки B3 (CVB3), как наиболее важный вирус, приводящий к миокардиту, может вызывать реакцию окислительного стресса и апоптоз в патогенезе VMC, но о специальном лечении VMC еще не сообщалось [5]. Более того, патогенез VMC недостаточно документирован, а точное клиническое лечение также отсутствует [3]. Поэтому срочно необходимы новые цели для улучшения прогноза болезни.
МикроРНК (миРНК) представляют собой эндогенные некодирующие РНК, которые могут регулировать экспрессию генов, кодирующих белок [6]. MiR-133a, как одна из кардиоспецифичных miRNA, участвует в развитии сердца и некоторых сердечно-сосудистых заболеваниях, включая инфаркт миокарда (ИМ) [7]. Кроме того, miR-133a является той, которая аномально экспрессируется при хронической кардиомиопатии при болезни Шагаса [8]. Более того, уровень miR-133a в миокарде связан с воспалением, функцией левого желудочка и клиническим исходом воспалительной кардиомиопатии [9]. MiRNA были обнаружены в экзосомах, полученных из тучных клеток мыши и человека [10]. Экзосомы, наноразмерные везикулы, высвобождаемые большинством видов клеток, обнаруживаются в различных биологических жидкостях [11]. Экзосомы могут передавать свой груз клеткам-реципиентам, что, как было показано, изменяет биохимический состав и сигнальные пути клеток-реципиентов [12, 13]. Доказано, что измененные миРНК экзосом связаны с патогенезом миокардита, индуцированного CVB3 [14]. Было решено, что экзосомальный miR-125b-5p из обусловленных гипоксией мезенхимальных стволовых клеток костного мозга снижает апоптоз кардиомиоцитов и усиливает ишемическое восстановление сердца [15]. Более того, экзосомальный miR-25-3p из МСК облегчает ИМ за счет уменьшения апоптоза кардиомиоцитов и воспалительной реакции [16]. Интересно, что miR-133a, экспрессируемая экзосомами в сердце, связана с сердечным тропонин-сердечным тропонином-I [17]. Mastermind-like 1 (MAML1) был перекрестно скрининговым нижестоящим геном miR-133a в нашем исследовании, который, как сообщалось, участвует в ишемии / реперфузии (I / R) миокарда [18]. Кроме того, в недавнем исследовании упоминалось, что нокдаун MAML1 обладает антифиброзной функцией при фиброзе печени [19].
В свете предыдущих исследований возникает вопрос, может ли происходящая из BMSC экзосомальная miR-133a опосредовать миокардит. Таким образом, это исследование было начато с гипотезы о том, что miR-133, перемещаемая экзосомой, производной от BMSC (BMSC-Exo), улучшает фиброз миокарда и эпителиально-мезенхимальный переход (EMT) у крыс VMC за счет регуляции MAML1.
Материалы и методы
Этическое одобрение
Исследование было разрешено Комитетом по уходу и использованию животных Четвертой дочерней больницы медицинского факультета Чжэцзянского университета. С животными обращались гуманно.
Изоляция BMSC
Экспериментальные животные представляли собой взрослых крыс-самцов Sprague – Dawley (SD), не содержащих специфических патогенов (SPF) (Центр экспериментальных животных Медицинской школы Университета Чжэцзян, Чжэцзян, Китай). Крыс умерщвляли внутрибрюшинной инъекцией пентобарбитала натрия и стерилизовали 75% спиртом. Бедренную и большеберцовую кости извлекали на ультрачистый стол, мышечную и соединительную ткани удаляли, а полость костного мозга многократно промывали модифицированной средой Игла Дульбекко с низким содержанием глюкозы (DMEM). Жидкость центрифугировали для сбора осадков, которые ресуспендировали и инкубировали в течение 24 ч (среду меняли каждые 2–3 дня). При выращивании до логарифмической фазы BMSC отделяли с помощью 0,25% трипсина (Gibco, Карлсбад, Калифорния, США), центрифугировали и ресуспендировали в культуральном растворе MSC (Cyagen Biosciences Inc., Гуанчжоу, Китай). Суспензию пропускали в соотношении 1:2. Вышеупомянутую операцию повторили, и BMSC 4-го пассажа использовали для последующих экспериментов.
Идентификация BMSC
Поверхностные антигены BMSC 4-го пассажа в логарифмическом росте идентифицировали с помощью проточной цитометрии. BMSC отделяли 0,25% трипсином (1 мл), содержащим этилендиаминтетрауксусную кислоту, центрифугировали, ресуспендировали в соответствующем фосфатно-солевом буфере (PBS) и центрифугировали при 151 g. Затем BMSC ресуспендировали в PBS, содержащем 2% свежей эмбриональной телячьей сыворотки (FBS) (Gibco), чтобы получить суспензию отдельных клеток. Моноклональные антитела FITC-CD34, PE-CD29 и PE-CD44 (по 5 мкл каждое, BD Biosciences, Франклин Лейкс, Нью-Джерси, США) инкубировали с суспензией клеток (100 мкл), центрифугировали при 151 g, повторно суспендировали с 500 мкл. мкл PBS, содержащего 1% параформальдегида и фиксированного в течение 30 мин. Фоновые маркеры были идентифицированы с использованием моноклональных антител из контроля гомотипа.
Проточная цитометрия:суспензию единичных клеток фиксировали и центрифугировали при 151 g. Затем BMSC повторно суспендировали в PBS, содержащем 1% параформальдегида, тестировали на проточном цитометре MACS Quart и анализировали с помощью соответствующего программного обеспечения.
Индукция остеогенеза и адипогенеза BMSC
BMSC в 4-м пассаже высевали в 6-луночные планшеты с концентрацией 200 клеток / мл. Раствор для индукции остеобластов и раствор для индукции адипогенности (Cyagen Biosciences Inc.) добавляли в BMSC с слиянием 60–70%. BMSC в другие две лунки не добавляли с индукционными жидкостями в качестве контролей. BMSC индуцировали в течение 14 дней и фиксировали 4% параформальдегидом. Затем дифференцированные остеобласты и адипоциты были окрашены ализарином красным и масляным красным О (Wuhan Pulande Biological Technology Co., Ltd., Ухань, Китай) и исследованы под микроскопом.
Выделение и идентификация экзосом
BMSC в 4-м пассаже культивировали в течение 48 часов для сбора супернатанта, который затем центрифугировали (800 g и 2000 g), фильтровали с помощью фильтрующих мембран 0,22 мкм и 100000 MW и центрифугировали (100000 g) для сбора осадков. Затем осадки ресуспендировали в PBS, снова центрифугировали при 100,00 g для получения преципитации экзосом. Суспензию BMSC-Exo в PBS подвергали определению концентрации с помощью бицинхониновой кислоты (BCA) и детекции белка-производителя экзосом (CD63, CD81 и CD9) с помощью вестерн-блоттинга (Proteintech, Чикаго, Иллинойс, США).
Рекомбинантная аденовирусная инфекция опосредует модификацию генов miR-133a BMSC
BMSC пассировали в течение ночи. Нормальный контроль (равное количество PBS), отрицательный контроль miR-133a (NC), сверхэкспрессия miR-133a (Ad-miR-133a) и miR-133a с низкой экспрессией (Adas-miR-133a) ( Hanbio Biotechnology Co., Ltd., Шанхай, Китай) трансфицировали BMSC в соответствии со 100 множественностью инфекции (MOI). BMSC культивировали, и соответствующие экзосомы (NC Exo , Северная Каролина Exo , Ad-miR-133a Exo , и Adas-miR-133a Exo ) были получены ультрацентрифугированием [20].
Создание модели VMC на крысах и группирование экспериментальных животных
Зрелых самцов крыс SD класса SPF разделили на 10 групп по восемь крыс в каждой. Вирус Коксаки B3 (CVB3) был предоставлен Институтом медицинской биотехнологии Китайской академии медицинских наук (Пекин, Китай).
CVB3 (10 мг / кг) вводили крысам внутрибрюшинно, тогда как PBS или BMSC-Exo (100 мкг) вводили через хвостовую вену. Нормальным крысам для контроля вводили культуральный раствор CVB3 и PBS. Крысам, которым вводили 10 мг / кг CVB3, дополнительно вводили PBS, MSC exo , Северная Каролина Exo , Адас-miR-133a Exo , Ad-miR-133a Exo , si-NC или si-MAML1 (RIBOBIO, Гуанчжоу, Китай).
Крысам непрерывно вводили инъекции в течение 7 дней и получали кровь из глазного яблока. Кровь центрифугировали, сыворотку собирали, упаковывали и хранили при -20 ° C. После умерщвления крыс отбирали образцы сердца, фиксировали 10% формальдегидом, обезвоживали градиентным спиртом, очищали ксилолом и заливали парафином, делали срезы для гистологического наблюдения. Часть срезов помещали при -80 ° C как материалы для экспериментов по молекулярной биологии.
Эхокардиография
На 7-й день после введения вируса крысам внутрибрюшинно вводили пентобарбитал натрия 25 мг / кг. После полной анестезии отведение от конечности аппарата электрокардиограммы, соединенное с иглой электрода, вводилось подкожно на концах конечностей крыс и записывалась электрокардиограмма отведения от конечности. Затем крыс фиксировали в положении лежа на спине немного левее, с грудной клетки депилировали и подключали электрокардиограмму во II отведении для получения доплеровского спектра пульса кровотока аорты в парастернальном четырехкамерном отделе сердца. Показатели включали толщину задней стенки левого желудочка (LVPW), конечный систолический диаметр левого желудочка (LVID), фракцию укорочения левого желудочка (FS) и фракцию выброса левого желудочка (LVEF).
Окрашивание гематоксилином – эозином (HE)
Ткани фиксировали 4% параформальдегидом, обезвоживали, очищали и заливали парафином. Затем срезы размером 4 мкм депонировали, окрашивали гематоксилином (Servicebio, Ухань, Китай), дифференцировали 1% -ным спиртом соляной кислоты, снова окрашивали в синий цвет и окрашивали эозином, обезвоживали, очищали ксилолом, запечатывали нейтральной камедью. и наблюдали под оптическим микроскопом (Olympus, Токио, Япония).
Окрашивание коллагеном Masson
Парафиновые срезы депарафинизировали, окрашивали гематоксилином менее чем за 2 мин, окрашивали раствором пурпурного Lichun и быстро промывали 0,5% раствором ледяной уксусной кислоты. Затем срезы окрашивали 1% водным раствором фосфата алюминия, окрашивали от темно-красного до ярко-красного до розового и наблюдали под микроскопом. Затем срезы окрашивали анилиновым синим (Pulande), обычно обезвоживали ксилолом и герметизировали. Программное обеспечение для анализа медицинских изображений Image-Proplus 6.0 использовалось для измерения площади положительного окрашивания коллагеновых волокон, а объемная доля коллагена (CVF) =коллагеновая площадь / общая площадь поля. Различали место окрашивания и цвет коллагеновых волокон (кардиомиоциты были красными, а коллагеновые волокна - синими полосами или однородными структурами в межклеточном пространстве).
Конечная дезоксинуклеотидилтрансфераза-опосредованная дезоксиуридинтрифосфат-биотиновая окраска концов метки по нику (TUNEL)
Парафиновые срезы удаляли парафином, помещали в цитратный буфер и запекали при 350 Вт в течение 10 мин. В срезы добавляли 50 мкл раствора TUNEL, соединяли с 50 мкл конверсионного агента-пероксидазы, проявляли с помощью DAB и наблюдали под микроскопом. Срезы помещали в гематоксилин, погружали в 95% этанол I – II, соединяли с безводным этанолом I – II, ксилолом I – II и запаивали. Результаты были проанализированы под оптическим микроскопом.
Иммуноферментный анализ (ELISA)
Фактор некроза опухоли α (TNF-α), интерлейкин (IL) -1β и IL-6 определяли с помощью наборов для ELISA (BOSTER Biological Technology Co. Ltd., Ухань, Китай). Кровь глазного яблока центрифугировали при 604 g для сбора верхней сыворотки. Супернатант, полученный центрифугированием из среды для культивирования клеток, детектировали в клеточном эксперименте. В стандарте разведения образцов было семь градиентов концентрации. К пустой лунке добавляли разбавитель для образца, а в другую лунку добавляли тетраметилбензидин (TMB), для каждой концентрации устанавливали две дублирующие лунки. К лункам для образцов по очереди добавляли 50 мкл разбавителя для образцов и образцы. В каждую лунку вводили 100 мкл первичных антител (кроме лунки TMB) в течение 1 часа, а также 300 мкл 0,01 M трис-забуференного физиологического раствора (TBS) и 100 мкл рабочего раствора комплекса авидин-биотин-пероксидаза (кроме лунки TMB. ). Затем в каждую лунку добавляли 300 мкл 0,01 М TBS и инкубировали со 100 мкл TMB. Немедленно измеряли значение оптической плотности (OD) и концентрацию в каждой лунке и строили стандартную кривую.
Количественная полимеразная цепная реакция с обратной транскрипцией (RT-qPCR)
Экспрессия MiR-133a, коллагена Ι, коллагена III, α-SMA, TGF-β1, CTGF, E-кадгерина и FSP-1 в тканях миокарда и кардиомиоцитах определялась с помощью RT-qPCR. Общую РНК экстрагировали из кардиомиоцитов или тканей миокарда и обратно транскрибировали в кДНК с помощью набора для экстракции РНК (Takara, Dalian, China), а праймеры RT-PCR были синтезированы с помощью Invitrogen (Гуанчжоу, Китай), последовательности показаны в таблице 1. Относительную количественную экспрессию генов анализировали с использованием глицеральдегид-3-фосфатдегидрогеназы (GAPDH) или U6 в качестве генов контроля нагрузки согласно 2 - △△ Ct метод.
Вестерн-блот-анализ
Крыс умерщвляли под наркозом. Ткани миокарда замораживали и измельчали в жидком азоте. Затем исходный раствор ингибиторов протеаз фенилметансульфонилфторида смешивали с буфером для лизиса клеток в соотношении 1:100 (Beyotime Biotechnology Co., Ltd., Шанхай, Китай). Образцы лизировали смешанным раствором и экстрагировали белки клеток. Концентрация общего белка определялась с помощью набора BCA. Образцы смешивали с 5-кратным загрузочным буфером в соотношении 4:1, добавляли кипящую водяную баню в течение 10 мин, охлаждали во льду и центрифугировали. Было выполнено разделение с помощью электрофореза, и белки были перенесены на мембрану из поливинилиденфторида (Servicebio) с раствором для электрического переноса. Затем мембрану блокировали 5% сухим обезжиренным молоком и соединяли с первичными антителами CD63, CD81 и CD9 (кроличьи поликлональные антитела против крысиных антител от Proteintech, 1:100), MAML1 (ab65090, Abcam, MA, USA, 1:1000) и GAPDH (Santa Cruz Biotechnology, Inc, Санта-Крус, Калифорния, США, 1:1000). Затем на мембрану капали вторичное антитело, IgG, меченное пероксидазой хрена (Cell Signaling Technology, Беверли, Массачусетс, США, 1:1000), и погружали в реакционный раствор с усиленной хемилюминесценцией (Пирс, Рокфорд, Иллинойс, США). Используя GAPDH в качестве контроля загрузки, изображения импринтинга белков анализировали с помощью программного обеспечения ImageJ2x.
Культура и пассаж кардиомиоцитов
Были взяты крысы SD в возрасте 3–5 дней (Центр экспериментальных животных медицинского факультета Чжэцзянского университета, Чжэцзян, Китай). Часть желудочка промывали предварительно охлажденным сбалансированным солевым раствором Хэнка, разрезали на мелкие кусочки и отделяли 0,25% трипсином. К кусочкам добавляли подходящее количество 10% полной среды для прекращения отделения и центрифугировали при 151 g. DMEM, содержащую 20% FBS, применяли для повторного суспендирования клеток. Кардиомиоциты очищали методом дифференциальной адгезии, выживаемость наблюдали с помощью окрашивания трипановым синим, выжившие кардиомиоциты культивировали. Через 24 ч кардиомиоциты прилипли к стенке и начали пульсировать. Через 72 ч псевдоподии расширились.
Создание модели кардиомиоцита VMC
Кардиомиоциты в 4-м пассаже в фазе логарифмического роста были отобраны и инфицированы MSC Exo , Северная Каролина Exo , Адас-miR-133a Exo , и Ad-miR-133a Exo . Раствор вируса 100 Tcid50 CVB3 (100 мкл) добавляли к клеткам для индукции модели клеточной VMC. В то же время равное количество поддерживающего раствора было добавлено к клеткам для контроля, и соответствующие экзосомы были объединены в кардиомиоциты после 1-часового заражения для 47-часового культивирования.
Набор для подсчета клеток (CCK) -8 Assay
Набор для обнаружения клеток CCK-8 (Beyotime) применяли для определения выживаемости кардиомиоцитов. При росте до логарифмической фазы клетки отделяли 0,25% трипсином и высевали в 96-луночный планшет для культивирования клеток при 2,5 × 10 4 клеток / на лунку. В сочетании с раствором CCK-8 (10 мкл / лунка) клетки непрерывно культивировали в течение 1–4 ч, и OD 450 нм значение было измерено с помощью считывающего устройства для микропланшетов.
Проточная цитометрия
Метод двойного окрашивания аннексином V-APC / пропидиум йодидом (PI) применялся для обнаружения апоптоза клеток. Клетки центрифугировали, ресуспендировали с 250 мкл буфера для связывания (4 мл буфера для связывания + 12 мл деионизированной воды) и доводили до 1 × 10 6 . клеток / мл. К 100 мкл клеточной суспензии добавляли 5 мкл аннексина V-APC (BD Biosciences) и 5 мкл раствора PI (BD Biosciences), загружали в проточный цитометр и автоматически анализировали с помощью компьютера.
Анализ двойного люциферазного репортерного гена
Последовательность дикого типа (wt) или мутантного типа (mut) MAML1 3-нетранслируемой области (UTR) клонировали в вектор pGL3-M (Promega, WI, США), затем MAML1-3-UTR-wt или MAML1- Были созданы 3-UTR-mut. Векторы вместе с миметиком miR-133a или NC были котрансфицированы в кардиомиоциты с помощью Lipofectamine 2000. Активность люциферазы тестировали через 48 часов с помощью двойной системы репортерных генов люциферазы (Promega) [21]
Анализ иммунопреципитации РНК (RIP)
Набор RIP (Millipore, США) использовали для обнаружения связывания MAML1 и miR-133a. Клетки лизировали буфером для анализа радиоиммунопреципитации (P0013B, Beyotime, Шанхай, Китай), центрифугировали при 1400 g и инкубировали с антителами для совместного преципитации. Магнитные шарики (50 мкл) ресуспендировали в 100 мкл промывочного буфера RIP и инкубировали с 5 мкг антитела против MAML1 (1 мкг / мл, ab155786) или IgG (1:100, ab172730). Комплекс магнитных шариков и антител ресуспендировали в 900 мкл промывочного буфера RIP, взаимодействовали со 100 мкл клеточного экстракта, расщепляли протеиназой K и детектировали с помощью RT-qPCR [22].
Статистический анализ
Для анализа данных применялось статистическое программное обеспечение SPSS 21.0 (IBM Corp. Armonk, NY, USA). Данные измерений были выражены как среднее ± стандартное отклонение. Для сравнения между двумя группами применялся t-критерий. Односторонний дисперсионный анализ (ANOVA) использовался для сравнения между группами, а апостериорный тест Тьюки - для попарных сравнений. Предикторы сохранялись, если они были значимыми при P значение 0,05 или меньше.
Результаты
Идентификация BMSC и BMSC-Exo
Микроскопически BMSC были веретенообразными и округлыми и прикреплялись к стенке в виде вихря или радиального узора (рис. 1A). После окрашивания масляным красным О для индукции адипогенеза липидные капли 4-го BMSC были красными, и были круглые липидные капли разного размера (рис. 1B). После индукции остеогенеза клетки, экспрессирующие кальцифицированные узелки, показали красный цвет после окрашивания ализарином красным и неравномерное распределение кальцифицированных узелков и перекрывающихся клеток (рис. 1С). Проточная цитометрия показала, что маркеры MSC CD29 и CD44 (> 95%) были экспрессированы, но поверхностный антиген гематопоэтических стволовых клеток CD34 (<95%) не экспрессировался (фиг. 1D). Эти результаты показали, что BMSC обладают высокой чистотой и соответствуют стандартам MSC Международного общества клеточной терапии.
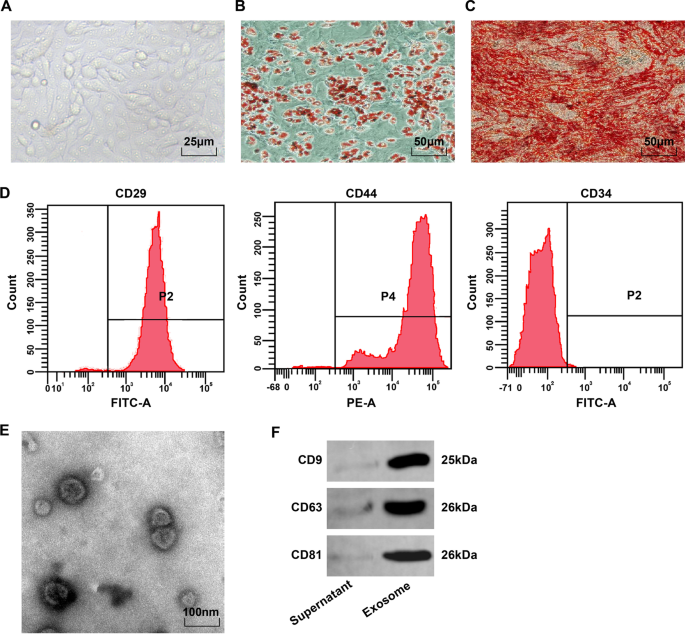
Наблюдение за фенотипом BMSC и идентификация BMSC-Exo. А Морфологическое наблюдение BMSC в 4-м пассаже; Б Результаты окрашивания адипоцитов масляным красным О; В Результаты окрашивания остеобластов ализариновым красным; D Фенотип BMSC определяется с помощью проточной цитометрии. E Наблюдение BMSC-Exo под электронным микроскопом; F Полосы белка CD9, CD63 и CD81
Просвечивающий электронный микроскоп обнаружил, что BMSC-Exos представляют собой везикулы овальной формы с четкой периферической мембранной структурой, разных размеров и диаметром 40–100 нм (рис. 1E). Вестерн-блоттинг показал, что экстрагированные продукты экспрессируют характерные белки CD9, CD63 и CD81, полученные из экзосом (рис. 1F).
Повышенный уровень экзосомальной miR-133a облегчает симптомы миокардита
Наблюдалась трансфекция BMSC рекомбинантным аденовирусом miR-133a (фиг. 2A). Большое количество зеленой флуоресценции экспрессии NC, Ad-miR-133a и Adas-miR-133a наблюдали под инвертированным флуоресцентным микроскопом, что указывает на то, что рекомбинантный аденовирусный вектор может эффективно трансфицировать BMSC. Чтобы проверить эффективность трансфекции miR-133a, экспрессию miR-133a в BMSC и их экзосомах измеряли с помощью RT-qPCR. Было обнаружено, что повышающая регуляция miR-133a увеличивает экспрессию miR-133a, тогда как понижающая регуляция miR-133a снижает экспрессию miR-133a (Рис. 2B). Впоследствии мы вводили крысам экзосомы, содержащие miR-133a. Под инвертированным флуоресцентным микроскопом наблюдали экспрессию зеленой флуоресценции у крыс VMC после обработки NC Exo , Ad-miR-133a Exo , или Adas-miR-133a Exo , что указывает на то, что рекомбинантный аденовирусный вектор инфицировал ткани миокарда крыс (фиг. 2C). Эксперимент RT-qPCR также обнаружил, что экспрессия miR-133a у крыс VMC была явно снижена; Экспрессия miR-133a была явно повышена у крыс VMC, которым вводили Ad-miR-133a Exo но снизился у крыс VMC, которым вводили Adas-miR-133a Exo (Рис. 2D). Что касается общих условий содержания крыс, было замечено, что общие условия нормальных контрольных крыс были нормальными, и характеристики VMC явно выражались у крыс VMC и крыс VMC, которым вводили Adas-miR-133a Exo , например, грубые и беспорядочные волосы, одышка и недостаточное питание. У крыс VMC, получавших MSC Exo , Северная Каролина Exo , и Ad-miR-133a Exo , эти признаки были улучшены в разной степени. Вес крыс VMC постоянно снижался через 1 день после заражения и инъекции MSC Exo , Северная Каролина Exo , или Ad-miR-133a Exo увеличился вес крыс. Вес крыс VMC, получавших Ad-miR-133a Exo был явно увеличен, и вес крыс VMC, которым вводили Adas-miR-133a Exo был явно уменьшен (рис. 2E).
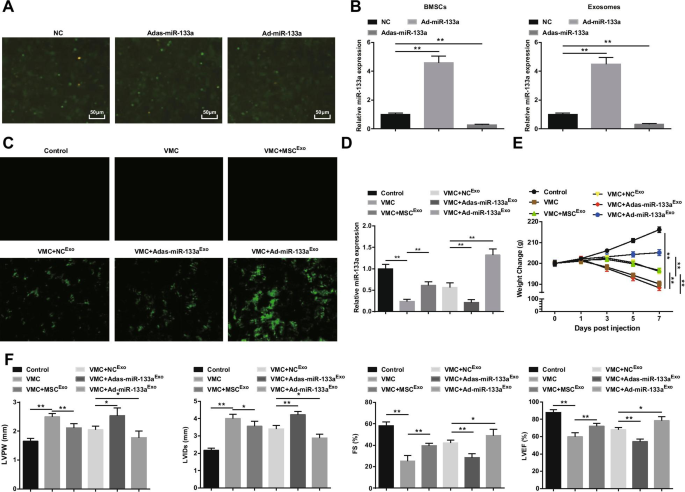
Активированная экзосомальная miR-133a облегчает миокардит. А Трансфекция BMSC рекомбинантного аденовируса miR-133a; Б RT-qPCR обнаружение экспрессии miR-133a в BMSC и их экзосомах после регуляции miR-133a; В эффективность трансфекции miR-133a проверена с помощью инвертированного флуоресцентного микроскопа; D Относительная экспрессия miR-133a в тканях миокарда, протестированная с помощью RT-qPCR; E Изменение веса крыс в каждой группе; F Определение LVPW, LVID, FS и LVEF у крыс каждой группы. * P <0,05; ** P <0,001
Наблюдение за функцией миокарда показало, что (рис. 2F) у крыс VMC были увеличены LVPW и LVID, а также уменьшены FS и LVEF. После инъекции экзосом у крыс VMC наблюдалось снижение LVPW и LVID и, очевидно, повышение FS и LVEF. Адас-miR-133a Exo лечение нарушено, пока Ad-miR-133a Exo улучшилась функция миокарда у крыс VMC.
Повышенная регуляция экзосомальной miR-133a подавляет воспаление в тканях миокарда крыс VMC
Окрашивание HE показало, что волокна миокарда у нормальных контрольных крыс были близко расположены и инфильтрации воспалительных клеток в мезенхиме не было. Кардиомиоциты у крыс VMC были дезорганизованы, и мезенхима была инфильтрирована большим количеством воспалительных клеток. Кардиомиоциты крыс VMC, которым вводили MSC Exo или NC Exo были упорядочены, с небольшим количеством воспалительных клеток, инфильтрирующих в мезенхиму. Кардиомиоциты крыс VMC после Adas-miR-133a Exo лечение было организовано беспорядочно, и воспалительные клетки в мезенхиме были инфильтрированы. Кардиомиоциты крыс VMC, получавших Ad-miR-133a Exo были организованы без явной инфильтрации воспалительных клеток (рис. 3A).
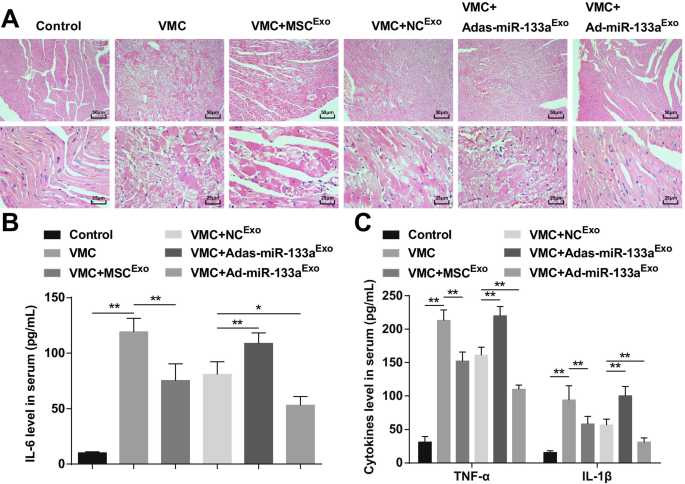
Повышенное количество экзосомальной miR-133a сдерживает воспаление в тканях миокарда с помощью VMC. А Окрашивание HE ткани миокарда крысы в каждой группе; Б Экспрессия IL-6 в сыворотке крови проверена с помощью ELISA; В Экспрессию TNF-α и IL-1β в сыворотке крови тестировали с помощью ELISA. * P <0,05; ** P <0,001
ELISA показал, что (фиг. 3B, C) воспалительные факторы (TNF-α, IL-1β и IL-6) явно увеличивались у крыс VMC. Крысам VMC вводили Ad-miR-133a Exo снизился уровень воспалительных факторов. Адас-miR-133a Exo лечение вызывало повышение воспалительных факторов у крыс VMC.
Повышенный уровень экзосомальной miR-133a снижает CVF в тканях миокарда крыс с VMC
Окрашивание по Массону показало, что волокна миокарда у нормальных крыс были расположены близко друг к другу и почти не имели синих коллагеновых волокон. После инъекции CVB3 кардиомиоциты были гипертрофированы, с гиперплазией соединительной ткани и большим количеством синих коллагеновых волокон, и CVF был явно повышен. После обработки экзосомами кардиомиоциты были упорядочены, межклеточная гиперплазия соединительной ткани уменьшилась, синие коллагеновые волокна и CVF были явно уменьшены. Межклеточное пространство миокарда крыс VMC, которым вводили Adas-miR-133a Exo был расширен, клетки были явно увеличены, синие коллагеновые волокна и CVF были явно увеличены; межклеточное пространство было уменьшено, распределение синих коллагеновых волокон и CVF уменьшилось у крыс VMC с Ad-miR-133a Exo лечение (рис. 4А, Б).
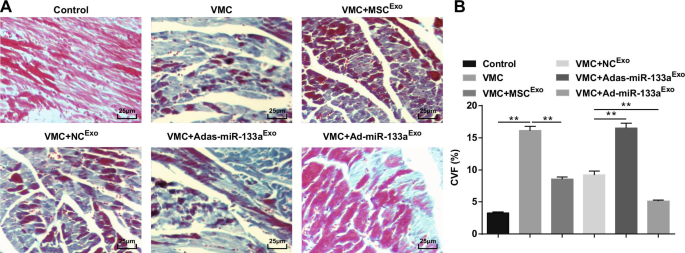
Активированная экзосомная miR-133a снижает CVF в тканях миокарда крыс с VMC. А Окрашивание тканей миокарда крыс по Массону; Б Объемная доля коллагена крыс в каждой группе. ** P <0,001
Увеличение экзосомальной miR-133a снижает экспрессию коллагена I, коллагена III, TGF-β1 и CTGF в тканях миокарда крыс с VMC
Коллаген I и коллаген III являются основными компонентами коллагена, которые в основном распределены в соединениях клеток и клеточных мембранах, межклеточном веществе и цитоплазме. TGF-β1 и CTGF являются отличительными белками фиброза. Результаты RT-qPCR продемонстрировали, что уровни экспрессии мРНК коллагена I, коллагена III, TGF-β1 и CTGF были увеличены у крыс VMC, но снизились после обработки экзосом. Крысы VMC, получавшие Ad-miR-133a Exo имели пониженные уровни экспрессии мРНК коллагена I, коллагена III, TGF-β1 и CTGF, в то время как крысы VMC после Adas-miR-133a Exo лечение показало противоположную ситуацию (рис. 5A – C).
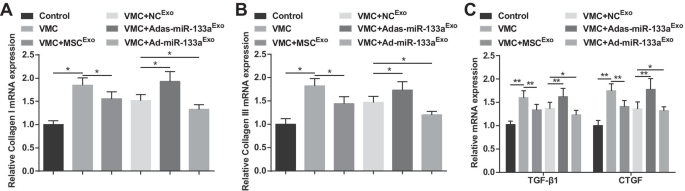
Повышенный уровень miR-133a в экзосоме снижает экспрессию мРНК коллагена I, коллагена III, TGF-β1 и CTGF в тканях миокарда крыс с VMC. А Экспрессию мРНК коллагена I в тканях миокарда крыс определяли с помощью RT-qPCR; Б Экспрессию мРНК коллагена III в тканях миокарда крыс определяли с помощью RT-qPCR; В Экспрессию мРНК TGF-β1 и CTGF в тканях миокарда крыс определяли с помощью RT-qPCR. * P <0,05; ** P <0,001
Повышенная регуляция экзосомальной miR-133a ингибирует апоптоз кардиомиоцитов в тканях миокарда крыс с VMC
Окрашивание TUNEL показало, что апоптотические кардиомиоциты были коричневато-черными или коричневато-желтыми с ядерной конденсацией. У крыс VMC увеличивалось количество апоптозных клеток, количество которых ослаблялось обработкой экзосомами. Крысам VMC вводили Ad-miR-133a Exo имели уменьшенное количество апоптотических клеток и клеток, которым вводили Adas-miR-133a Exo увеличилось количество апоптотических клеток (рис. 6A, B).
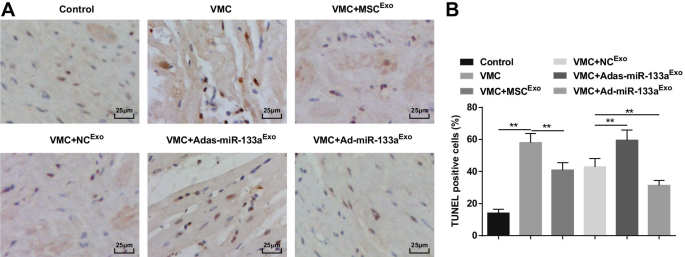
Повышенный уровень miR-133a в экзосоме подавляет апоптоз кардиомиоцитов в тканях миокарда крыс с VMC. А Окрашивание тканей миокарда крыс в каждой группе по TUNEL; Б Количество TUNEL-положительных клеток в каждой группе. ** P <0,001
Повышенная экзосомальная miR-133a подавляет ЭМП в тканях миокарда крыс с VMC
E-кадгерин, α-SMA и FSP-1 являются ключевыми индикаторами EMT. Результаты RT-qPCR продемонстрировали, что уровни экспрессии мРНК α-SMA и FSP-1 были повышены, а уровень экспрессии мРНК E-кадгерина был снижен у крыс VMC. Кроме того, уровни экспрессии мРНК α-SMA и FSP-1 были снижены, а уровень экспрессии мРНК E-кадгерина повысился у крыс VMC после обработки экзосомами. Уровни экспрессии мРНК α-SMA и FSP-1 были повышены, а уровень экспрессии мРНК E-кадгерина был снижен у крыс VMC, получавших Adas-miR-133a Exo , в то время как экспрессия этих индикаторов была противоположной у крыс VMC, которым вводили Ad-miR-133a Exo (Рис. 7A, B).
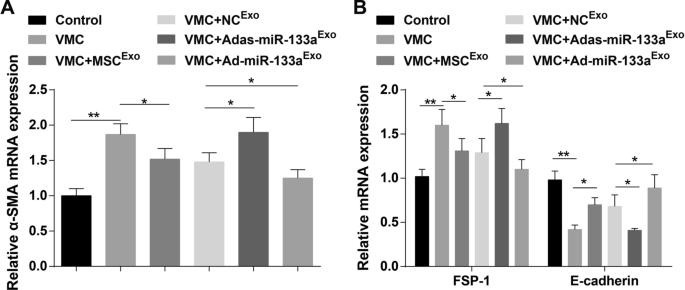
Активированная экзосомальная miR-133a репрессирует EMT в тканях миокарда крыс с VMC. А Экспрессию мРНК α-SMA в тканях миокарда крысы в каждой группе определяли с помощью RT-qPCR; Б Экспрессию мРНК FSP-1 и E-кадгерина в тканях миокарда крысы в каждой группе определяли с помощью RT-qPCR. * P <0,05; ** P <0,001
Активированная экзосомная miR-133a подавляет воспаление кардиомиоцитов в VMC
В результате флуоресцентная микроскопия зафиксировала зеленую флуоресцентную экспрессию у крыс VMC, получавших NC Exo , Ad-miR-133a Exo , или Adas-miR-133a Exo , что указывает на то, что рекомбинантный аденовирусный вектор инфицировал кардиомиоциты крыс (фиг. 8A). RT-qPCR и ELISA обнаружили, что (рис. 8B, D) экспрессия miR-133a была снижена, а воспалительные факторы (TNF-α, IL-1β и IL-6) были увеличены у крыс VMC, что было бы отменено обработкой экзосом. . Крысы VMC, получавшие Ad-miR-133a Exo имели повышенную регуляцию miR-133a и снижали воспалительные факторы у крыс VMC, в то время как у крыс, получавших Adas-miR-133a Exo представили снижение miR-133a и повышение уровня воспалительных факторов у крыс VMC.
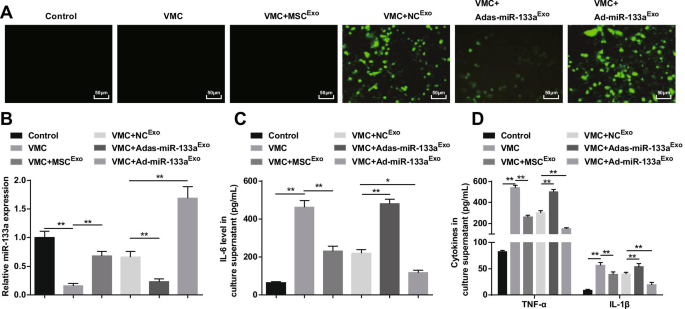
Повышенная экзосомальная miR-133a сдерживает воспаление кардиомиоцитов в VMC. А эффективность трансфекции miR-133a проверена с помощью инвертированного флуоресцентного микроскопа; Б Относительная экспрессия miR-133a в кардиомиоцитах крыс в каждой группе; В Экспрессия IL-6 в культуральном супернатанте кардиомиоцитов в каждой группе; D Экспрессия TNF-α и IL-1β в супернатанте культуры кардиомиоцитов в каждой группе. * P <0,05; ** P <0,001
Повышенное содержание экзосомальной miR-133a способствует жизнеспособности клеток и подавляет апоптоз кардиомиоцитов в VMC
Апоптоз и жизнеспособность клеток определяли с помощью двойного окрашивания AnnexinV-APC / PI и анализа CCK-8. Результаты показали, что было очевидное увеличение скорости апоптоза, снижение жизнеспособности клеток кардиомиоцитов у крыс VMC. Обработка экзосом снижает скорость апоптоза и повышает жизнеспособность кардиомиоцитов. Адас-miR-133a Exo увеличивал скорость апоптоза и нарушал жизнеспособность кардиомиоцитов у крыс VMC. Ad-miR-133a Exo лечение имело противоположный эффект на кардиомиоциты крыс VMC (рис. 9A – C).
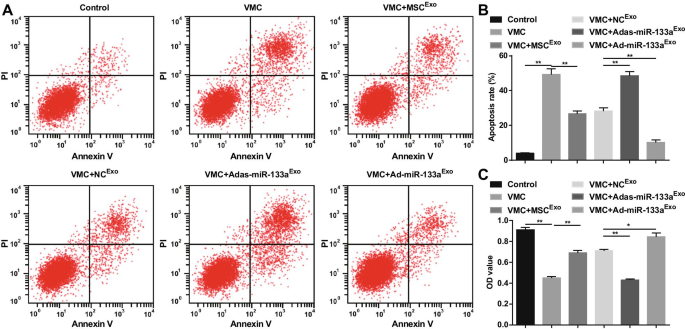
Увеличение экзосомальной miR-133a способствует жизнеспособности и подавляет апоптоз кардиомиоцитов в VMC. А Апоптоз кардиомиоцитов определяется с помощью проточной цитометрии; Б Результаты количественной оценки A; В Жизнеспособность клеток определяют с помощью анализа CCK-8. * P <0,05; ** P <0,001
miR-133a нацелен на MAML1
Сообщалось, что активированная miRNA-193b снижает повреждение I / R миокарда за счет воздействия на MAML1 [18]. На основании этого мы провели перекрестный скрининг нижестоящих генов miR-133a через биоинформатические сайты PITA, miRanda, PicTar, microT и miRmap и выбрали MAML1 в качестве мишени для miR-133a (рис. 10A). Мы сконструировали MAML1-wt или MAML1-mut и котрансфицировали кардиомиоциты с миметиком miR-133a или NC. Результаты показали, что миметик miR-133a снижает люциферазную активность MAML1-wt (фиг. 10B). Эксперимент RIP дополнительно подтвердил взаимосвязь нацеливания между miR-133a и MAML1 (фиг. 10C). Обнаружение экспрессии MAML1 с помощью RT-qPCR и вестерн-блоттинга показало, что экспрессия MAML1 была снижена в кардиомиоцитах, трансфицированных миметиком miR-133a (рис. 10D, E).
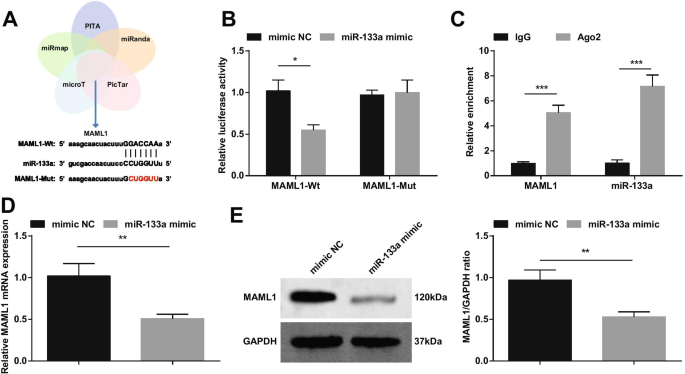
miR-133a нацелена на MAML1. А мишени miR-133a, предсказываемые на сайтах биоинформатики; Б Отношения нацеливания между miR-133a и MAML1 подтверждены экспериментом с двойным люциферазным репортерным геном; В Отношения нацеливания между miR-133a и MAML1 подтверждены экспериментом RIP; D / E Экспрессия MAML1 изменяется после активации miR-133a, обнаруживаемой с помощью RT-qPCR и вестерн-блоттинга. * P <0,05; ** P <0,01; *** P <0,001
Ингибирование MAML1 оказывает защитное действие на крыс с миокардитом и обращает действие экзосом, ингибированных miR-133a, на крыс с VMC
Для дальнейшего изучения эффекта регулируемого miR-133a MAML1 на крысах с VMC мы вводили аденовирус si-MAML1 или si-NC крысам VMC или крысам VMC, которых лечили экзосомами с подавлением miR-133a. Успех инъекции подтверждали с помощью RT-qPCR и вестерн-блоттинга (фиг. 11A, B). Результаты показали, что инъекция si-MAML1, вес крыс VMC увеличился (фиг. 11C), сердечная функция улучшилась (фиг. 11D-G), патология ткани миокарда и фиброз уменьшились (фиг. 12A-C), воспаление сыворотки (фиг. 12D, E) и апоптоз кардиомиоцитов (фиг. 13A – G) подавлялись. Кроме того, пагубные эффекты экзосом с молчанием miR-133a у крыс VMC были обращены вспять после инъекции si-MAML1.
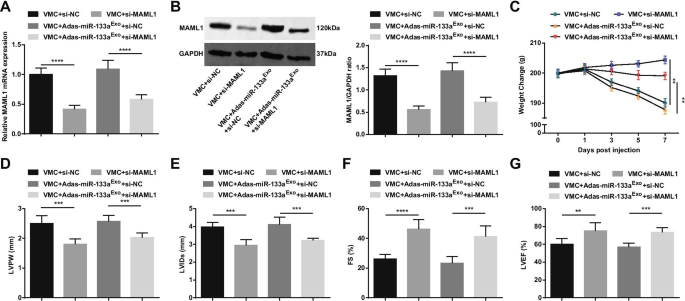
Ингибирование MAML1 оказывает защитное действие на крыс с миокардитом и может обратить действие экзосом, заглушающих miR-133a, на крыс с VMC. А / B Экспрессия MAML1 в ткани миокарда крыс, обнаруженная с помощью RT-qPCR и вестерн-блоттинга; C. Изменение веса крыс. D - G Определение LVPW, LVID, FS и LVEF у крыс; ** P <0,01; *** P <0,001; **** P <0,0001
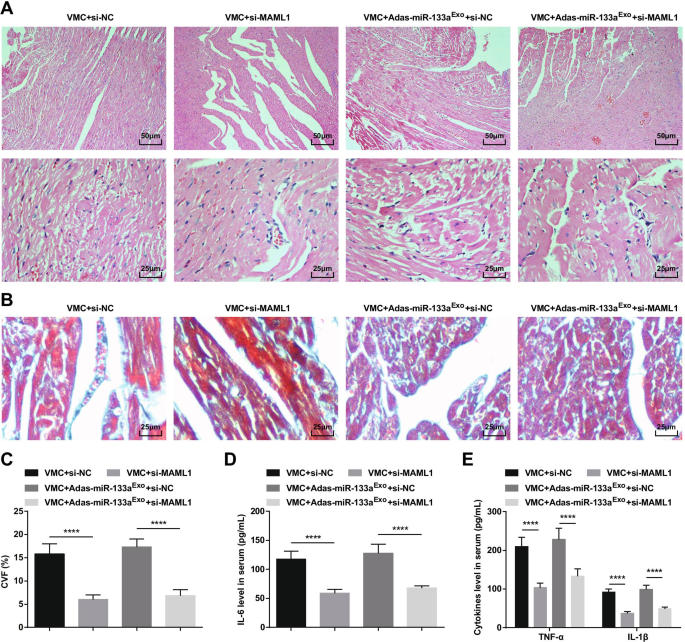
Ингибирование MAML1 может обратить действие экзосом с молчанием miR-133a на крыс с VMC. А Окрашивание ткани миокарда крыс HE; Б Окрашивание тканей миокарда крыс по Массону; В CVF крыс; D Экспрессия IL-6 в сыворотке крови проверена с помощью ELISA; E Экспрессия TNF-α и IL-1β в сыворотке крови проверена с помощью ELISA. **** P <0,0001
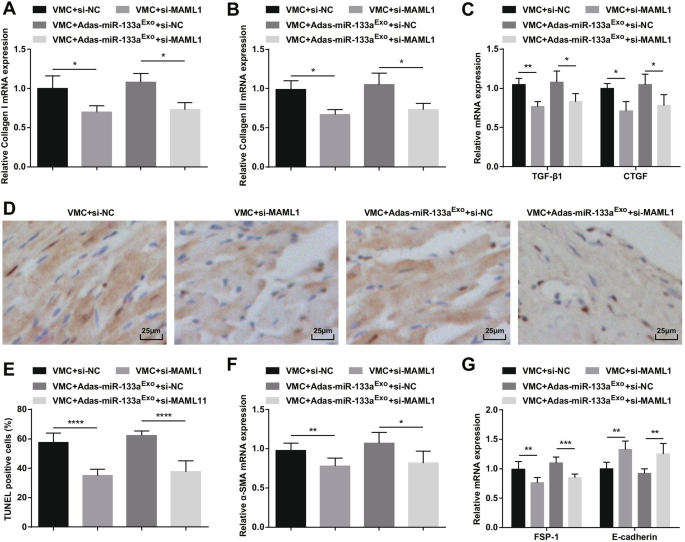
Ингибирование MAML1 может обратить действие экзосом, ингибирующих miR-133a, на крыс с VMC. А Экспрессию мРНК коллагена I в тканях миокарда крыс определяли с помощью RT-qPCR; Б Экспрессию мРНК коллагена III в тканях миокарда крыс определяли с помощью RT-qPCR; В Экспрессию мРНК TGF-β1 и CTGF в тканях миокарда крыс определяли с помощью RT-qPCR; D Окрашивание тканей миокарда крыс в каждой группе по TUNEL; E Количество TUNEL-положительных клеток в каждой группе. F Экспрессию мРНК α-SMA в тканях миокарда крысы в каждой группе определяли с помощью RT-qPCR; G Экспрессию мРНК FSP-1 и E-кадгерина в тканях миокарда крысы в каждой группе определяли с помощью RT-qPCR. * P <0,05; ** P <0,01; **** P <0,0001
Обсуждение
Миокардит - воспалительное заболевание сердца, приводящее к ДКМП и сердечной недостаточности, и чаще всего вызывается вирусными инфекциями, такими как CVB3 [2]. Исследование показало, что miR-133 снимает апоптоз кардиомиоцитов и электрическое ремоделирование у мышей с VMC [23]. Кроме того, обнаружено, что измененные экзосомальные miRNAs связаны с патогенезом CVB3-индуцированного миокардита [14]. Экзосомы, полученные из кардиальных клеток-предшественников, облегчают апоптоз, индуцированный CVB3, посредством сдерживания пролиферации CVB3 в VMC [24]. В этом исследовании изучался регуляторный механизм экзосомальной miR-133, производной от BMSC, на фиброз миокарда и EMT у крыс VMC (дополнительный файл 1:рис. 1).
Исследование показало, что экспрессия miR-133a была снижена в VMC. Как было показано ранее, экспрессия miR-133a снижена при MI [7]. Исследование также показало, что относительная экспрессия miR-133 в сердцах мышей VMC, очевидно, снижена по сравнению с контролем [23]. Есть некоторые связи миРНК с экзосомами. Дифференциальная экспрессия экзосом и экзосомальных miRNAs при заболевании рассматривается как биомаркеры заболевания с выполнением неинвазивной клинической диагностики вместе с их терапевтическим потенциалом [25]. Лин и др. . обнаружили, что miR-133 специально сортируется в экзосомы, вызванные гипоксией / реоксигенацией (H / R), происходящие из эндотелиальных клеток-предшественников человека, для увеличения ангиогенеза фибробластов и EMT [26]. Другое исследование показало, что МСК взаимодействуют с паренхимными клетками головного мозга и могут модулировать рост нейритов путем передачи miR-133b нервным клеткам через экзосомы [27].
Главный результат этой работы показал, что активированная экзосомная miR-133a способствует жизнеспособности клеток, ингибирует воспаление, апоптоз, EMT и фиброз у крыс с VMC. Они хорошо согласуются с предыдущим исследованием того, что тишина miR-133a меняет местами полисахариды астрагала ингибирование пролиферации клеток остеосаркомы MG63, вызванное лечением, вместе со стимулированием апоптоза клеток [28]. Другое исследование показало, что сверхэкспрессия miR-133a подавляет ангиогенез, апоптоз, фиброз и воспаление, одновременно ускоряя терапевтическое ремоделирование сердца при ишемических заболеваниях миокарда [29]. Подобно нашему исследованию Ли и др. . заявили, что miR-133 ингибирует апоптоз кардиомиоцитов, регулируя экспрессию связанных с апоптозом генов в сердцах мышей VMC [23]. Сверхэкспрессия miR-133a подавляет апоптоз, вызванный гипоксией, и увеличивает выживаемость кардиомиоцитов [30]. Между тем, активированные сывороточные экзосомальные miR-30a и miR-181d могут иметь потенциал для использования в качестве биомаркеров для диагностики VMC [14].
Другое открытие в нашем исследовании заключалось в том, что активированная экзосомная miR-133a снижает CVF, снижает экспрессию коллагена I и коллагена III у крыс с VMC. В статье выяснено, что высвобождение фактора роста фибробластов-18 из коллагеновой мембраны вызывает активность остеобластов, участвующих в подавлении miR-133a [31]. Избыточная экспрессия miR-133a in vitro подавляет гипертрофию кардиомиоцитов и снижает экспрессию коллагена [32], как показано в другом исследовании. CVF равняется отношению площади коллагена к сумме площади миокарда и площади коллагена, а среднее значение показывает CVF секции [33]. Об этом открытии также сообщает Ван и др. . Эта модель мышей VMC успешно сконструирована с помощью инфекции CVB3, демонстрируя очевидную более высокую экспрессию CVF по сравнению с контрольной группой [34]. Более того, результат согласуется с выводом Феррейры и др. . который демонстрирует, что miR-133a может играть важную роль в модуляции экспрессии генов в патогенезе кардиомиопатии при хронической болезни Шагаса, с потенциальной связью в качестве диагностических и прогностических инструментов [8]. Более того, данные показали, что отключение MAML1 может уменьшить гипертрофию предварительно обработанных кардиомиоцитов [35]. В нашем исследовании мы обнаружили, что MAML1 был геном-мишенью для miR-133a, а ингибирование MAML1 обращало действие экзосом, заглушающих miR-133a, на крыс с VMC. При ишемии-реперфузионном повреждении миокарда miR-193b-опосредованное подавление MAML1 может частично снижать инфаркт и ферменты миокарда, а также ослаблять апоптоз кардиомиоцитов [18]. Кроме того, есть отчет, предполагающий, что дефицит MAML1 может уменьшить фиброгенез печени [19].
Заключение
В заключение, настоящее исследование предлагает доказательства того, что miR-133a подавляется у крыс с VMC, а повышенный уровень экзосомального miR-133a улучшает сердечную функцию и сдерживает фиброз миокарда и EMT у крыс с VMC, а также увеличивает жизнеспособность и подавляет апоптоз кардиомиоциты в VMC посредством нацеливания на MAML1. Наше исследование также предполагает, что ингибирование MAML1 оказывает защитное действие на крыс с миокардитом и обращает действие экзосом, ингибированных miR-133a, на крыс с VMC. Идентификация экзосомальной miR-133a при фиброзе миокарда и EMT миокардита может потенциально расширить наше понимание механизмов, лежащих в основе миокардита, а также иметь клиническую ценность в качестве новой молекулярной мишени. Необходимо провести дополнительные исследования, чтобы найти способы лечения этого заболевания.
Сокращения
- miR-133:
-
МикроРНК-133
- BMSC-Exo:
-
Экзосома костного мозга, полученная из мезенхимальных стволовых клеток
- EMT:
-
Эпителиально-мезенхимальный переход
- VMC:
-
Вирусный миокардит
- CVF:
-
Объемная доля коллагена
- DCM:
-
Дилатационная кардиомиопатия
- CVB3:
-
Вирус Коксаки B3
- miRNA:
-
МикроРНК
- MI:
-
Инфаркт миокарда
- SPF:
-
Отсутствие специфических патогенов
- SD:
-
Спраг – Доули
- DMEM:
-
Среда Игла, модифицированная Дульбекко
- PBS:
-
Физиологический раствор с фосфатным буфером
- FBS:
-
Фетальная бычья сыворотка
- NC:
-
Отрицательный контроль
- MOI:
-
Множественность заражения
- LVPW:
-
Толщина задней стенки левого желудочка
- LVID:
-
Конечный систолический диаметр левого желудочка
- LVEF:
-
Фракция выброса левого желудочка
- HE:
-
Гематоксилин – эозин
- ТУНЕЛЬ:
-
Терминальная дезоксинуклеотидилтрансфераза, опосредованная дезоксиуридинтрифосфат-биотином, ник-конец-метка
- ELISA:
-
Иммуноферментный анализ
- TMB:
-
Тетраметилбензидин
- TBS:
-
Трис-буферный раствор
- ABC:
-
Комплекс авидин-биотин-пероксидаза
- OD:
-
Оптическая плотность
- RT-qPCR:
-
Количественная полимеразная цепная реакция с обратной транскрипцией
- GAPDH:
-
Глицеральдегид-3-фосфатдегидрогеназа
- CTGF:
-
Фактор роста соединительной ткани
- CCK:
-
Набор для подсчета клеток
- PI:
-
Иодид пропидия
- ANOVA:
-
Дисперсионный анализ
Наноматериалы
- Наночастицы черного фосфора способствуют остеогенной дифференцировке EMSC посредством усиленной экспрессии …
- Структура и электронные свойства наноглины каолинита, легированной переходным металлом
- Волокна из активированного угля, «густо заросшие» от Ag Nanohair в результате самосборки и быстрого термического …
- Изучение силы адгезии и стеклования тонких пленок полистирола с помощью атомно-силовой микроскопии
- Исследования TEM и STEM морфологии поперечного сечения двух- / трехслойных широкополосных антиотражающих пленок…
- Сравнительное исследование биосинтезированных наночастиц оксида меди и цинка in vivo путем внутрибрюшинного и…
- Разработка и характеристика Sr-содержащих стеклокерамических композитов на основе биогенного гидроксиапати…
- Материалы семейства графена в регенерации костной ткани:перспективы и проблемы
- Наноскопия хондрогенно дифференцирующихся стволовых клеток человека, полученных из жировой ткани:нанострук…
- Меченные наночастицами берлинского голубого мезенхимальные стволовые клетки:оценка жизнеспособности, прол…