


Восстановленная микроРНК-326-5p ингибирует апоптоз нейронов и ослабляет повреждение митохондрий за счет подавления STAT3 при церебральной ишемии / реперфузионной травме
Аннотация
Исследования в значительной степени изучали роль микроРНК (миРНК) в ишемии головного мозга / реперфузионном повреждении (CI / RI). Но специфический механизм miR-326-5p в CI / RI все еще неуловим. Следовательно, это исследование должно было раскрыть механизм miR-326-5p / сигнального преобразователя и активатора оси транскрипции-3 (STAT3) в CI / RI. Две модели (кислородная и глюкозная депривация [OGD] в первичных кортикальных нейронах крыс и окклюзия средней мозговой артерии [MCAO] у крыс Sprague-Dawley) были созданы для имитации CI / RI in vitro и in vivo, соответственно. Анализы потери и увеличения функции проводили с нейронами, обработанными OGD, и с крысами MCAO. После этого были протестированы жизнеспособность, апоптоз, окислительный стресс и потенциал митохондриальной мембраны в нейронах, обработанных OGD, а также патологические изменения, апоптоз и потенциал митохондриальной мембраны в тканях мозга крыс MCAO. Выявлена экспрессия митофузина-2 (Mfn2), miR-326-5p и STAT3 в нейронах, обработанных OGD, и в тканях мозга крыс MCAO. Mfn2 и miR-326-5p были снижены, а STAT3 был повышен в обработанных OGD нейронах и тканях мозга крыс MCAO. miR-326-5p нацелена на экспрессию STAT3 и негативно регулируется. Восстановление miR-326-5p или снижение STAT3 усиливает жизнеспособность, ингибирует апоптоз и окислительный стресс, увеличивает потенциал митохондриальной мембраны и увеличивает экспрессию Mfn2 в нейронах, обработанных OGD. Повышающая регуляция miR-326-5p или понижающая регуляция STAT3 ослабляли патологические изменения, подавляли апоптоз и повышали потенциал митохондриальной мембраны и экспрессию Mfn2 в тканях мозга крыс с MCAO. Это исследование выясняет, что повышенная регуляция miR-326-5p или пониженная регуляция STAT3 защищает от CI / RI за счет повышения экспрессии Mfn2.
Введение
Церебральная ишемия / реперфузионное повреждение (CI / RI) - это вид повреждения головного мозга, за которым следует ишемический инсульт [1]. Наиболее характерным признаком церебрального I / R является начальная преходящая церебральная ишемия после реперфузии [2]. CI / RI активирует апоптоз нейронов и приводит к повреждению гиппокампа и коры головного мозга [3]. В настоящее время наиболее распространенным клиническим применением является тромболитик для CI / RI [4]. Однако реперфузия может увеличить продукцию активных форм кислорода (АФК), что приведет к повреждению внутриклеточной ДНК, повреждению, связанному с окислительным стрессом, окислению белков и перекисному окислению липидов, тем самым еще больше ухудшив гематоэнцефалический барьер и отек [5]. Таким образом, срочность поиска новых целевых опций превышает приоритет CI / RI.
МикроРНК (miRNAs) широко рассматриваются как потенциальные и прогностические роли при CI / RI [6, 7]. Например, в статье показано, что miR-202-5p ослабляет нейрональное повреждение и неврологический дефицит, а также вызванное кислородной и глюкозной депривацией (OGD) клеточное повреждение у крыс с моделью окклюзии средней мозговой артерии (MCAO) при ишемическом повреждении [8] . Кроме того, обнаружено, что повышающая регуляция miR-98 улучшает неврологические исходы у мышей с I / R инсультом [9], а избыточная экспрессия miR-451 облегчает ишемический церебральный апоптоз у мышей с CI / RI [10]. В частности, miR-326-5p обладает проангиогенной способностью к эндотелиальным клеткам-предшественникам и может улучшать сердечную функцию после острого инфаркта миокарда [11]. Однако исследование miR-326-5p в CI / RI все еще находится в зачаточном состоянии. Сигнальные преобразователи и активатор транскрипции (STAT) представляют собой уникальное семейство белков, которые активируются CI / RI [12]. STAT3 был предсказан как мишень для miR-326-5p на сайте биоинформатики; Таким образом, мы изучили роль miR-326-5p-опосредованного STAT3 в CI / RI. STAT3 ухудшает ишемические нейровоспалительные процессы и вторичное повреждение головного мозга за счет высвобождения провоспалительных медиаторов [13, 14]. В частности, путь Janus-активированной киназы 2 (JAK2) / STAT3 играет функциональную роль в блокировании апоптоза клеток, индуцированного CI / RI [15]. Митофузин-2 (Mfn2) представляет собой фактор слияния митохондрий, который может защищать от сердечно-цереброваскулярного I / RI [16] и улучшать индуцированный гипоксией апоптоз нейронов при ишемическом повреждении головного мозга [17]. Также было зарегистрировано, что Mfn2 оказывает защитное действие на CI / RI [18].
Одним словом, менее глубокое исследование обнаружило комбинированную роль miR-326-5p и STAT3 в CI / RI. Учитывая это, данное исследование запускается с гипотезой о том, что miR-326-5p ослабляет CI / RI посредством нацеливания на STAT3.
Материалы и методы
Этика
Эксперименты были одобрены Комитетом по уходу за животными и их использованию дочерней больницы Университета Янчжоу; Университет Янчжоу, и он был проведен с Руководством по уходу и использованию экспериментальных крыс дочерней больницы Университета Янчжоу; Университет Янчжоу.
Экспериментальные животные
Взрослые самцы крыс Sprague-Dawley (возраст 6-8 недель, 250 ± 30 г) из Центра сравнительной медицины Университета Янчжоу (Янчжоу, Китай) содержались в специальной среде, свободной от патогенов, со свободным доступом к пище и воде. (24 ± 1) ° C, (50 ± 5)% влажности и 12-часовой цикл свет / темнота.
Изоляция и идентификация ячеек
Крыс умерщвляли обезглавливанием, а их мозг разрезали на 1 мм 3 для конфигурирования клеточных суспензий, которые были смешаны с 0,4% раствором трипанового синего в соотношении 9:1 (конечная концентрация трипанового синего составляла 0,04%), а плотность и выживаемость клеток рассчитывались с помощью гемоцитометра. Ячейки (1 × 10 6 клеток / мл) высевали в колбу для культивирования клеток, предварительно покрытую L-полилизином (0,1 мг / мл) в концентрации 5 × 10 6 клеток в колбе. Когда клетки прилипли к стенкам, их инкубировали с обновленной средой (NeurobasalA + B27 + L-глутамин) и непрерывно культивировали в течение 6–7 дней, меняя среду наполовину каждые 2–3 дня.
Идентификация клеток:нейроны коры головного мозга крысы использовали для приготовления слайдов нейронов, которые инкубировали с первичным кроличьим поликлональным антителом против нестина крысы (NES) (1:200), а также с меченным флуоресцеином изотиоцианатом (FITC) козьим антителом против кроличьего иммуноглобулина. G (1:50, оба от Cell Signaling Technology, Беверли, Массачусетс, США). После этого клетки закрывали антифлуоресцентным гасителем и наблюдали с помощью флуоресцентной микроскопии.
Создание модели OGD
Нейроны культивировали в растворе Эрла без глюкозы и подвергали воздействию газовой смеси 95% N 2 и 5% CO 2 . После 30 мин вентиляции с положительным давлением образовывалась гипоксическая среда, в которой клетки инкубировались (моделирование ишемии с помощью OGD in vitro). После 90 мин инкубации клетки инкубировали с поддерживающей клеточной средой после удаления раствора Эрла, не содержащего глюкозу. Клетки обычно культивировали для последующих экспериментов (моделирование реперфузии путем реперфузии кислород-глюкоза in vitro). Нормальная среда при нормоксии служила контролем.
Нейроны были разделены на контрольную группу; группа OGD, группа отрицательного контроля (NC) (нейроны, обработанные OGD, трансфицированные скремблированными олигонуклеотидами), группа агомира miR-326-5p (нейроны, обработанные OGD, трансфицированные агомиром miR-326-5p), группа sh-STAT3 (Обработанные OGD нейроны, трансфицированные агомиром STAT3) и группа miR-326-5p агомир + сверхэкспрессия (oe) -STAT3 (обработанные OGD нейроны, трансфицированные агомиром miR-326-5p и вектором pcDNA-STAT3). Олигонуклеотиды и векторы были предоставлены компанией GenePharma (Шанхай, Китай).
Используя липофектамин 2000 (Invitrogen; Thermo Fisher Scientific), скремблированные олигонуклеотиды, агомир miR-326-5p, shRNA STAT3 или агомир miR-326-5p и pcDNA-STAT3 трансфицировали в первичные нейроны крысы, и эффективность трансфекции проверяли с помощью количественной обратной транскрипции. полимеразная цепная реакция (RT-qPCR) или вестерн-блоттинг через 48 часов.
Анализ 3- (4, 5-диметилтиазол-2-ил) -2,5-дифенилтетразолийбромида (МТТ)
Нейроны были доведены до суспензии 1 × 10 6 клеток / мл и культивировали в течение 0, 24 и 48 ч соответственно. В стерильной среде нейроны инкубировали в течение 2–4 ч в 10% растворе МТТ и добавляли 150 мкл растворенного раствора форманзана (диметилсульфоксида) до тех пор, пока кристалл не растворился в достаточной степени. Значение оптической плотности (OD) измеряли на считывающем устройстве для микропланшетов при 570 нм.
Анализ апоптоза проточной цитометрией
Нейроны отделяли с 0,25% трипсином, центрифугировали при 2000 об / мин и ресуспендировали в PBS. Затем нейроны центрифугировали при 2000 об / мин, осадок ресуспендировали связывающим буфером и инкубировали последовательно с 5 мкл аннексина V-FITC и 5 мкл йодида пропидия (PI). Для определения скорости апоптоза применялся проточный цитометр.
Обнаружение потенциала митохондриальной мембраны JC-1
Клетки в 6-луночном планшете промывали PBS, добавляли 1 мл среды для культивирования клеток и 1 мл рабочего раствора для окрашивания JC-1 (50 мкл JC-1, 8 мл сверхчистой воды и 2 мл буфера для окрашивания JC-1) (SolarbioScience , Пекин, Китай) и инкубировали 15 мин. Буфер для окрашивания 1 × JC-1 был сконфигурирован из 4 мл дистиллированной воды и 1 мл буфера для окрашивания JC-1. После инкубации клетки обрабатывали трипсином, ресуспендировали в буфере JC-1 и тестировали на проточном цитометре. Была рассчитана относительная доля зеленой флуоресценции.
Обнаружение окислительного стресса
Уровни внутриклеточных ROS измеряли с помощью набора для обнаружения 2,7-дихлорфлуоресцеина диацетата (DCF-DA) (Abcam). Короче говоря, нейроны отделяли с 0,25% трипсином, ресуспендировали и инкубировали с 10 мкМ DCF-DA в течение 30 мин. После этого флуоресценцию DCF с возбуждением / испусканием света при 495 нм / 529 нм измеряли с помощью флуоресцентной спектроскопии (BD Biosciences).
Обнаружение внутриклеточной малондиалхехиохимии (MDA), содержания глутатиона (GSH) и активности супероксиддисмутазы (SOD) определяли с помощью химической колориметрии с использованием коммерческих наборов для анализа (Beyotime Biotechnology Co., Шанхай, Китай). Значение OD было измерено с помощью ридера для микропланшетов.
Создание модели крыс
Крыс анестезировали внутрибрюшинной инъекцией пентобарбитала натрия (50 мкг / кг). На шее сделали срединный разрез, перевязали наружную сонную артерию и заблокировали общую сонную артерию артериальным зажимом. Нить из мононити нейлона с силиконовым покрытием 3–0 осторожно вводили во внутреннюю сонную артерию до легкого сопротивления. Через 90 минут преходящей церебральной ишемии шов сняли, и реперфузию проводили еще 24 часа. Имитационная группа получала такое же лечение, за исключением того, что нить не входила во внутреннюю сонную артерию. Во время операции использовалась термостатическая кровать для поддержания ректальной температуры на уровне 37 ± 0,5 ° C [19].
Крысы были разделены на 6 групп (n =6) и инъецированы плазмидой или miR в боковой церебровентрикулярный канал до создания моделей MCAO, включая фиктивную группу, группу MCAO (моделируемые крысы), группу NC (моделируемые крысы с боковой церебровентрикулярной инъекцией). 5 мкл скремблированных олигонуклеотидов [100 мкМ]), группа агомира miR-326-5p (моделированные крысы с боковой церебровентрикулярной инъекцией 5 мкл агомира miR-326-5p [100 мкМ]), группа sh-STAT3 (моделированные крысы с боковая церебровентрикулярная инъекция 5 мкл интерференционного вектора STAT3 [100 мкМ]) и группа miR-326-5p агомир + oe-STAT3 (моделированные крысы с боковой церебровентрикулярной инъекцией 5 мкл агомира miR-326-5p [100 мкМ] и 5 мкл Вектор сверхэкспрессии STAT3 [100 мкМ]) [20, 21].
Коллекция образцов
Крыс умерщвляли под наркозом, сердца перфузировали и получали весь мозг. Поврежденные ткани головного мозга разрезали на 1 × 1 × 1 мм 3 тканевые массы для подготовки парафиновых срезов, RT-qPCR, вестерн-блоттинга и определения потенциала митохондриальной мембраны.
Обнаружение потенциала митохондриальной мембраны в коре головного мозга
Кора головного мозга крыс SD была отделена и разрезана на 1 мм 3 . Блоки ткани помещали на нейлоновую сетку 300 меш и добавляли PBS. Затем суспензию клеток центрифугировали при 1000 об / мин, добавляли 70% этанол и снова центрифугировали при 1000 об / мин. После этого образец ресуспендировали в 1 мл PBS, и суспензия (1 × 10 6 клеток, 100 мкл) реагировали с раствором красителя родамин123 (10 мкл, 5 мкг / мл) и анализировали на проточном цитометре при 488 нм.
Окрашивание HE
Ткань головного мозга крысы погружали в 4% параформальдегид на 24 часа, готовили парафиновые срезы, разрезали на 4 мкм, намазывали водой при 40 ° C и запекали при 60 ° C. Обычно депарафинированные и гидратированные парафиновые срезы окрашивали окрашивающим раствором НЕ. После окрашивания HE поврежденные нейроны показали сокращение ядра, клеточный отек, вакуолизацию и потемнение ядер. Для получения изображения использовали световой микроскоп (Nikon, Япония) и количество выживших нейронов (мм 2 ) ишемической коры.
Окрашивание ТУНЕЛЯ
Парафиновые срезы депарафинизировали и обезвоживали 100%, 95%, 80% и 70% спиртом. Затем срезы погружали в 4% параформальдегид с последующей инкубацией с 0,1% натрийцитратным буфером Triton X-100 в течение 20 мин и реакционной смесью TUNEL в течение 1–1,5 ч. Затем срезы проявляли раствором пероксидазы и диаминобензидином (DAB), контрастировали гематоксилином, дегидратировали градиентным спиртом, проницаемо и герметизировали нейтральной смолой. Срезы наблюдали под световым микроскопом для подсчета TUNEL-положительных клеток. Коричневато-желтые частицы в ядре были определены как TUNEL-положительные клетки (апоптотические клетки).
Иммуногистохимия
Срезы депарафинизировали, гидратировали и извлекали цитратным антигеном, а эндогенная каталаза блокировалась 3% H 2 О 2 . В каждый срез добавляли козью сыворотку (50 мкл), инкубировали с первичным антителом (50 мкл) и подвергали реакции со вторичным антителом (50 мкл) и стрептавидином, меченным пероксидазой хрена (50 мкл). Срезы были проявлены DAB и окрашены гематоксилином, после чего следовала градиентная дегидратация спирта, проницаемость ксилола и герметизация нейтральной смолы. Срезы наблюдали под микроскопом (цитоплазма или ядро ишемизированных тканей головного мозга представляли собой желтые или коричневато-желтые частицы). Значения OD были измерены и усреднены.
RT-qPCR
Экстракцию тотальной РНК из тканей и клеток проводили Trizol (Invitrogen). Экспрессию мРНК анализировали посредством кПЦР с использованием смеси SYBR Green PCR Mix (Applied Biosystems, Калифорния, США) и системы ПЦР в реальном времени QuantStudio TM 6Flex (Applied Biosystems). Для анализа уровней миРНК были разработаны специфические праймеры обратной транскрипции «стебель-петля» и прямые / обратные праймеры (BioTNT, Шанхай, Китай). В таблице 1 перечислены все праймеры. 2 - △△ CT был принят метод расчета уровней миРНК или мРНК.
Вестерн-блот-анализ
Образцы белка разделяли электрофорезом в 10% -ном полиакриламидном геле с додецилсульфатом натрия и наносили электроблоттингом на поливинилиденфторидную (PVDF) мембрану. После этого PVDF-мембрану блокировали 5% сухим обезжиренным молоком в трис-содержащем физиологическом растворе, зондировали первичными антителами и повторно зондировали вторичным антителом в течение 2 часов. Затем полосы белка анализировали с помощью буфера щелочной фосфатазы, содержащего хлорид тетразолия Nitroblue и 5-бром-4-хлор-3-индолилфосфат, и количественно оценивали с помощью программного обеспечения ImageJ (NIH, Bethesda, MD, США). Основными антителами были STAT3 (1:1000), Mfn2 (1:1000, Abcam, MA, США) и глицеральдегид-3-фосфатдегидрогеназа (GAPDH) (1:1000, Cell Signaling Technology, Беверли, Массачусетс, США). / P>
Анализ двойного люциферазного репортерного гена
STAT3 3'UTR, содержащий сайты связывания miR-326-5p дикого типа (WT) или мутантные (Mut), клонировали в вектор pmirGLO (Promega, WI, США), названный STAT3-WT и STAT3-Mut. Клетки высевали в 24-луночный планшет в количестве 2 × 10 4 . клеток / лунку и котрансфицировали 100 нг вектора люциферазы STAT3-WT или STAT3-Mut и 50 нМ miR-326-5p агомиром или его агомиром NC при 70% конфлюэнтности. Люциферазную активность измеряли с использованием системы анализа репортеров двойной люциферазы (Promega Corporation, WI, США).
Статистический анализ
Для анализа данных использовалось статистическое программное обеспечение SPSS 21.0 (IBM, Нью-Йорк, США). Данные были выражены как среднее значение ± стандартное отклонение (SD). Разницу между двумя группами анализировали с помощью t-критерия Стьюдента, в то время как множественные сравнения анализировали с помощью одностороннего дисперсионного анализа (ANOVA) с последующим апостериорным критерием Тьюки. P представляет собой двусторонний тест, а P <0,05 считалось статистически значимым.
Результаты
Повышающая регуляция miR-326-5p или понижающая регуляция STAT3 ослабляет апоптоз корковых нейронов, обработанных OGD
При культивировании в течение недели нейроны коры головного мозга крысы дифференцировались и созревали. Микроскопически нейроны показали более крупные тела нейронов, более прозрачную цитоплазму, большое ядро с низкой плотностью и сильным показателем преломления. Тела нейронов имели форму конуса или округлой формы, а дендриты располагались в шахматном порядке и прилегали к стенкам (рис. 1а, б). NES был специфическим химическим маркером нейронов головного мозга для обнаружения нейронов с высокой специфичностью с помощью иммунофлуоресцентного анализа. Было обнаружено, что нейроны специфически связываются с NES и демонстрируют зеленую флуоресценцию (рис. 1c).
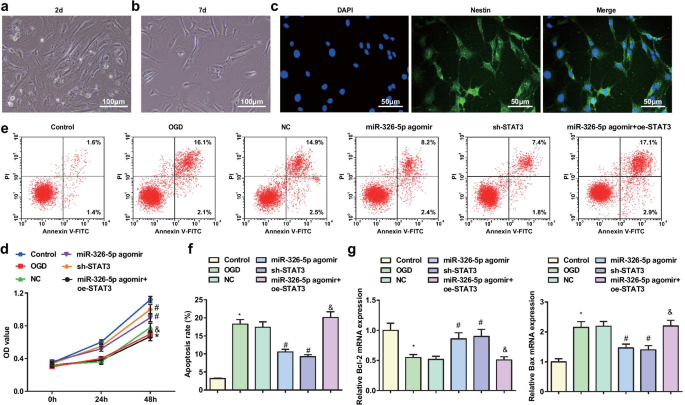
Повышающая регуляция miR-326-5p или понижающая регуляция STAT3 ослабляет апоптоз кортикальных нейронов, обработанных OGD. а Церебральные нейроны с двумерной прилипшей культурой (масштабная линейка:100 мкм); б Церебральные нейроны с 7-дневной прилипшей культурой (шкала:100 мкм); c Церебральные нейроны с 7-дневной адгезивной культурой, окрашенные иммунофлуоресцентным анализом NES (шкала:50 мкм); г Жизнеспособность обработанных OGD кортикальных нейронов по методу MTT; е Апоптоз обработанных OGD кортикальных нейронов, обнаруженный с помощью проточной цитометрии; е Скорость апоптоза корковых нейронов, обработанных OGD; г Экспрессия мРНК Bcl-2 и Bax в нейронах коры головного мозга, обработанных OGD; * P <0,05 по сравнению с контрольной группой; # P <0,05 по сравнению с группой NC; & P <0,05 по сравнению с группой агомира miR-326-5p
МТТ-анализ применяли для определения жизнеспособности кортикальных нейронов (рис. 1d). Результаты продемонстрировали, что жизнеспособность кортикальных нейронов крыс была нарушена в группе OGD и группе miR-326-5p агомир + oe-STAT3 по сравнению с контрольной группой и группой агомира miR-326-5p, но повышена в группе miR-326-5p. группа agomir и группа sh-STAT3 по сравнению с группой NC (все P <0,05).
Апоптоз кортикальных нейронов крыс определяли с помощью проточной цитометрии, а экспрессию мРНК Bcl-2 и Bax с помощью RT-qPCR (рис. 1e-g). Результаты объясняют, что по сравнению с контрольной группой и группой агомира miR-326-5p, группа OGD и группа агомир miR-326-5p + oe-STAT3 продемонстрировали повышенную скорость апоптоза, снижение экспрессии Bcl-2 и увеличение экспрессии Bax. (все P <0,05). Напротив, в группах NC скорость апоптоза была снижена, уровень Bcl-2 повышен, а уровень Bax был подавлен в группе агомира miR-326-5p и группе sh-STAT3 (все P <0,05).
Повышающая регуляция miR-326-5p или понижающая регуляция STAT3 увеличивает Mfn2 и уровень потенциала митохондриальной мембраны в корковых нейронах после повреждения OGD
Экспрессия Mfn2 в нейронах коры головного мозга, обработанных OGD, была обнаружена с помощью вестерн-блоттинга (рис. 2a, b). Что касается контрольной группы и группы агомира miR-326-5p, экспрессия белка Mfn2 снижалась в группе OGD и группе агомира miR-326-5p + oe-STAT3, тогда как повышалась в группе агомира miR-326-5p и sh -STAT3 группа по сравнению с NC-группой (все P <0,05).
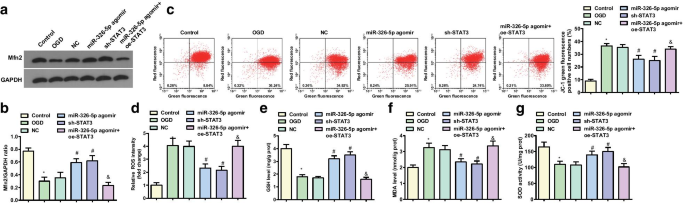
Повышающая регуляция miR-326-5p или понижающая регуляция STAT3 увеличивает Mfn2 и уровень митохондриального мембранного потенциала в обработанных OGD кортикальных нейронах. а , b Экспрессия белка Mfn2 в нейронах коры головного мозга, обработанных OGD; c JC-1 проточная цитометрия уровня митохондриального мембранного потенциала в обработанных OGD корковых нейронах; г Содержание ROS в нейронах коры головного мозга, обработанных OGD, после повышающей регуляции miR-326-5p или понижающей регуляции STAT3; е Активность GSH в обработанных OGD корковых нейронах после повышающей регуляции miR-326-5p или понижающей регуляции STAT3; е Содержание MDA в нейронах коры головного мозга, обработанных OGD, после повышающей регуляции miR-326-5p или понижающей регуляции STAT3; г Активность SOD в нейронах после повышающей регуляции miR-326-5p или понижающей регуляции STAT3; * P <0,05 по сравнению с контрольной группой; # P <0,05 по сравнению с группой NC; & P <0,05 по сравнению с группой агомира miR-326-5p
Потенциал митохондриальной мембраны, обнаруженный с помощью JC-1, обнаружил, что (рис. 2c, d) относительно контрольной группы и группы агомира miR-326-5p уровень митохондриального мембранного потенциала снизился в группе OGD и агомире miR-326-5p. + Группа oe-STAT3 (все P <0,05). По сравнению с группами NC, уровень митохондриального мембранного потенциала увеличился в группе агомира miR-326-5p и группе sh-STAT3 (все P <0,05).
Было определено содержание ROS и MDA, а также активности GSH и SOD в обработанных OGD кортикальных нейронах, и результаты показали, что по сравнению с контрольной группой и группой агомира miR-326-5p, группой OGD и агомиром miR-326-5p + oe -Группа STAT3 представила повышенное содержание ROS и MDA и нарушение активности GSH и SOD (все P <0,05). По сравнению с группами NC, группа агомира miR-326-5p и группа sh-STAT3 показали снижение содержания ROS и MDA и усиление активности GSH и SOD (все P <0,05) (рис. 2д – ж).
Повышающая регуляция miR-326-5p или понижающая регуляция STAT3 препятствует патологическому повреждению корковых нейронов и подавляет апоптоз у крыс с CI / RI
Окрашивание HE было принято для наблюдения за патологическим повреждением тканей головного мозга (рис. 3a, b), показывая, что в фиктивной группе нейроны имели нормальную структуру и аккуратно располагались с бледно-красной цитоплазмой, синим ядром и прозрачным ядрышком. В группе MCAO, группе NC и группе miR-326-5p agomir + oe-STAT3, за исключением некроза, наблюдалось, что нейроны были беспорядочно расположены и окрашены в более темный цвет, с разрывом ядерной мембраны, исчезновением клеточной структуры, кариопикнозом, глубоким окрашенное ядро и лизис в большом количестве. В группе агомира miR-326-5p и группе sh-STAT3 набухание нейронов было облегчено, нейроны были упорядочены, а количество некротических клеток уменьшилось, что указывает на лучший статус по сравнению с группой MCAO и группой NC.
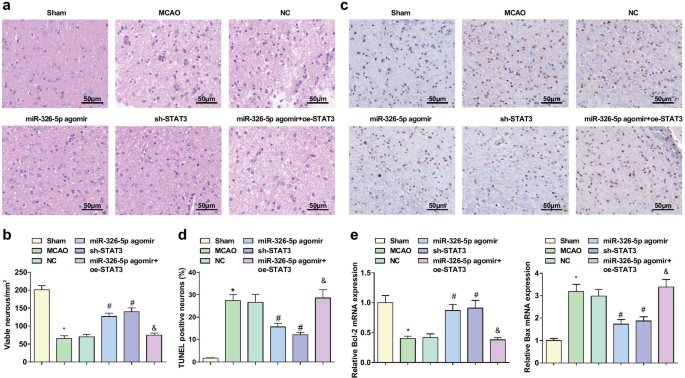
Повышающая регуляция miR-326-5p или понижающая регуляция STAT3 препятствует патологическому повреждению кортикальных нейронов и подавляет апоптоз у крыс с CI / RI. а Патологическое повреждение тканей головного мозга крыс MCAO, наблюдаемое по окрашиванию HE (шкала:50 мкм); б Число интактных нейронов в тканях мозга крыс MCAO после повышающей регуляции miR-326-5p или понижающей регуляции STAT3; c Апоптоз нейронов в тканях мозга крыс MCAO, обнаруженный с помощью окрашивания TUNEL (шкала:50 мкм); г TUNEL-положительные показатели в тканях мозга крыс MCAO после повышающей регуляции miR-326-5p или понижающей регуляции STAT3; е Экспрессия мРНК Bcl-2 и Bax в тканях мозга крыс MCAO после повышающей регуляции miR-326-5p или понижающей регуляции STAT3; * P <0,05 по сравнению с фиктивной группой; # P <0,05 по сравнению с группой NC; & P <0,05 по сравнению с группой агомира miR-326-5p
Окрашивание TUNEL использовали для обнаружения апоптоза нейронов и ОТ-КПЦР для экспрессии мРНК Bcl-2 и Bax в тканях мозга. Результаты показали, что (рис. 3c-e) по сравнению с фиктивной группой и группой агомира miR-326-5p, частота TUNEL-положительных результатов увеличивалась, экспрессия мРНК Bcl-2 снижалась, а экспрессия мРНК Bax увеличивалась в группе MCAO и группа miR-326-5p agomir + oe-STAT3 (все P <0,05). Более низкая частота TUNEL-положительных результатов и экспрессия мРНК Bax, а также более высокая экспрессия мРНК Bcl-2 были обнаружены в агомире miR-326-5p и группе sh-STAT3, чем в группе NC (все P <0,05).
Повышающая регуляция miR-326-5p или понижающая регуляция STAT3 увеличивает потенциальный уровень митохондриальной мембраны в тканях мозга у крыс с CI / RI
Обнаружение экспрессии белка Mfn2 в ткани головного мозга крысы было выполнено с помощью иммуногистохимии, и результаты показали, что (рис. 4a, b) более низкая экспрессия белка Mfn2 была протестирована в фиктивной группе и группе агомира miR-326-5p по сравнению с группой MCAO и группой агомира miR-326-5p. miR-326-5p agomir + oe-STAT3 группа, в то время как более высокая экспрессия белка Mfn2 была измерена в группе agomir miR-326-5p и группе sh-STAT3 вместо группы NC (все P <0,05).
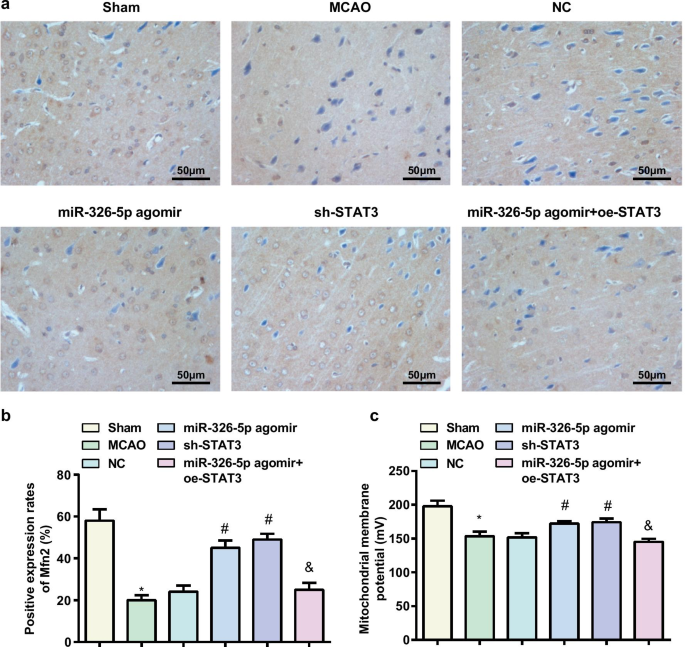
Повышающая регуляция miR-326-5p или понижающая регуляция STAT3 увеличивает уровень потенциала митохондриальной мембраны в тканях мозга у крыс с CI / RI. а Экспрессия Mfn2, обнаруженная с помощью иммуногистохимии в ткани мозга крысы (масштабная линейка:50 мкм); б Количество Mfn2-положительных клеток в тканях мозга крыс MCAO после повышающей регуляции miR-326-5p или понижающей регуляции STAT3; c Митохондриальные трансмембранные потенциалы в нейронах тканей мозга крыс MCAO после повышающей регуляции miR-326-5p или понижающей регуляции STAT3; * P <0,05 по сравнению с фиктивной группой; # P <0,05 по сравнению с группой NC; & P <0,05 по сравнению с группой агомира miR-326-5p
Обнаружение уровня митохондриального мембранного потенциала показало, что (рис. 4c) по сравнению с фиктивной группой и группой агомира miR-326-5p, уровень митохондриального мембранного потенциала снизился в группе MCAO и агомире miR-326-5p + oe-STAT3. в группе агомира miR-326-5p и группе sh-STAT3, в отличие от группы NC (все P <0,05).
miR-326-5p нацеливается на STAT3
Экспрессию miR-326-5p и STAT3 в корковых нейронах и тканях мозга определяли с помощью RT-qPCR и вестерн-блоттинга. В корковых нейронах in vitro предполагалось снижение экспрессии miR-326-5p и увеличение экспрессии STAT3 в группе OGD по сравнению с контрольной группой (обе P <0,05). Что касается группы NC, экспрессия miR-326-5p повышена, а экспрессия STAT3 снижена в группе агомира miR-326-5p (обе P <0,05); Выражение STAT3 уменьшилось ( P <0,05) в группе sh-STAT3. В отличие от группы агомира miR-326-5p, экспрессия STAT3 была повышена в группе агомира miR-326-5p + oe-STAT3 ( P <0,05) (рис. 5a – c).
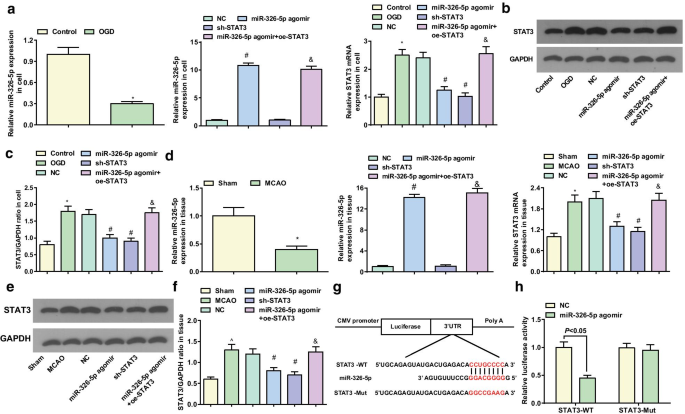
miR-326-5p нацелен на STAT3. а экспрессия мРНК miR-326-5p и STAT3 в обработанных OGD кортикальных нейронах in vitro; б , c Экспрессия белка STAT3 в обработанных OGD корковых нейронах in vitro; г экспрессия мРНК miR-326-5p и STAT3 в тканях мозга крыс MCAO; е , f Экспрессия белка STAT3 в тканях мозга крыс MCAO; г Предсказанный сайт связывания miR-326-5p и STAT3 с помощью программного обеспечения для биоинформатики; ч Отношения нацеливания между miR-326-5p и STAT3 подтверждены двойным анализом репортерного гена люциферазы; На рисунке а - c , * P <0,05 по сравнению с контрольной группой; # P <0,05 по сравнению с группой NC; & P <0,05 по сравнению с группой агомира miR-326-5p. На рисунке d - е , * P <0,05 по сравнению с фиктивной группой; # P <0,05 по сравнению с группой NC; & P <0,05 по сравнению с группой агомира miR-326-5p
В тканях мозга in vivo по отношению к фиктивной группе более низкий уровень miR-326-5p и более высокий STAT3 появлялись в группе MCAO (обе P <0,05). По сравнению с группой NC, экспрессия miR-326-5p выросла, в то время как экспрессия STAT3 снизилась в группе агомира miR-326-5p (обе P <0,05). Экспрессия STAT3 снизилась в группе sh-STAT3 ( P <0,05). Напротив, в группе агомира miR-326-5p экспрессия STAT3 повышена в группе агомира miR-326-5p + oe-STAT3 ( P <0,05) (рис. 5г – е).
Возможные сайты нацеливания miR-326-5p на STAT3 3'UTR были обнаружены с помощью программного обеспечения DIANA и miRDB (рис. 5g). STAT3 3'UTR, содержащий сайт связывания miR-326-5p WT и сайт связывания Mut, были сконструированы и вставлены в плазмиду pmirGLO. Связывание miR-326-5p с STAT3 3'UTR было дополнительно подтверждено анализом люциферазного репортера. Анализ люциферазного репортера показал, что агомир miR-326-5p снижает относительную люциферазную активность STAT3-WT, но не STAT3-Mut (рис. 5h), что означает, что STAT3 был прямым геном-мишенью для miR-326-5p.>
Обсуждение
CI / RI является ведущей причиной смерти от цереброваскулярных заболеваний [22]. Показано, что miRNAs участвуют в CI / RI [23]. Однако всестороннее понимание механизма, связанного с miR-326-5p, все еще недостаточно для CI / RI. Таким образом, это исследование нацелено на выяснение конкретных ролей miR-326-5p в CI / RI с акцентом на комбинированном реципроке miR-326-5p и STAT3. Продуктивно это исследование показало, что повышающая регуляция miR-326-5p ослабляет CI / RI и повышает экспрессию Mfn2 посредством ингибирования STAT3.
Вначале было определено, что экспрессия miR-326-5p снижается в CI / RI in vivo и in vitro. Впоследствии мы организовали серию анализов и обнаружили, что повышающая регуляция miR-326-5p увеличивает жизнеспособность нейронов и функцию митохондрий, повышает экспрессию Mfn2 и сдерживает окислительный стресс и апоптоз. Кроме того, эксперименты in vivo на крысах дополнительно подтвердили функциональную роль восстановленной miR-326-5p в CI / RI. As a matter of fact, high miR-326 is proved to correlate with long overall survival of patients with glioblastoma, the common type of brain tumor [24]. Also, miR-326 expression has been further evidenced to reduce in Parkinson's disease and miR-326 could suppress apoptosis of dopaminergic neurons and reduce inflammatory response [25]. Except for that, the reduction in miR-326-5p is manifested in endothelial progenitor cells in the course of myocardial infarction, and introduction of endothelial progenitor cells overexpressing could promote cardiac function recovery [11]. There has been a study illustrating that up-regulation of miR-326 improves the behavioral function, enhances neuronal viability and depresses neuronal apoptosis in mice with Alzheimer's disease [26]. Furthermore, it is documented that up-regulation of miR-326 by miR-326 mimic could suppress inducible nitric oxide synthase in dopaminergic neurons in Parkinson's disease [27].
In this study, we found that STAT3 was a target gene of miR-326-5p. Supported by an advanced study, it is indicated that STAT3 expression is negatively regulated by miR-326 in human endometrial carcinoma stem cells [28]. In the present study, we measured that STAT3 expression was enhanced in CI/RI. Currently, it has been found that I/R injury and OGD would induce STAT3 expression to increase [29]. Intriguingly, STAT3 mRNA expression trends to an increment in I/R animals [30]. Additionally, STAT3 protein expression is reported to up-regulate in CI/RI [31]. Next, our study revealed that down-regulating STAT3 had the therapeutic effects on CI/RI rats and OGD-treated neurons. As supported by a study, it is concluded that miR-31 induction discourages oxidative stress by inactivation of JAK/STAT3 pathway in ischemic stroke [32]. Also, inhibition of JAK2/STAT3 pathway is beneficial to oxidative stress impairment in CI/RI [31]. Furthermore, knockdown of JAK/STAT3 pathway is identified to enhance cell viability, and reduce oxidative stress and neuron apoptosis in ischemic brain injury [33]. Lately, inhibited JAK/STAT3 signaling pathway is witnessed to relieve myocardial infarction in myocardial I/R injury [34]. As demonstrated, depleted STAT3 undermines neuronal apoptosis in rats with white matter injury [35]. Moreover, the suppression of neuronal apoptosis, and alleviation of cerebral infarct size are attributed to JAK2/STAT3 inhibition in rats with CI/RI [36].
Finally, we focused on the effects of miR-326-5p and STAT3 on Mfn2 expression and discovered that the reduced level Mfn2 in rats with CI/RI and OGD-treated neurons could be elevated by either restoring miR-326-5p or silencing STAT3. As implicated by a previous study, it is documented that Mfn2 is decreased in the lately phase of reperfusion and poorly expressed of Mfn2 exacerbates CI/RI via restrained autophagosome formation and autophagosome and lysosome fusion [18]. Additionally, Mfn2 is implied to reduce even disappear in I/R injury in old hepatocytes [37]. However, few researches have discussed the regulatory mechanism of miR-326-5p and STAT3 for Mfn2.
Conclusion
In general, this study has elucidated the mechanisms that elevated miR-326-5p inhibits neuronal apoptosis, attenuates pathological damage of neurons and increases the expression of Mfn2 via STAT3 downregulation in CI/RI. The study may update the potential mechanism of miR-326-5p and STAT3 in CI/RI. However, more studies are still needed for further development of the molecular mechanism in CI/RI.
Доступность данных и материалов
Not applicable.
Сокращения
- miRNAs:
-
MicroRNAs
- STAT3:
-
Signal transducer and activator of transcription-3
- OGD:
-
Oxygen and glucose deprivation
- MCAO:
-
Middle cerebral artery occlusion
- ROS:
-
Reactive oxygen species
- STAT:
-
Signal transducers and transcriptional activator
- Mfn2:
-
Mitofusin-2
- FITC:
-
Fluorescein isothiocyanate
- NES:
-
Nestin
- NC:
-
Negative control
- Oe:
-
Overexpression
- RT-qPCR:
-
Reverse transcription quantitative polymerase chain reaction
- OD:
-
Оптическая плотность
- PI:
-
Иодид пропидия
- MDA:
-
Malondialchehyche
- GSH:
-
Glutathione
- SOD:
-
Superoxide dismutase
- DAB:
-
Diaminobenzidine
- PVDF:
-
Поливинилиденфторид
- WT:
-
Wild-type
- Mut:
-
Mutant
- SD:
-
Стандартное отклонение
- ANOVA:
-
One-way analysis of variance
Наноматериалы
- Ущерб от огня и предотвращение
- Понижающая регуляция микроРНК-342-5p или повышающая регуляция Wnt3a ингибирует ангиогенез и поддерживает стабиль…
- Получение наночастиц mPEG-ICA, нагруженных ICA, и их применение в лечении LPS-индуцированного повреждения клеток H9c2…
- Гибридный композит на основе наноструктурированного диоксида кремния / золота и целлюлозы с амино-POSS, получе…
- Повышение противоопухолевой эффективности и фармакокинетики буфалина с помощью пегилированных липосом
- Легкий синтез червоточиноподобного мезопористого оксида олова за счет самосборки, вызванной испарением, и у…
- Разработка и регулировка работы выхода графена с помощью размера, модификации, дефектов и легирования:исслед…
- Новые наночастицы, нацеленные на двойные митохондрии и рецепторы CD44, для высвобождения, инициируемого окисли…
- Повышенная протонная проводимость и снижение проницаемости для метанола с помощью биомембраны из альгината…
- Материал и оптические свойства флуоресцентных углеродных квантовых точек, полученных из лимонного сока поср…