


Оценка активности цитохрома P450 3A4, ингибируемого наночастицами золота, и молекулярных механизмов, лежащих в основе его клеточной токсичности, в клеточной линии C3A гепатоцеллюлярной карциномы человека
Аннотация
Взаимодействие наночастиц золота (AuNP) размером 40 и 80 нм, функционализированных катионным разветвленным полиэтиленимином (BPEI), анионной липоевой кислотой (LA) или нейтральным полиэтиленгликолем (PEG), с клеточной линией C3A гепатоцеллюлярной карциномы человека (HCC) было исследовано в отсутствие и наличие короны белков плазмы крови человека (ПК). Все чистые (без PC) AuNP, кроме 80 нм LA-AuNP, были цитотоксичны по отношению к C3A, но PC ослабляли их цитотоксичность. Зависящее от времени поглощение клетками AuNP увеличивалось помимо 40 нм BPEI-AuNP, но PC подавляло их поглощение помимо 80 нм PEG-AuNP. Двухфазные ответы на окислительный / нитрозативный стресс с помощью BPEI-AuNP наблюдались в клетках C3A, тогда как PEG-AuNP был мощным антиоксидантом. Все незащищенные AuNP ингибировали активность цитохрома P450 (CYP) 3A4 независимо от размера и поверхностного заряда, но PC восстанавливал свою активность помимо PEG-AuNP. Экспрессия гена, модулированная 40 нм PEG-AuNP, в основном участвует в β-окислении митохондриальных жирных кислот и, в меньшей степени, в переносчиках оттока / захвата через печень. Эти исследования способствуют лучшему пониманию взаимодействия AuNP с ключевыми биологическими процессами и лежащими в их основе молекулярными механизмами при ГЦК, что может быть в дальнейшем задействовано в разработке более эффективных терапевтических мишеней для лечения ГЦК.
Фон
Гепатоцеллюлярная карцинома (ГЦК) - один из наиболее распространенных онкологических заболеваний во всем мире и наиболее быстро растущая причина смертности от рака в США [1, 2]. Учитывая, что ГЦК диагностирован на поздних стадиях, лечебные методы лечения ГЦК включают трансплантацию печени или хирургическую резекцию на ранней стадии развития опухоли, а также химио- и лучевую терапию при запущенном состоянии опухоли. ГЦК часто развивает высокую устойчивость к обычным противоопухолевым агентам, неселективной цитотоксической молекуле, которая может приводить к системным побочным эффектам. Последние достижения в генной терапии, то есть генная терапия на основе РНК-интерференции (РНКи), были использованы в текущем лечении ГЦК [3, 4]. Эффективность РНКи требует доставки вектора внутрь клетки-мишени [5]. Векторами для успешной доставки генов являются вирусные и невирусные векторы. Вирусы предлагают более высокую эффективность доставки генов, но невирусные векторы предпочтительны из-за проблем с безопасностью вирусных векторов. Наночастицы (NP) как невирусные векторы для адресной доставки генов или системы доставки лекарств привлекли большое внимание для повышения терапевтической эффективности и снижения токсичности на системном и / или клеточном уровнях при лечении ГЦК [4, 6]. Таким образом, становится очень важным определить молекулярный механизм и биологический путь, лежащий в основе клеточных нарушений и токсичности NP в клетках-мишенях и тканях. Недавние исследования in vitro показали, что профилирование экспрессии генов в сочетании с клеточными и биохимическими ответами обеспечивает прямую оценку клеточных нарушений и потенциальной токсичности NP [7,8,9,10].
Золотые наночастицы (AuNP) использовались в качестве носителя для доставки для целенаправленной доставки фрагментов, подавляющих гены, по отдельности или в комбинации с другими лекарствами благодаря их уникальным физико-химическим свойствам и химическому составу поверхности [11, 12]. Взаимодействие AuNP с белками плазмы крови формирует белковую корону, которая, в свою очередь, изменяет химический состав поверхности NP и влияет на последующие биологические реакции, такие как клеточное поглощение и потенциальная токсичность [13, 14]. На клеточное поглощение AuNP в различных линиях раковых клеток человека и первичных клетках критически влияет формирование короны белка, независимо от размера и поверхностного заряда [7,8,9, 14,15,16,17].
Окислительный стресс, зависящий от размера и поверхностного заряда, также наблюдался в клеточной линии рака молочной железы человека, MDA-MB-231, гепатоцеллюлярной карциноме HepG2 и клетках лейкемии человека HL-60 в ответ на AuNP, что было связано с цитотоксичностью NP [18 , 19]. Цитотоксичность, вызванная AuNP, проявлялась в различных линиях раковых клеток человека и первичных клетках человека в зависимости от типа клеток [7,8,9, 20, 21].
Ферменты цитохрома P450 (CYP) играют важную роль в биоактивации или инактивации многих цитотоксических препаратов, а также в повышении чувствительности организма к канцерогенности противоопухолевых препаратов [22]. AuNP влиял на каталитическую активность ферментов CYP на клеточном и молекулярном уровнях in vivo и in vitro [7, 23,24,25]. AuNP в значительной степени продемонстрировал дифференциальную экспрессию генов, преимущественно участвующих в маркерах окислительного стресса в линии клеток фибробластов легких человека MRC-5, и митохондриальную дисфункцию в клетках пупочной вены человека (HUVEC) и гепатоцитах человека, что коррелирует с увеличением продукции перекиси липидов и высокая цитотоксичность [8, 9, 26]. Хотя эти знания взаимно предполагают, что AuNP вызывает апоптотическую или некротическую гибель клеток в различных типах клеток и изменяет клеточные и биохимические функции в сочетании с дифференциальной экспрессией генов в путях стрессовой реакции и токсичности, конкретные пути, посредством которых AuNP проявляет свои токсические эффекты внутри клетки или биологических система остается неизвестной.
В данном исследовании изучали влияние короны, размера и поверхностного заряда белка на взаимодействие AuNP с клеткой C3A человека HCC. В первую очередь, зависящее от времени поглощение клетками AuNP 40 и 80 нм, функционализированных катионным BPEI, анионной липоевой кислотой (LA) или нейтральным полиэтиленгликолем (PEG) в клетках C3A, определяли с и без короны белков плазмы крови человека (PC). Во-вторых, отслеживались индуцированная AuNP цитотоксичность и продукция активных форм кислорода (ROS) / активных форм азота (RNS), а также их ингибирующее действие на активность CYP3A4. Наконец, молекулярный механизм действия, связанный с токсичностью AuNP, был охарактеризован с использованием программы Human Molecular Toxicology Pathway Finder и Human Drug Transporters RT 2 ПЦР-массив Profiler ™.
Методы
Синтез наночастиц золота
Катионный BPEI с длиной волны 40 и 80 нм, анионный LA и нейтральный PEG Biopure ™ AuNP были синтезированы на заказ из nanoComposix (Сан-Диего, Калифорния). Размер частиц, индекс полидисперсности (PDI), дзета (z) -потенциал и спектральные свойства были охарактеризованы с помощью динамического рассеяния света (DLS), просвечивающей электронной микроскопии (TEM) и УФ-видимой спектроскопии. AuNP были синтезированы восстановлением гидрата тетрахлораурата (III) водорода в водном растворе карбоната калия с последующим процессом старения и фильтрацией в тангенциальном потоке (TFF). Поверхность AuNP функционализировали с помощью LA или PEG путем добавления дигидролипоевой кислоты (0,2:1, w / w ) или ПЭГ с концевыми тиолметоксигруппами (Laysan Bio Inc., Arab, AL) (0,5:1, w / w ) соответственно с последующей промывкой TFF и стерильной фильтрацией. Функционализированные BPEI поверхности AuNP были синтезированы с помощью химии EDC путем связывания карбоновой кислоты LA со свободными аминами BPEI с последующей промывкой TFF и последующим центрифугированием для удаления несвязанного BPEI.
Приготовление протеиновой короны
Объединенная плазма крови человека ( n =5) были получены от Biological Specialty Corp. (Кольмар, Пенсильвания). AuNP инкубировали в плазме человека (55%, v / v ) с постоянной скоростью 250 об / мин при 37 ° C в течение 1 ч, как сообщалось [7, 8]. Несвязанные и слабо связанные белки удаляли повторными промывками фосфатно-солевым буфером (PBS) при 20 000 × г в течение 20 мин при 20 ° C. Конечный AuNP, покрытый корой (PC) белков плазмы крови человека, диспергировали в PBS и затем разбавляли в среде для культивирования клеток для дальнейшей физико-химической характеристики или дозирования. Подробный протокол приведен в Дополнительном файле 1.
Физико-химическая характеристика AuNP
Гидродинамические диаметры ( D H ), PDI и z-потенциал 40- и 80-нм чистого (без PC) AuNP, функционализированного BPEI, LA и PEG в деионизированной (DI) воде, анализировали при 25 ° C в течение 0 часов с использованием Zetasizer Nano ZS (Malvern Instruments, Вустершир, Великобритания); для покрытого ПК AuNP в PBS при 25 ° C в течение 0 часов; и для всех чистых и PC AuNP в полной среде для культивирования клеток при 37 ° C через 0 и 24 часа. Полная среда для культивирования клеток содержала минимальную необходимую среду Игла (EMEM) с добавлением 10% FBS (ATCC ® , Манассас, Вирджиния). Образец был измерен 5 раз с 11 частями по 10 с каждый. Кроме того, спектры оптического поглощения были измерены с использованием гибридного многомодового микропланшетного ридера Synergy H1 (BioTek Instruments Inc., Winooski, VT) при комнатной температуре в течение 0 часов.
Просвечивающий электронный микроскоп
Морфологию AuNP охарактеризовали с помощью просвечивающей электронной микроскопии. Раствор AuNP без покрытия и PC (5 мкл) помещали на медные сетки 200 меш с последующей сушкой на воздухе при комнатной температуре. Образцы просматривали на Tecnai G2 Spirit BioTWIN с детектором Oxford (FEI Company, Hillsboro, OR) при ускоряющем напряжении 120 кВ. Комплект для микроскопии GATAN (GATAN Inc., Плезантон, Калифорния) измерял диаметры AuNP.
Измерение культуры клеток и их жизнеспособности
Клетки гепатоцеллюлярной карциномы человека C3A (ATCC ® CRL-10741 ™) были приобретены в ATCC ® (Манассас, Вирджиния), выращенные в полном EMEM (ATCC ® , Manassas, VA) с добавлением 10% FBS и увеличивался примерно до 80% слияния в колбе T75 со сменой среды каждые 4 дня. Через 0,25% ( w / v ) расщепление трипсин – 0,53 мМ этилендиаминтетрауксусной кислоты (ЭДТА), клетки высевали в 96-луночные планшеты при 8 × 10 4 клеток на лунку и инкубируют при 37 ° C во влажной атмосфере, состоящей из 95% воздуха и 5% CO 2 После 48-часовой инкубации в клетки добавляли AuNP в отсутствие и в присутствии ПК. Клетки C3A между 9 и 12 пассажами использовали для дозирования.
Жизнеспособность C3A определялась с использованием alamarBlue ® анализ жизнеспособности (Thermo Sci., Waltham, MA), как описано [7, 27]. Клетки в 96-луночных планшетах обрабатывали 40 и 80 нм BPEI-, LA- и PEG-AuNP с ПК и без него в диапазоне от 0 до 250 мкг / см 2 . Через 24 часа 10% alamarBlue ® реагент в комплекте EMEM ( v / v ) добавляли к культуре клеток и инкубировали 3 ч при 37 ° C. Полный EMEM служил диспергатором. Взаимодействие AuNP с активным ингредиентом alamarBlue ® реагент, резазурин или восстановленный продукт, резоруфин, измеряли в качестве контролей. AuNP и резазурин (без клеток) или поддерживающая среда (без клеток) служили в качестве фонового контроля. Флуоресценция, пропорциональная жизнеспособности клеток, была нормализована к контролю и выражена в процентах относительно контрольной группы клеток.
Измерение клеточного поглощения с помощью масс-спектрометрии с индуктивно связанной плазмой
Клетки были засеяны размером 8 × 10 4 . клеток на лунку 96-луночных планшетов с нетоксичной концентрацией 1,56 мкг / см 2 всего чистого и ПК AuNP в течение 0,5, 1, 3, 6, 12 и 24 часов. Стадия травления была включена для удаления AuNP, связанного с клеточной мембраной, и его неспецифического связывания с лунками, как сообщалось ранее [28]. Сбор клеток сушили и переваривали в царской водке, а внутриклеточную концентрацию Au определяли количественно с помощью NexION ™ 350X масс-спектрометрия с индуктивно связанной плазмой (ICP-MS) (PerkinElmer, Waltham, MA). Поглощение AuNP клетками рассчитывали, как сообщалось ранее, и выражали как количество AuNP на клетку [29]. Подробный протокол приведен в Дополнительном файле 1.
Измерения окислительного / нитрозативного стресса
Клетки были засеяны размером 8 × 10 4 . клеток на лунку 96-луночных планшетов с дозировкой 40 нм чистого BPEI- и PEG-AuNP до 125 мкг / см 2 на 1, 3 и 24 часа. Прямое измерение кислородного / нитрозативного стресса проводили с помощью набора для анализа общего количества активных форм кислорода (ROS) / супероксида (SO) (Enzo Life Sciences, Фармингдейл, штат Нью-Йорк), как описано ранее [30]. Флуоресценцию, пропорциональную увеличению ROS / активных форм азота (RNS) (Ex488 / Em520 нм) или SO (Ex550 / Em610 нм), измеряли с помощью считывающего устройства для микропланшетов. Подробный протокол приведен в Дополнительном файле 1.
Активность цитохрома P450 3A4
Неблагоприятные эффекты 40- и 80-нм чистого и PC AuNP на активность CYP3A4 были охарактеризованы с помощью анализов P450-Glo ™ (Promega Corp., Мэдисон, Висконсин), как полностью описано [7]. Клетки C3A в 96-луночных планшетах дозировали до средней летальной концентрации (LC 50 ) значения:127,3 мкг / см 2 40 нм BPEI-AuNP, 205,5 мкг / см 2 80 нм BPEI-AuNP, 192,5 мкг / см 2 40 нм LA-AuNP и 129,5 мкг / см 2 40 нм ПЭГ-AuNP. Начиная с LC 50 значения 80 нм LA- и PEG-AuNP не определялись, клетки обрабатывали LC 50 значения 40 нм LA- и PEG-AuNP (192,5 мкг / см 2 и 129,5 мкг / см 2 , соответственно). По окончании 24-часовой инкубации клетки инкубировали с субстратом CYP3A4 (люциферин-IPA) при 37 ° C в течение 3 часов. Сигнал люминесценции, пропорциональный активности фермента, измеряли с помощью считывающего устройства для микропланшетов и затем нормализовали для контроля. Контроли были назначены для оценки взаимодействия AuNP с родительскими субстратами или метаболитами и бесклеточными субстратами. Активность CYP выражалась в процентах относительно контрольной группы клеток.
Профилирование экспрессии генов
Поскольку токсичный 40 нм PEG-AuNP использовался для ингибирования активности CYP3A4 и антиоксидантной активности в клетках C3A, демонстрирующих высокое клеточное поглощение, он был выбран для характеристики молекулярных механизмов действия, лежащих в основе его токсичности и дифференциальных клеточных ответов. Клетки были засеяны размером 2,5 × 10 6 . клеток на лунку 6-луночных планшетов с дозировкой LC 50 значение 40 нм PEG-AuNP в течение 24 ч при 37 ° C. В конце инкубации клетки подвергали выделению РНК, а затем проводили синтез кДНК с использованием общей РНК со средним числом целостности РНК (RIN) 7,8, как описано ранее [7,8,9]. Полученная кДНК была смешана с RT 2 . Мастер-микс SYBR green (Qiagen Inc., Валенсия, Калифорния), а затем нанесен на средство поиска путей молекулярной токсикологии человека или переносчики лекарств человека RT 2 Массивы ПЦР Profiler ™ в Quantstudio ™ 7 Flex (Applied BioSystem, Foster City, CA). Дифференциально экспрессируемые гены со кратностью изменения <- 2 и> 2 и p <0,05 представляет понижающую и повышающую регуляцию интересующего гена. Для проверки RT 2 Данные массива ПЦР, экспрессия девяти выбранных генов оценивалась с помощью синтеза кДНК и последующей ПЦР в реальном времени. Последовательности праймеров приведены в Дополнительном файле 1:Таблица S1. Все реакции ПЦР проводили в трех экземплярах. Подробный протокол условий и количественной оценки ПЦР в реальном времени приведен в Дополнительном файле 1.
Статистический анализ
Средняя летальная концентрация (LC 50 ) значения AuNP в клетках C3A были оценены путем подбора уравнения Хилла с переменным наклоном к наблюдаемым данным (ввод уровней концентрации AuNP и соответствующая жизнеспособность клеток) с использованием GraphPad Prism 6 (La Jolla, CA), как описано [7]. Односторонний дисперсионный анализ (ANOVA) был проведен с использованием SAS 9.4 (Институт SAS, Кэри, Северная Каролина) для оценки эффектов AuNP на продукцию ROS / RNS и клеточное поглощение в клетках C3A. Если значимо, множественное сравнение выполнялось с помощью теста честной значимой разницы Тьюки (HSD) при p <0,05.
Результаты и обсуждение
Физико-химическая характеристика AuNP на ПК без оболочки и плазмы человека
Влияние размера НЧ, поверхностного заряда и образования ПК в плазме человека вокруг AuNP на гидродинамический диаметр (D H ), индекс полидисперсности (PDI), z-потенциал и спектральные свойства, а также морфология были охарактеризованы с помощью DLS, ПЭМ и УФ-видимой спектроскопии (рис. 1). На изображениях ПЭМ все чистые (без ПК) AuNP в деионизированной воде были монодисперсными с плотным распределением по размерам и уникальными диапазонами спектра в УФ-видимой области 521–553 нм (рис. 1а). Образования ПК вокруг AuNP наблюдались с изменением распределения по размерам и красными смещениями спектров поглощения (рис. 1б). D H и значения PDI для 40 и 80 нм чистого и PC AuNP в полной EMEM были совместимы до 24 часов при 37 ° C, за исключением 40 и 80 нм PC BPEI-AuNP, которые показали снижение значений PDI (0,29 и 0,32, соответственно. ) через 24 ч при 37 ° C по сравнению с 0 ч при 37 ° C (0,62 и 1,0 соответственно) (Таблица 1). Значения Z-потенциала всех AuNP без покрытия и ПК относительно снизились через 24 часа при 37 ° C по сравнению с таковыми через 0 часов при 37 ° C. Наблюдалась агрегация 40- и 80-нм PC BPEI-AuNP в PBS и полной EMEM, что коррелировало с множественными пиками в распределении размеров и изменениями D H и красные смещения спектров поглощения относительно чистого BPEI-AuNP (рис. 1 и дополнительный файл 1:рис. S1, таблица 1). Эти результаты были подтверждены недавними исследованиями, в которых PC 40 и 80 нм и BPEI-AuNP, покрытый коронным разрядом человеческого сывороточного альбумина, были агрегированы в PBS и различных средах для культивирования клеток [7,8,9].
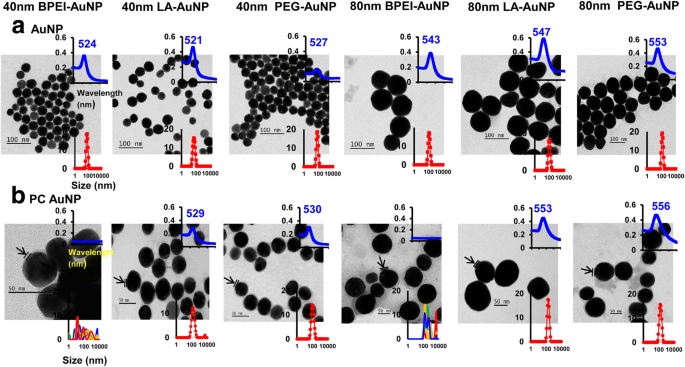
Электронные микрофотографии a AuNP в деионизированной воде и b PC AuNP в PBS при 0 ч при 25 ° C, длина волны УФ-видимого спектра (верхняя вставка) и распределение динамического светорассеяния (нижняя вставка). Стрелки указывают на формирование ПК. ПК протеин плазмы человека корона, BPEI разветвленный полиэтиленимин, LA липоевая кислота, ПЭГ полиэтиленгликоль
Цитотоксичность AuNP
Цитотоксичность AuNP измеряли с использованием средней летальной концентрации (LC 50 ) в клетках C3A. Заряд поверхности НЧ, размер частиц и образование ПК вокруг НЧ-зависимого ЖК 50 Анализы с AuNP показаны на фиг. 2. Все 40 нм BPEI-, LA и PEG-AuNP и 80 нм BPEI-AuNP были цитотоксичны по отношению к клеткам C3A с соответствующими LC 50 колеблется от 127,3 до 205,5 мкг / см 2 (Рис. 2а). 80 нм чистый ПЭГ-AuNP показал 59% жизнеспособности клеток при самой высокой концентрации 250 мкг / см 2 , тогда как 80 нм LA-AuNP не были цитотоксичными. ПК снижал токсичность AuNP в зависимости от размера и модификации поверхностного заряда, за исключением 80 нм BPEI-AuNP, который показал жизнеспособность 51% клеток при 250 мкг / см 2 через 24 ч (рис. 2б). Недавние исследования показали, что 40-нм чистый BPEI-AuNP был токсичным для первичных гепатоцитов человека, HUVEC и клеток проксимальных канальцев почек человека (HPTC) (LC 50 колеблется от 22,4 до 80,3 мкг / см 2 ) [7,8,9]. BPEI-AuNP, покрытый PC, был цитотоксичным для гепатоцитов человека, но AuNP, покрытый HSA, не был цитотоксичным [7]. Эти результаты свидетельствуют о том, что клетки C3A более устойчивы к токсичности AuNP, чем первичные клетки человека, из-за высокой скорости пролиферации и метаболической активности линии раковых клеток [31].
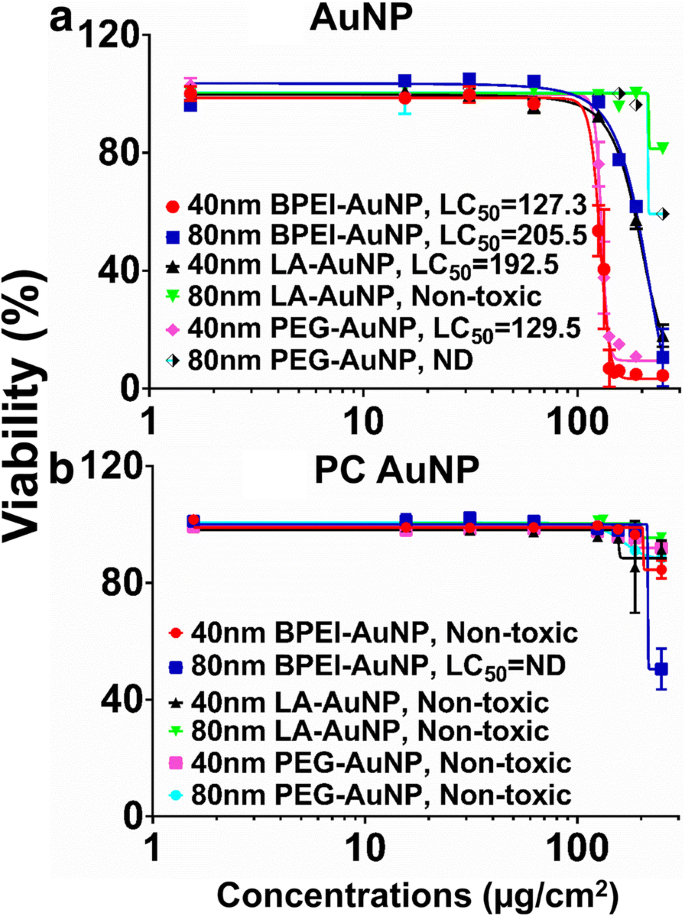
Жизнеспособность C3A и LC 50 значения 40 и 80 нм a AuNP и b ПК AuNP. Данные представляют собой среднее значение ± стандартное отклонение. ( нет =3). PC корона белка плазмы человека, ND не определено, разветвленный полиэтиленимин BPEI, липоевая кислота LA, полиэтиленгликоль PEG, LC 50 средняя летальная концентрация
Внутриклеточное поглощение AuNP
Размер НЧ, поверхностный заряд и Зависимое от ПК поглощение клетками всего чистого и ПК AuNP были определены при 1,56 мкг / см 2 до 24 ч. ANOVA показал значительные изменения в зависимости от размера, ПК и времени ( p <0,0001) и взаимодействия (ПК × размер, ПК × время, размер × время и ПК × размер × время) ( p <0,001) для всего поглощения AuNP, за исключением незначительного взаимодействия (ПК × размер) для поглощения LA- и PEG-AuNP ( p =0,2). Как показано на рис. 3a-f, линейное увеличение клеточного поглощения 40 и 80 нм чистого и ПК AuNP наблюдалось помимо 40 нм чистого и ПК BPEI-AuNP, которое достигло наивысшего клеточного поглощения через 6 часов и снизилось впоследствии ( Рис. 3а). Однако через 24 часа катионный BPEI-AuNP с длиной волны 40 нм содержал наибольшее поглощение, за которым следовал нейтральный PEG-AuNP с длиной волны 40 нм, а затем анионный LA-AuNP с длиной волны 40 нм, что было связано с порядком цитотоксичности AuNP для клеток C3A (рис. 2а). ). Этот результат согласуется с предыдущими исследованиями, согласно которым катионный поли (N- (2-аминоэтил) акриламид) и BPEI-AuNP имели наибольшее поглощение клетками по сравнению с таковыми анионной поли (акриловой кислоты) и LA-AuNP и нейтрального поли ( N- (2,3-дигидроксипропил) акриламид- и PEG-AuNP в клетках колоректальной аденокарциномы человека Caco-2, HPTC и гепатоцитах человека [9, 32]. Кроме того, комплекс NP-PC ослаблял все 40- и 80-нм BPEI- и LA-AuNP и 40 нм PEG-AuNP в клетках C3A, но ускоряют захват 80 нм PEG-AuNP (рис. 3f). Эти результаты подтверждаются недавними исследованиями, согласно которым ПК ингибировал поглощение AuNP в HUVEC, HEK и HPTC, независимо от размера и поверхностного заряда [8, 9, 33]. Напротив, коронки PC и HSA увеличивали поглощение 40 нм PEG-AuNP в гепатоцитах человека, но последние индуцировали поглощение 80 нм PEG-AuNP в HEK [7, 33] .
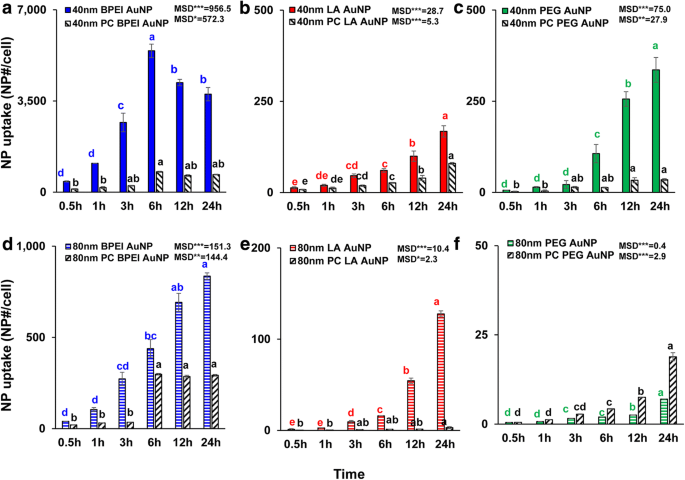
Зависящее от времени поглощение клетками 40 нм a BPEI-AuNP, b LA-AuNP и c PEG-AuNP и 80 нм d BPEI-AuNP, e LA-AuNP и f ПЭГ-AuNP в отсутствие и в присутствии ПК в клетках C3A до 24 часов. Данные представляют собой среднее значение ± стандартное отклонение. ( нет =3). Буквы значительно различались согласно тесту Тьюки HSD. BPEI разветвленный полиэтиленимин, LA липоевая кислота, ПЭГ полиэтиленгликоль, ПК корона белка плазмы человека, MSD минимально значимая разница. * p <0,05; ** p <0,005; *** p <0,0001
Измерения окислительного и нитрозионного стресса
Поскольку 40-нм чистый BPEI- и PEG-AuNP проявляли более высокую цитотоксичность и клеточное поглощение в клетках C3A по сравнению с другими AuNP, они были выбраны для исследования индуцированного AuNP окислительного / нитрозативного стресса. Оба AuNP модулировали генерацию ROS / RNS в клетках C3A в зависимости от времени и концентрации ( p <0,0001) и по взаимодействию (время × концентрация, p <0,0001). Как показано на рис. 4a, образование ROS / RNS уменьшалось при более высоких концентрациях 40 нм BPEI-AuNP (62,5 мкг / см 2 и 125 мкг / см 2 ) через 1 час при 37 ° C, но затем увеличивалась до 24 часов. Напротив, 40 нм PEG-AuNP существенно подавлял генерацию ROS / RNS при 39,1 мкг / см 2 и далее с кратностью изменения <0,5 до 24 ч (рис. 4b). Активация гибели клеток часто способствует токсичности NP, и в большинстве случаев увеличение продукции ROS / RNS, приводящее к оксидативному стрессу, отвечает за токсичность NP [34]. Зависимое от поверхностного заряда образование ROS / RNS наблюдали с 40 нм катионным BPEI- и нейтральным PEG-AuNP. 40 нм BPEI-AuNP показал двухфазный паттерн образования ROS / RNS (антиоксидант через 1 час и прооксидант через 3 часа и далее) при высоких концентрациях, что было связано с его цитотоксичностью в клетках C3A (рис. 2a). Этот результат согласуется с предыдущими исследованиями, согласно которым генерация АФК, индуцированная 40 и 80 нм BPEI-AuNP и 20 нм цитрат-AuNP, была связана с их цитотоксичностью в гепатоцитах человека и клетках HepG2 соответственно во времени и в зависимости от концентрации. [7, 35]. AuNP показал индуцированную окислительным стрессом цитотоксичность в клетках промиелоцитарного лейкоза человека, HL-60 с общим восстановлением глутатиона, независимо от размера [19]. Напротив, 40 нм PEG-AuNP служил антиоксидантом, что позволяет предположить, что окислительный / нитрозативный стресс не может быть прямым механизмом цитотоксичности, вызванной 40 нм PEG-AuNP в клетках C3A (рис. 2b).
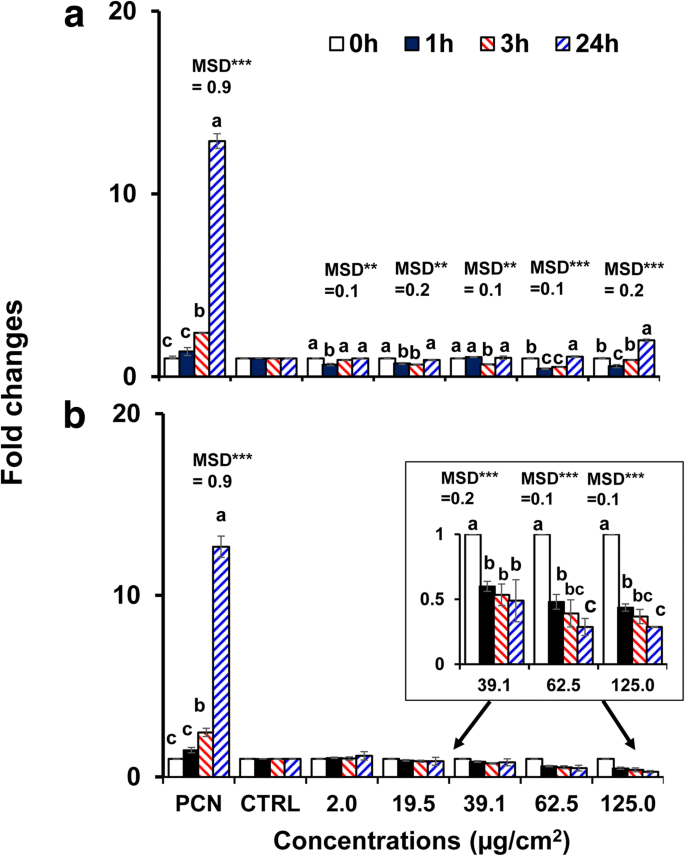
Производство ROS / RNS в зависимости от времени и концентрации в клетках C3A, подвергшихся воздействию a 40 нм BPEI-AuNP и b 40 нм PEG-AuNP до 24 часов. Данные представляют собой среднее значение ± стандартное отклонение. ( нет =3). Буквы значительно различались согласно тесту Тьюки HSD. BPEI разветвленный полиэтиленимин, LA липоевая кислота, ПЭГ полиэтиленгликоль, CTRL контроль, MSD минимальная значимая разница, PCN пиоцианин (индуктор АФК). ** p <0,005; *** p <0,0001
Измерение активности CYP3A4
Были охарактеризованы ингибирующие эффекты 40- и 80-нм чистого AuNP и PC AuNP на активность CYP3A4. Как показано на рис. 5, 40 нм BPEI-, LA- и PEG-AuNP и 80 нм BPEI-AuNP при LC 50 значения ингибировали каталитическую активность CYP3A4 в клетках C3A с соответствующей активностью от 20,1 до 31,4% относительно контроля, независимо от размера и заряда поверхности. Нетоксичные концентрации 80 нм LA- и PEG-AuNP также подавляли его активность (31,4 и 26,6% соответственно). Однако ПК значительно улучшил индуцированное AuNP 40 и 80 нм ингибирование CYP3A4, помимо PEG-AuNP 40 и 80 нм, показав активность 63 и 46% по сравнению с контролем. Это согласуется с исследованиями in vitro с тканью печени и гепатоцитами человека, согласно которым анионная дубильная кислота-AuNP и катионная 40- и 80-нм BPEI-AuNP существенно ингибируют каталитическую активность CYP3A4 [7, 25]. Напротив, катионный PEI-AuNP и нейтральный поливинилпирролидон-AuNP-индуцированная экспрессия мРНК CYP1A2, CYP2C9 и CYP3A4 в клетках HepG2 и CYP2B и CYP3A в срезе печени крысы, соответственно [36, 37]. В недавнем исследовании сообщается, что 40- и 80-нм чистый и PC BPEI-AuNP существенно подавляли активность CYP3A4 в гепатоцитах человека за счет конформационного изменения белка или блокирования кармана субстрата в качестве обратимого ингибирования [7].
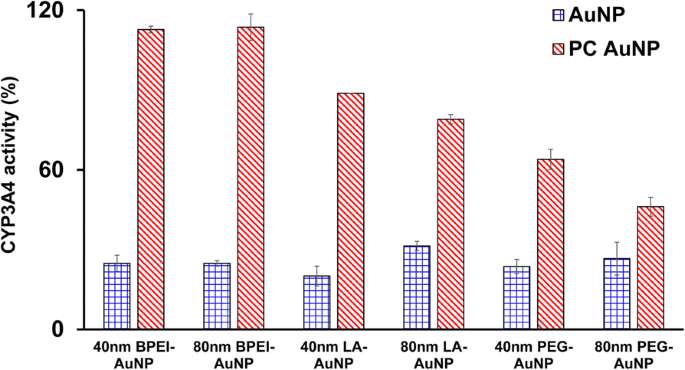
Ингибирующий эффект AuNP на активность CYP3A4 в клетках C3A, подвергнутых воздействию 40 и 80 нм BPEI-, LA- и PEG-AuNP в отсутствие и в присутствии PC в течение 24 часов. Значения представляют собой среднее ± стандартное отклонение. ( нет =3). BPEI разветвленный полиэтиленимин, LA липоевая кислота, ПЭГ полиэтиленгликоль, ПК белок плазмы человека корона
Профилирование экспрессии генов, ориентированное на токсический путь, 40 нм PEG-AuNP
Из репрезентативных генов, охватывающих 13 различных путей стресса и токсичности, всего 212 генов (↓ 186 и ↑ 26) дифференциально экспрессировались в LC 50 значение 40 нм PEG-AuNP (рис. 6, дополнительный файл 1:таблицы S2 – S7). 12,3% (26 генов, ↓ 26, ↑ 0 гены) от общего числа генов (212 генов) были преимущественно вовлечены в β-окисление митохондриальных жирных кислот; на апоптоз 11,3% (24 гена, ↓ 18, ↑ 6 генов); по пути повреждения и репарации ДНК - 11,3% (24 гена, ↓ 18, ↑ 6 генов); и для реакции теплового шока 11,3% (24 гена, ↓ 22, ↑ 2).
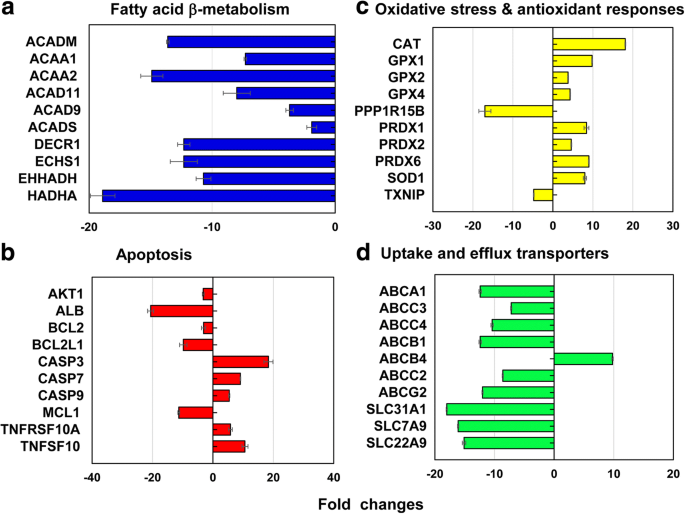
Репрезентативные гены, участвующие в a β-окисление митохондриальных жирных кислот, b апоптоз, c окислительный стресс и антиоксидантные реакции, и d печеночные транспортеры поглощения и оттока при LC 50 значение 40 нМ ПЭГ-AuNP. Все данные имели кратное изменение <- 2 и> 2 при p <0,05. Анализ онтологии генов приведен в Дополнительном файле 1:Таблицы S2 – S7
В митохондриальном пути β-окисления жирных кислот гены, кодирующие три различных фермента, участвующих в производстве ацил-КоА и восстанавливающих эквивалентов НАДН и ФАДН 2 в основном подавлялись; Гены ACAD11, ACAD9, ACADM и ACADS в ацил-CoA дегидрогеназах (от 2,0 до 13,6 раз); ACAA1 и ACAA2 в кетоацил-CoA тиолазах (от 7,3 до 14,9 раза); DECR1, ECHS1, EHHADH и HADHA (от 10,7 до 18,9 раза) в еноил-КоА гидратазе (рис. 6а, дополнительный файл 1:таблица S2). Β-окисление митохондриальных жирных кислот играет важную роль в производстве ацил-КоА и восстановлении эквивалентов НАДН и ФАДН 2 , который связан с четырьмя основными ферментами (ацил-CoA дегидрогеназы, еноил-CoA гидратазы, гидроксиацил-CoA дегидрогеназы и кетоацил-CoA тиолазы [38, 39]. Кроме того, переносчики электронов, NADH и FADH 2 , участвуют в цикле трикарбоновых кислот (ТСА) и дыхательной цепи митохондрий, что приводит к выработке АТФ. В текущем исследовании 40 нм ПЭГ индуцировал митохондриальную дисфункцию, потерю содержания АТФ за счет снижения внутриклеточных уровней АТФ и FADH 2 , следовательно, определяя его цитотоксичность в клетках C3 (рис. 2а). Подобный феномен был зарегистрирован в гепатоцитах человека, HUVEC и HPTC, подвергнутых воздействию 40 нм BPEI-AuNP, что указывает на то, что дисфункция митохондрий может быть общим механизмом токсичности AuNP, независимо от заряда поверхности и типов клеток [7,8,9]. Недавнее исследование показало, что цитотоксичность, связанная с митохондриальной дисфункцией, наблюдалась в иммортализованных эпителиальных клетках рака простаты и эпителиальных клетках рака легких в ответ на ингибитор фосфорилирования STAT3, OPB-51602 [40].
В пути апоптоза шесть проапоптотических генов CASP3, CASP7, CASP9, TNFRSF10A, TNFRSF10B и TNFSF10 были активированы, тогда как шесть антиапоптотических генов AKT1, ALB, BCL2, BCL2L1, MCL1 были подавлены и XI 6b, дополнительный файл 1:таблица S3), который коррелировал с дозозависимой цитотоксичностью в клетках C3A (рис. 2a). In DNA damage and repair check point, genes of the checkpoint kinases (CHEK1/2), the DNA excision repair genes (ERCC1/2/3), and the DNA ligase IV (LIG4) were upregulated but other excision repair genes (ERCC5/6, XRCC1/5), the checkpoint kinase (CDKN1A), and protein kinases (PRKDC) genes were downregulated (2- to 19-fold). These results suggested that the 40 nm PEG-AuNP-induced interference with cell cycle and DNA repair system may correlate with an induction of cell death in C3A cells (Fig. 2a, Additional file 1:Table S3). Genes encoding two different heat shock proteins (HSP) (A1A and A1B) were upregulated (10.2- to 14.2-fold) but HSP40 subfamily A, B, and C; HSP90 member 1; and HSP60 were downregulated (2- to 16-fold) (Additional file 1:Table S4).
In oxidative stress and antioxidant response, the 40 nm PEG-AuNP at LC50 value induced antioxidants genes and suppressed pro-oxidants, which was associated with a decrease in ROS/RNS generation being antioxidant itself (Fig. 4b). In antioxidant genes, glutathione peroxidase (GPX) 1, GPX2, GPX4, PRDX1, PRDX2, PRDX6, superoxide dismutase (SOD) 1, and CAT were induced (3.8- to 18.1-fold). In pro-oxidant genes, TXNIP and PPP1R15B were suppressed (4.8- and 17-fold, respectively) (Fig. 6c, Additional file 1:Table S5). This is consistent with a previous study that AuNP displayed oxidative stress-induced cytotoxicity in HepG2 and human hepatocytes, irrespective of size [7, 19].
In phase I metabolism, CYP3A4 and ESD genes were extensively suppressed (7-fold and 12-fold, respectively). Especially, inhibitory effect of 40 nm PEG-AuNP on CYP3A4 expression was correlated with a decrease in CYP3A4 activity (Fig. 5). Recent studies reported that the 40 nm BPEI-AuNP inhibited gene expression of CYP1A2, CYP2C9, and CYP3A4 in human hepatocytes; ESD in HUVEC; and CYP1A1 in HPTC [7,8,9]. Epidemiology study demonstrated that CYP enzymes in liver tissue of HCC patient were substantially inhibited by the tumorigenic process at the molecular and the functional level [41].
Drug Uptake and Efflux Transporter Gene Expression Profiling
The development of multidrug resistance (MDR) by tumor cells is one of the main causes of cancer treatment failures [42, 43]. Integral membrane transporters-mediated decrease in drug uptake and increase in drug efflux including P-glycoprotein (P-gp) and breast cancer resistance protein (BCRP) is one of the major mechanisms of MDR.
Differential gene expression of drug efflux and uptake transporters in C3A cells exposed to 40 nm PEG-AuNP showed that a total of 14 genes of ABC transporters (↓12 and ↑2 genes) and a total of 21 genes of SLC transporters (↓21 and ↑0 genes) were substantially modulated at LC50 value (Figs. 6d and 7, Additional file 1:Table S7). In drug efflux transporters of ABC family, genes of multidrug resistance-associated protein (MRP3/ABCC3), MRP4 (ABCC4), and cholesterol efflux regulated protein (CERP/ABCA1) in basolateral membrane were downregulated (7.2- to 10.4-fold). The genes encoding P-gp (ABCB1), MRP2 (ABCC2), BCRP (ABCG2), and sterolin 2 (ABCG8) in canalicular efflux transporters were also suppressed (8.6- to 13.8-fold). In contrast, multidrug resistance (MDR4/ABCB4) in canalicular membrane and mitochondria ABC transporter (MTABC3/ABCB6) in the outer mitochondrial membrane were highly upregulated (9.8-fold and 5.8-fold, respectively). In drug uptake transporters, genes of copper transporter protein (CTR1/SLC31A1) and to a lesser degree organic anion transporting (OAT7/SLC22A9) were also inhibited (18- fold and 15-fold, respectively). These results support a recent study that the 40 nm BPEI-AuNP downregulate MDR3 in human hepatocytes but upregulates MRP3 in HUVEC indicating surface charge- and cell type-dependent interaction between AuNP and efflux transporters [7, 8]. Epidemiology study exhibited that a high expression of BCRP and a low expression of OCT3 occurred in HCC tumor, which was closely associated with the tumor progression and its size [44]. A previous study exhibited that P-gp inhibitor, verapamil enhanced cytotoxicity of glutathione-AuNP conjugated with doxorubicin in feline fibrosarcoma cell lines by increasing intracellular drug concentration [45]. The current study emphasizes that the mechanisms-derived information on the 40 nm PEG-AuNP identified a separate but still complementary action on mitochondrial fatty acid β-oxidation, TCA cycle and respiratory chain, drug efflux and uptake transporters, as well as CYP3A4 activity in C3A cells (Fig. 7). To the end, this will highlight AuNP interaction with key biological processes and its underlying molecular mechanism in HCC, which may be further implicated in the development of more effective therapeutic target in HCC treatment.

A schematic representation of the basic mechanisms of action of 40 nm PEG-AuNP in HCC treatment. Green bars (an inhibition) and pink triangles (an induction) indicate the 40 nm PEG-AuNP-modified biological markers and pathways. Gene ontology analysis is listed in Additional file 1:Tables S2–S7
To validate gene expression analysis from RT 2 array, the nine genes were selected for real-time PCR. In Additional file 1:Table S1, all nine genes were modulated at LC50 of the 40 nm PEG-AuNP. These transcriptional changes were consistent with those in gene expression analysis with PCR arrays (Fig. 6, Additional file 1:Tables S2–S7).
Conclusions
We have presented that cationic BPEI-, anionic LA-, or neutral PEG-AuNP interaction with human plasma protein corona (PC) caused the changes in D H , PDI, and z-potential of AuNP and further influenced cellular responses in C3A cells. All bare (no PC) 40 and 80 nm AuNP were cytotoxic to C3A cells besides the 80 nm LA-AuNP but PC completely ameliorated their cytotoxicities besides the 80 nm BPEI-AuNP. The 40 nm bare BPEI-AuNP showed the highest cellular uptake followed by the 40 nm PEG-AuNP and then the 40 nm LA-AuNP, whereas PC suppressed AuNP uptake besides the 80 nm PEG-AuNP. The 40 nm BPEI-AuNP caused biphasic responses of oxidative stress (pro- and antioxidant) in C3A cells, whereas the 40 nm PEG-AuNP was antioxidant. CYP3A4 activity was extensively suppressed by all bare AuNP, irrespective of size and surface charges, whereas PC substantially ameliorated its inhibitory effect on enzyme activity besides the 40 and 80 nm PEG-AuNP. Differentially expressed genes at LC50 value of 40 nm PEG-AuNP were mainly involved in mitochondrial fatty acid β-oxidation and to a lesser degree hepatic efflux/uptake transporters. The 40 nm PEG-AuNP inhibited three main enzymes in β-oxidation (acyl-CoA dehydrogenase, enoyl-CoA hydratase, and ketoacyl-CoA thiolase), other enzymes in TCA cycle, and the mitochondrial respiratory chain for ATP production. The 40 nm PEG-AuNP increased the expression of pro-apoptotic genes and decreased anti-apoptotic genes at LC50 ценность. A high level of antioxidants and a low level of pro-oxidants genes were observed in C3A cells exposed to 40 nm PEG-AuNP. In addition, genes of drug efflux and uptake transporters located in both basolateral and canalicular membrane were substantially modulated.
Сокращения
- ANOVA:
-
One-way analysis of variance
- AuNP:
-
Gold nanoparticles bare:no PC
- BPEI:
-
Branched polyethylenimine
- CYP:
-
Cytochrome P450
- D H :
-
Hydrodynamic diameters
- DLS:
-
Dynamic light scattering
- EDTA:
-
Ethylenediaminetetraacetic acid
- EMEM:
-
Eagle’s minimum essential medium
- HCC:
-
Human hepatocellular carcinoma
- HPTC:
-
Human renal proximal tubular cells
- HSD:
-
Tukey’s honest significant difference test
- HUVEC:
-
Human umbilical vein cells
- ICP-MS:
-
Inductively coupled plasma mass spectrometry
- LA:
-
Anionic lipoic acid
- LC50 :
-
Median lethal concentration
- MDR:
-
Multidrug resistance
- NP:
-
Наночастицы
- PBS:
-
Phosphate-buffered saline
- PC:
-
Human plasma protein corona
- PDI:
-
Polydispersity index
- PEG:
-
Neutral polyethylene glycol
- RNAi:
-
RNA interference
- RNS:
-
Reactive nitrogen species
- ROS:
-
Активные формы кислорода
- SO:
-
Superoxide
- ТЕМ:
-
Просвечивающая электронная микроскопия
- TFF:
-
Tangential flow filtration
Наноматериалы
- Обслуживание отеля и его влияние на вашу прибыль
- Золотой нанобиосенсор на основе локализованного поверхностного плазмонного резонанса способен диагностиро…
- Синтез и активность окисления CO 1D смешанного бинарного оксида CeO2-LaO x, нанесенного золотыми катализаторами
- Синтез композита поли (3,4-этилендиокситиофен) / золото / графен в твердом состоянии и его применение для амперо…
- Стимуляция роста клеток SH-SY5Y наночастицами золота, модифицированными 6-меркаптопурином и проникающим в нейро…
- Моноклональное антитело к гепараназе, меченное магнитными золотыми наночастицами, и его последующее примене…
- Влияние фуллерена C60 на взаимодействие дифенил-N- (трихлорацетил) амидофосфата с ДНК in Silico и его цитотоксическ…
- Оценка токсичности наночастиц PEG-PCCL и предварительное исследование его противоопухолевого эффекта от загруз…
- Влияние углеродных нанотрубок и их производных на опухолевые клетки in vitro и биохимические параметры, состав к…
- Оценка активности цитохрома P450 3A4, ингибируемого наночастицами золота, и молекулярных механизмов, лежащих в о…