


Экзосомальная микроРНК-326, полученная из макрофагов M1, подавляет развитие клеток гепатоцеллюлярной карциномы посредством опосредования сигнального пути NF-κB
Аннотация
Накапливающиеся данные показали, что микроРНК (miR), происходящая из экзосом, происходящих из макрофагов M1, может регулировать прогрессирование гепатоцеллюлярной карциномы (HCC). Однако о влиянии miR-326, происходящей из экзосом, происходящих из макрофагов M1, на ГЦК не сообщалось. Следовательно, целью настоящего исследования было изучить механизм экзосомальной miR-326 из макрофагов M1 в регуляции прогрессии клеток HCC. RT-qPCR выявила экспрессию miR-326 в линиях клеток HCC. Экспрессия miR-326 в HCC была изменена трансфекцией, и было обнаружено влияние miR-326 на экспрессию CD206 и NF-κB, пролиферацию клеток, образование колоний, миграцию, апоптоз и инвазию. Впоследствии экзосомы были выделены из макрофагов M1. RT-qPCR идентифицировала экспрессию miR-326 в экзосомах, происходящих из макрофагов M1. Экспрессия miR-326 в экзосомах, происходящих из макрофагов M1, была изменена трансфекцией. Экзосомы, полученные из макрофагов M1, культивировали совместно с клетками HCC, чтобы выяснить их влияние на биологический прогресс клеток HCC. Наконец, были проведены эксперименты in vivo для проверки результатов in vitro. MiR-326 был снижен в клетках HCC и обогащен экзосомами, происходящими из макрофагов M1. Повышающая регуляция miR-326 будет ингибировать пролиферацию клеток HCC, образование колоний, миграцию, инвазию, а также экспрессию CD206 и NF-κB и стимулировать апоптоз, а также ингибировать рост опухолей HCC in vivo , тогда как подавление miR-326 показало противоположные эффекты. Экзосомы, происходящие из макрофагов M1, ингибировали пролиферацию клеток HCC, образование колоний, миграцию, инвазию и экспрессию CD206 и NF-κB и повышали апоптоз, в то время как избыточная экспрессия miR-326 усиливала эффект экзосом, полученных из макрофагов M1, на клетки HCC. Выявлено, что происходящая из макрофагов M1 экзосомная miR-326 подавляет пролиферацию, миграцию и инвазию, а также способствует апоптозу ГЦК за счет подавления экспрессии NF-κB.
Введение
Гепатоцеллюлярная карцинома (ГЦК) - пятая по распространенности злокачественная опухоль в мире и самый распространенный первичный рак печени [1]. Согласно данным Китайского национального онкологического реестра, смертность от первичного рака печени занимает третье место, а по заболеваемости - четвертое среди распространенных злокачественных новообразований [2]. Основными факторами риска ГЦК являются хроническая инфекция вирусом гепатита С и вирусом гепатита В, продукты питания, загрязненные афлатоксином, ожирение, курение, злоупотребление алкоголем и диабет 2 типа [3]. Трансартериальная химиоэмболизация - это общепринятый метод лечения ГЦК на промежуточной стадии, который улучшает выживаемость у большинства пациентов с ГЦК на промежуточной или поздней стадии [4]. В настоящее время диагноз ГЦК во многом зависит от биомаркеров сыворотки и методов визуализации [5]. Пятилетняя выживаемость, связанная с ГЦК, составляет всего около 60%, и в последние годы заболеваемость растет с каждым годом [6]. Учитывая это, поиск точных терапевтических целей является приоритетом при лечении ГЦК.
Макрофаги являются эффекторными клетками и основными регуляторами иммунной системы, выполняют огромные функции по ремоделированию и восстановлению тканей, а также организуют метаболические функции почти во всех тканях in vivo [7]. Выявлено, что макрофаги М1 обладают опухолевым действием и повышают подвижность клеток ГЦК [8]. Экзосома представляет собой дискоидную везикулу диаметром 40–150 нм [9]. Согласно Xu et al., Экзосомальные микроРНК (miRNAs) выполняют функции по пролиферации, инвазивности, метастазированию и лекарственной устойчивости ГЦК посредством модуляции экспрессии генов в клетках-мишенях [10]. Исследование продемонстрировало, что экзосомы, содержащие miR-326, могут быть потенциальной клинической мишенью при рассеянном склерозе [11]. MiRNA могут действовать как онкогены или ингибиторы опухолей, модулируя экспрессию большого количества кодируемых белками генов посредством деградации мРНК и блокировки трансляции в определенной последовательности [12]. В исследовании обсуждалось, что miR-326 подавляет рост клеток HCC посредством нарушения прогрессирования клеточного цикла, а также усиления апоптоза, кроме того, подавляет клеточную инвазию посредством снижения фенотипа эпителиально-мезенхимального перехода [13]. Другое исследование показало, что miR-326 может быть потенциальной терапевтической мишенью для лечения пациентов с HCC [14]. Таким образом, в настоящем исследовании обсуждался механизм экзосомальной miR-326, регулирующей инвазию и миграцию клеток HCC.
Материалы и методы
Этика
Все эксперименты на животных проводились в соответствии с Руководством по уходу и использованию лабораторных животных международных комитетов. Протокол был одобрен Комитетом по использованию институциональных средств ухода за животными Третьей больницы Цзилиньского университета.
Индукция и идентификация макрофагов
Линия моноцитарных клеток человека THP-1 (Kunming Institute of Zoology, CAS, Kunming, China) культивировалась в среде RPMI 1640 (Gibco, CA, США; Thermo Fisher Scientific, MA, США), содержащей 10% инактивированной нагреванием фетальной телячьей сыворотки ( FBS). Клетки THP-1 реагировали с 150 нг / мл форбола 12-миристат 13-ацетата (PMA; P8139, Sigma-Aldrich, SF, CA, USA) и инкубировали в среде RPMI в течение 24 часов для получения макрофагов M0. Затем морфологию клеток до и после индукции наблюдали путем окрашивания по Райту. Клетки THP-1 и индуцированные макрофаги ресуспендировали в 5 мкл PBS, капали на предметное стекло, окрашивали раствором красителя Райта и смешивали с буферным раствором в соотношении 1:2. Окрашенные в течение 10 мин и промытые проточной водой клетки наблюдали под микроскопом. Кроме того, маркеры макрофагов M0 (CD68 и CD206) измеряли с помощью количественной полимеразной цепной реакции с обратной транскрипцией (RT-qPCR). Затем макрофаги были индуцированы в макрофаги M1 посредством инкубации с 20 нг / мл интерферона (IFN) -γ (# 285-IF; R&D Systems, MN, USA) и 10 пг / мл LPS (# 8630; Sigma-Aldrich) в течение 18 лет. час Маркеры макрофагов M1 (IDO1 и IL-12 p35) исследовали с помощью RT-qPCR [15].
Извлечение экзосом
Экзосомы экстрагировали с помощью набора для разделения экзосом (ExoEasy Maxi Kit, Qiagen, Hilden, Германия). Супернатант макрофагов собирали в центрифужную пробирку емкостью 15 мл в асептических условиях и фильтровали через 0,8 мкм фильтровальную пленку. К супернатанту клеток в каждой группе добавляли буфер XBP (1:1), а затем центрифугировали на колонке для аффинного центрифугирования с мембраной exoEasy при 500 g . В клетки добавляли 10 мл буфера XWP и центрифугировали с 3000–5000 g . Колонку для аффинного центрифугирования с мембраной exoEasy заполняли 400 мкл буфера для элюирования XE и центрифугировали при 500 g . Буфер для элюирования переносили в колонку для аффинного центрифугирования с мембраной exoEasy и центрифугировали при 500 g . Буфер для элюирования хранили в течение 24 ч при 4 ° C, а затем использовали для идентификации. Остальное хранили при -80 ° C.
Наблюдение с помощью ТЕА и анализ отслеживания наночастиц (NTA)
Полученные выше экзосомы закапывали в медную сетку мембраны на углеродном носителе и добавляли 2% фосфорновольфрамовой кислоты. Образец наблюдали с помощью просвечивающего электронного микроскопа, и оптимальное изображение было собрано и проанализировано.
Примеси и частицы в PBS удаляли с помощью микропористого фильтра 0,22 мкм. В соответствии с плотностью частиц экзосом отфильтрованный PBS был разбавлен до соответствующей концентрации и обнаружен с помощью детектора наночастиц Nanosight NS300 (Малверн, Вестборо, Массачусетс, США).
После идентификации происходящие из макрофагов экзосомы, трансфицированные ингибитором miR-326 и отрицательный контроль (NC) ингибитора miR-326, миметик miR-326 и миметик miR-326 NC были экстрагированы с помощью набора для разделения экзосом (Invitrogen).
RT-qPCR
Суммарную РНК экстрагировали с помощью Trizol Reagent (Thermo Fisher), и ПЦР в реальном времени выполняли с использованием SYBR-Green PCR Master Mix (Roche) и ABI 7500 Real-Time PCR System (Life Technologies, Grand Island, NY, USA. ). Последовательности праймеров показаны в таблице 1. Количественный анализ проводился с использованием метода 2 - △△ Ct .
Вестерн-блоттинг
Общий белок клеток и экзосом экстрагировали. Концентрацию белка определяли с помощью набора для бицинхониновой кислоты (BCA) (Boster Biological Technology Co. Ltd., Wuhan, Hubei, China). К белку добавляли буфер для образцов и кипятили при 95 ° C, и в каждую лунку загружали 30 мкг. Белок отделяли электрофорезом в 10% полиакриламидном геле (Boster Biological Technology) и наносили электроблоттинг на поливинилиденфторидную мембрану с последующим запечатыванием в 5% бычьем сывороточном альбумине (BSA). Мембрана была засеяна первичными антителами против CD63 (1:1000, Developmental Studies Hybridoma Bank, Университет Айовы, Эймс, Айова, США), CD181 (1:1000, R&D Systems), GAPDH (1:2000, Jackson ImmunoResearch Laboratories, PA, USA) и вторичным антителом (1:500, Jackson ImmunoResearch Laboratories), меченным пероксидазой хрена. Изображения были получены с помощью двухцветной инфракрасной флуоресцентной сканирующей системы визуализации Odyssey, а значения серого в полосах были измерены с помощью программного обеспечения для анализа изображений Quantity One.
Культура клеток и скрининг
Линию нормальных клеток печени HL-7702 и линию клеток HCC человека BEL-7404, HepG2, SMMC-7721 и QGY-7703 отбирали и культивировали в среде Gibco RPMI 1640 с 10% фетальной бычьей сывороткой (FBS), пенициллином (100 Ед / мл. ) и стрептомицин (100 мг / мл). Экспрессия MiR-326 была обнаружена с помощью RT-qPCR, и подходящие клеточные линии были подвергнуты скринингу.
Маркировка экзосом и поглощение экзосом
Экзосомы ресуспендировали в 250 мкл разбавителя C и осторожно растирали, избегая повреждения мембраны экзосомы. Краситель PKH67 (1 мкл, Sigma-Aldrich) добавляли к 250 мкл разбавителя C до 500 мкл и инкубировали. К раствору добавляли 500 мкл 1% BSA и инкубировали при 37 ° C в течение 1 мин. Экзосомы получали центрифугированием при 120 000 g 4 ° C в течение 2 часов. Экзосомы, меченные PKH67, были получены центрифугированием при 120 000 g 4 ° C в течение 2 часов. Экзосомы ресуспендировали в 6 мл среды RPMI-1640, избегая света. Затем меченые экзосомы культивировали совместно с клетками HCC в течение 12 часов. После этого культуральную среду удаляли и промывали PBS 3 раза по 5 мин / раз, а экзосомы с флуоресцентной меткой, которые не абсорбировались внутри клеток HCC, тщательно смывали. Экзосомы закрепляли 4% параформальдегидом и окрашивали 4'-6-диамидино-2-фенилиндолом. После герметизации распределение флуоресценции наблюдали с помощью лазерного конфокального микроскопа.
Группировка и обработка ячеек
Клетки HepG2 и клетки SMMC-7721 высевали в 12-луночный планшет при 0,5–1 × 10 6 ячеек / лунка. При 50–60% конфлюэнции клетки трансфицировали липофектамином 2000 (Invitrogen, Карлсбад, Калифорния). Клетки HepG2 были распределены в группу, имитирующую miR-326 (трансфицированную миметиком miR-326) и группу, имитирующую miR-326 (трансфицированную миметиком miR-326). Клетки SMMC-7721 были отнесены к группе ингибитора miR-326 (трансфицированной ингибитором miR-326) и группе ингибитора NC (трансфицированной ингибитором miR-326 NC). miR-326-миметик, miR-326-ингибитор и их NC смешивали с липофектамином 2000 для трансфекции. Клетки HepG2 и клетки SMMC-7721 без какой-либо обработки были выбраны в качестве пустой группы. miR-326-миметик, miR-326-ингибитор и их NC были разработаны и составлены Guangzhou RibBio Co., Ltd. (Гуанчжоу, Китай) (таблица 1).
Совместное культивирование экзосом, полученных из макрофагов M1, с клетками HCC
Концентрацию белка в суспензии экзосом, полученных из макрофагов M1, определяли методом BCA и рассчитывали объем соответствующей суспензии экзосом с 50 мкг белка. Клетки HepG2 и клетки SMMC-7721 высевали в 12-луночный планшет в количестве 1 × 10 5 . клеток / мл на лунку. Клетки HepG2 были разделены на 4 группы:контрольная группа (клетки HepG2, не культивируемые совместно с экзосомами), группа экзосом (Exo) (клетки HepG2, совместно культивируемые с экзосомами, происходящими из макрофагов M1), группа, имитирующая Exo-miR-326 (HepG2 клетки, совместно культивируемые с экзосомами, происходящими из макрофагов M1, которые трансфицированы миметиком miR-326), группа Exo-NC-миметиков (клетки HepG2, совместно культивируемые с экзосомами, происходящими из макрофагов M1, которые трансфицированы миметиком miR-326). Клетки SMMC-7721 также были разделены на 4 группы:пустая группа (клетки SMMC-7721, не культивируемые совместно с экзосомами), группа Exo (клетки SMMC-7721, совместно культивируемые с экзосомами, происходящими из макрофагов M1), Exo-miR-326- группа ингибиторов (клетки SMMC-7721, совместно культивируемые с экзосомами, происходящими из макрофагов M1, которые трансфицированы ингибитором miR-326), группа ингибиторов Exo-NC (клетки SMMC-7721, совместно культивируемые с экзосомами, происходящими из макрофагов M1, которые трансфицированы miR- 326 ингибитор NC).
Анализ 3- (4, 5-диметилтиазол-2-ил) -2,5-дифенилтетразолия бромида (МТТ)
Клетки отделяли трипсином и высевали на 96-луночный планшет с плотностью клеток 4 × 10 4 . клеток на лунку. Культуральная среда была оставлена после культивирования через 12, 24, 36, 48, 60 часов соответственно. К клеткам, инкубированным с 500 мкл 0,5 г / л раствора МТТ, добавляли 200 мкл раствора диметилсульфоксида, растирали и вылупляли. Значения оптической плотности (OD, 490 нм) измеряли с помощью ридера для микропланшетов.
Анализ образования колоний
Культивировали в течение 24 часов и отделили трипсином, клетки высевали в маленькую чашку диаметром 35 мм по 300 клеток на чашку. Раствор заменяли каждые 3 дня. Через 10 дней культивирования клетки фиксировали 40 г / л -1 . параформальдегид и окрашенный 1 г / л -1 кристаллический фиолетовый раствор и сушат. Количество колоний (более 50 клеток) подсчитывали под микроскопом.
Анализ Transwell
Ячейки (1 × 10 5 ) были суспендированы с 200 мкл пустой культуральной среды. Эксперименты проводились в соответствии с инструкциями камеры Transwell (Corning Glass Works, Corning, N.Y., USA) (матригель требовался для эксперимента по инвазии, но не для эксперимента по миграции). RPIM 1640 (10% FBS, 600 мкл) добавляли в нижнюю камеру. Верхняя и нижняя камеры были разделены мембраной Transwell, предварительно покрытой матригелем (BD Biosciences, Франклин Лейкс, Нью-Джерси, США). После культивирования в течение 24 ч камеру фиксировали 95% спиртом. После окрашивания раствором кристаллического фиолетового клетки наблюдали в пяти полях зрения под микроскопом.
Проточная цитометрия
Клеточный цикл:клетки отделялись трипсином. Ячейки (1 × 10 6 ) суспендировали в 0,5 мл PBS и растирали до единой суспензии. Смешанные с 4,5 мл предварительно охлажденного 70% этанола на льду, клетки центрифугировали при 3000 g , промыли 5 мл PBS и снова центрифугировали при 3000 g . После этого клетки суспендировали в 1 мл окрашивающего раствора PI / Triton X-100 (20 мкг PI / 0,1% Triton X-100), содержащего 0,2 мг РНКазы. Клеточный цикл был определен с помощью проточной цитометрии.
Апоптоз клеток:трипсинизированные клетки (1 × 10 6 ) суспендировали в 1 мл PBS, растирали и центрифугировали при 3000 g . Клетки промывали по очереди инкубационным буфером (10 ммоль / л Hepes / NaOH, pH 7,4, 140 ммоль / л NaCl, 5 ммоль / л CaCl 2 ) и центрифугировали при 3000 g . Затем клетки инкубировали со 100 мкл маркировочного раствора (FITC-Annexin V и PI были добавлены в инкубационный буфер до достижения 1 мкг / мл), центрифугировали при 3000 g , промыли один раз инкубационным буфером и заштриховали флуоресцентным (SA-FLOUS) раствором. Апоптоз клеток определяли методом проточной цитометрии. Длина волны проточной цитометрии составляла 488 нм, флуоресценцию FITC детектировали полосовым фильтром при 515 нм, в то время как PI с длиной волны больше 560 нм. Результаты были автоматически проанализированы компьютером.
Ксенотрансплантат опухоли у голых мышей
Сорок мышей (Колледж зоотехники, Университет Цзилинь, Цзилинь, Китай) в возрасте 4–6 недель были случайным образом распределены в 8 групп, по 5 мышей в каждой группе. Мышей кормили в специальной лаборатории для животных, свободной от патогенов, в течение 1 недели, и корм, подушка и бутыль с водой меняли вовремя. За состоянием здоровья мышей следует наблюдать каждый день. Через неделю из клеток HCC готовили суспензию клеток и вводили подкожно в шею и обратно в концентрации 0,1 мл суспензии клеток (1 × 10 6 ). Рост опухоли наблюдался через 3-5 дней. Голых мышей взвешивали каждые 4 дня и измеряли объем опухоли. Голых мышей умерщвляли через 20 дней после инъекции.
Статистический анализ
Все данные были интерпретированы с помощью программного обеспечения SPSS 17.0 (SPSS Statistics, Чикаго, Иллинойс, США). Данные измерений были указаны как среднее ± стандартное отклонение. Сравнения между двумя группами были сформулированы t -тест, в то время как сравнения между несколькими группами оценивались с помощью одностороннего дисперсионного анализа (ANOVA). P значение <0,05 свидетельствует о статистически значимой разнице.
Результаты
Идентификация макрофагов и экзосом M1
Окрашивание Райта использовали для наблюдения за морфологией клеток THP-1, индуцированной PMA. Было показано, что объем клеток THP-1 до индукции был небольшим, а доля кариоплазмы была выше; морфология клеток после индукции была неправильной, объем увеличивался, а доля кариоплазмы уменьшалась; цитоплазма была богаче и имела светло-голубой цвет, с обильными частицами и несколькими вакуолями; ядро было пурпурно-красным и часто было наклонено в одну сторону, показывая, что клетки имели типичные морфологические характеристики макрофагов (рис. 1а).
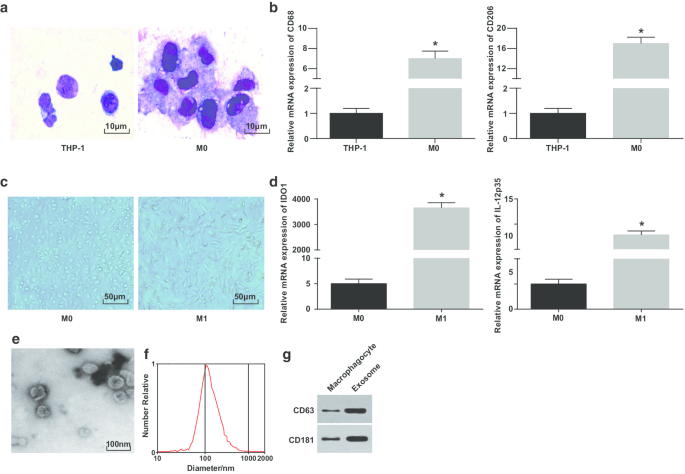
Идентификация макрофагов M1 и экзосом. а Окрашивание Райта для наблюдения за морфологией клеток THP-1 до и после обработки PMA. б Экспрессия CD68 и CD206 в клетках THP-1 и обработанных PMA клетках THP-1, обнаруженная с помощью RT-qPCR; c морфология макрофагов и макрофагов M1. г Экспрессия IDO1 и IL-12 p35 в макрофагах M0 и макрофагах M0, обработанных LPS и INF-γ, обнаруживается с помощью RT-qPCR. е ТЕМ для наблюдения за экзосомами. е Обнаружение гранулометрического состава экзосом с помощью NTA. г Полосы белка CD63 и CD181. На панели b * P <0,05 по сравнению с клетками THP-1; На панели d * P <0,05 по сравнению с макрофагами M0. Данные измерений были представлены как среднее значение ± стандартное отклонение ( N =3), сравнения между двумя группами проводились с помощью t-критерия
Для дополнительной проверки успешной индукции макрофагов экспрессию CD68 и CD206 до и после индукции тестировали с помощью RT-qPCR. Было продемонстрировано, что экспрессия CD68 и CD206 была повышена после индукции PMA, что указывает на то, что PMA успешно индуцировал клетки THP-1 в макрофаги M0 (фиг. 1b). Затем макрофаги M0 были поляризованы в макрофаги M1 путем индукции LPS и INF-γ. Наблюдали и тестировали морфологию и поверхностный маркер макрофагов типа M1 IDO1 и экспрессию IL-12 p35 макрофагов. Макрофаги M0 имели различную и неправильную морфологию прикрепления, показывая округлую, эллиптическую или веретеновую форму. Морфология макрофагов, стимулированных IFN-γ, демонстрирует большее количество ложноножек, выступов и веретенообразных форм (рис. 1c). RT-qPCR показывает (рис. 1d), что после обработки LPS и INF-γ макрофаги M1 показали увеличение своих маркеров (IDO1 и IL-12p35).
Впоследствии экзосомы, полученные из макрофагов, наблюдались с помощью ТЕМ. Было обнаружено, что экзосомы, полученные из макрофагов, были богатыми и имели форму круглой или овальной, с мембранной структурой, однородным размером и меньшим количеством загрязняющих веществ (рис. 1e). NTA показал, что экзосомы с централизованным пиком кривой MODE и гладкой линейной имеют более концентрированный диаметр и меньше примесей (рис. 1f). Вестерн-блоттинг показал, что по сравнению с макрофагами экспрессия специфических маркерных белков CD63 и CD181 повышена в экзосомах, полученных из макрофагов (рис. 1g). Эти результаты показывают, что мы успешно индуцировали дифференцировку моноцитов в макрофаги и поляризовали их в макрофаги M1.
Экзосомы, полученные из макрофагов M1, доставляют miR-326 в клетки HCC и влияют на экспрессию miR-326 в клетках HCC
Чтобы проверить, переносят ли экзосомы, полученные из макрофагов M1, miR-326 в клетки HCC, клетки HepG2 и SMMC-7721 культивировали совместно с экзосомами. Можно было узнать, что большое количество экзосом ассимилировалось клетками HepG2 и SMMC-7721 после 4-часовой трансфекции под флуоресцентным микроскопом (рис. 2a, b).
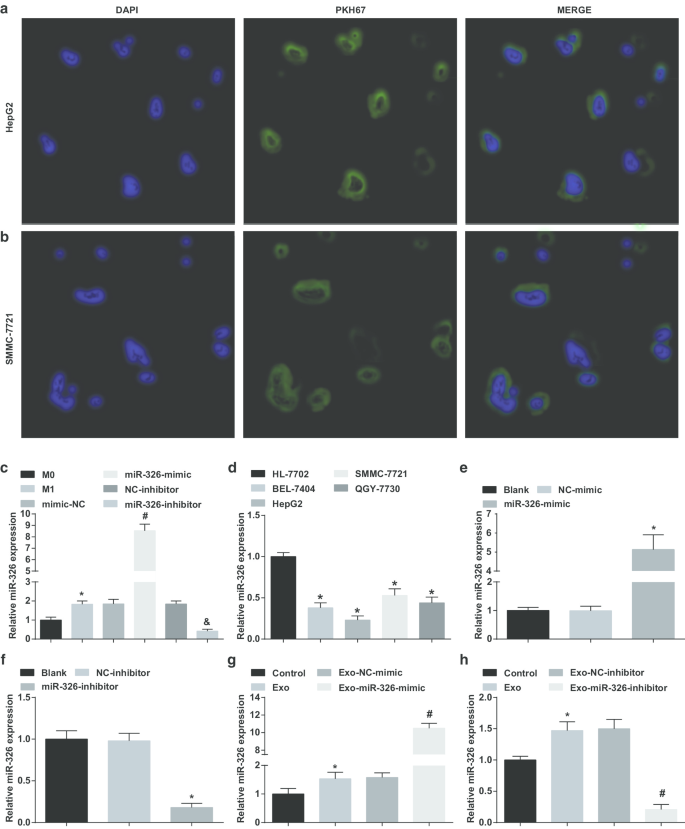
Экзосомы, происходящие из макрофагов M1, доставляют miR-326 в клетки HCC и влияют на экспрессию miR-326 в клетках HCC. а Поглощение экзосом, происходящих от макрофагов M1, клетками HepG2. б Поглощение экзосом, происходящих из макрофагов M1, клетками SMMC-7721. c Сравнение экспрессии miR-326 экзосом макрофагов в каждой группе до и после индукции INF-γ и LPS. г Экспрессия miR-326 в клеточных линиях HCC (BEL-7404, HepG2, SMMC-7721, QGY-7703) и клеточной линии нормальных гепатоцитов человека HL-7702, обнаруженная с помощью RT-qPCR. е RT-qPCR выявила влияние миметика miR-326 на экспрессию miR-326 в клетках HepG2. е RT-qPCR выявила эффект ингибитора miR-326 на экспрессию miR-326 в клетках SMMC-7721. г RT-qPCR выявила влияние экзосом, полученных из макрофагов M1, трансфицированных миметиком miR-326, на экспрессию miR-326 в клетках HepG2. ч RT-qPCR выявила эффекты трансфицированных ингибитором miR-326 экзосом, происходящих из макрофагов M1, на экспрессию miR-326 в клетках SMMC-7721. На панели c * P <0,05 по сравнению с макрофагом M0; На панели d * P <0,05 по сравнению с клетками HL-7702; На панели e * P <0,05 по сравнению с группой NC-миметиков; На панели f * P <0,05 по сравнению с группой ингибиторов NC; На панели g * P <0,05 по сравнению с контрольной группой, # P <0,05 по сравнению с группой имитаторов Exo-NC; На панели h * P <0,05 по сравнению с контрольной группой, # P <0,05 по сравнению с группой ингибиторов Exo-NC. Данные измерений были представлены как среднее значение ± стандартное отклонение ( N =3), сравнения между несколькими группами проводились с помощью одностороннего дисперсионного анализа
Впоследствии миметик / ингибитор miR-326 трансфицировали в макрофаги и проверяли экспрессию miR-326 в их экзосомах до и после трансфекции. Было показано, что по сравнению с экзосомами из макрофагов M0 экспрессия miR-326 повышалась в экзосомах, происходящих из макрофагов M1. Повышенная экспрессия miR-326 наблюдалась в экзосомах, происходящих из макрофагов M1, с обработкой, имитирующей miR-326. Экспрессия MiR-326 снижена в экзосомах, происходящих из макрофагов M1, при лечении ингибитором miR-326 (рис. 2c).
Затем была протестирована экспрессия miR-326 в линиях клеток HCC. Как показано, экспрессия miR-326 была снижена в клетках BEL-7404, HepG2, SMMC-7721 и QGY-7703 по сравнению с клетками HL-7702, среди которых клетки HepG2 проявлялись с наименьшей экспрессией, а клетки SMMC-7721 с наибольшей экспрессией. (Рис. 2d).
После этого миметик miR-326 и ингибитор miR-326 были трансфицированы в клетки HepG2 и SSMC-7721, соответственно, и исследовали их влияние на экспрессию miR-326. MiR-326 имитирует повышенную экспрессию miR-326 в клетках HepG2, тогда как ингибитор miR-326 снижает экспрессию miR-326 в клетках SSMC-7721 (рис. 2e, f).
Затем экзосомы из макрофагов M1, трансфицированных миметиком miR-326 и ингибитором miR-326, культивировали совместно с клетками HepG2 и SSMC-7721 соответственно. Было подчеркнуто, что совместное культивирование с экзосомами увеличивает экспрессию miR-326 в клетках HCC, экзосомы, полученные из макрофагов M1, трансфицированные miR-326, дополнительно повышают экспрессию miR-326 в клетках HepG2, в то время как макрофаги M1, трансфицированные ингибитором miR-326. -производные экзосомы снижали экспрессию miR-326 в клетках SSMC-7721 (рис. 2g, h). Предполагается, что экзосомы, происходящие из макрофагов M1, доставляют miR-326 в клетки HCC и влияют на экспрессию miR-326 в клетках HCC.
Экзосомный miR-326, полученный из макрофагов M1, снижает способность к пролиферации и образованию колоний в клетках HCC
При изучении влияния экзосомальной miR-326 на пролиферацию клеток HCC, МТТ и анализы образования колоний были проведены для изучения пролиферации клеток HCC. Было высказано предположение, что в клетках HepG2 восстановление miR-326 нарушает пролиферацию клеток и способность образовывать колонии (рис. 3a, e, f). Экзосомы, происходящие из макрофагов M1, препятствовали пролиферации клеток HepG2 и образованию колоний. Экзосомы, имитирующие трансфекцию MiR-326 из макрофагов M1, дополнительно нарушали способность к пролиферации клеток и образованию колоний (рис. 3c, i, j).
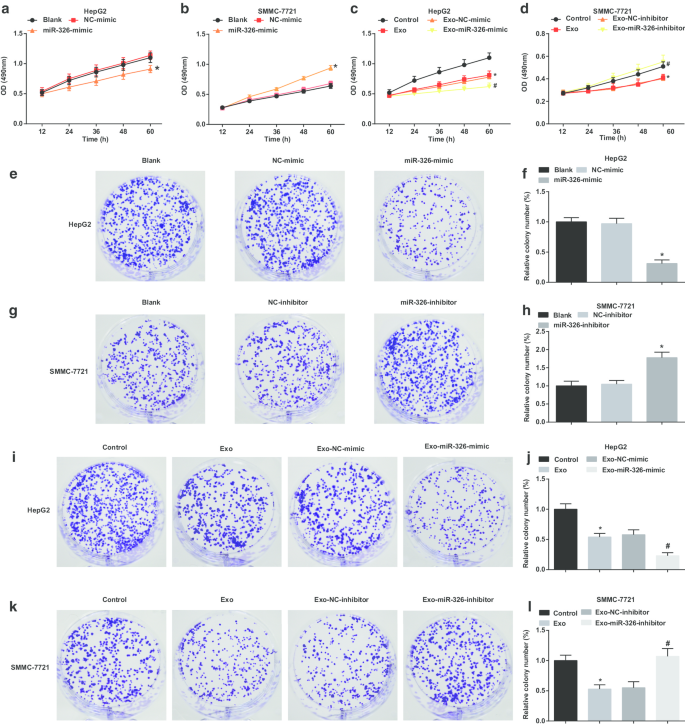
Экзосомный miR-326 макрофага M1 ингибирует пролиферацию клеток и образование колоний в клетках HCC. а Анализ МТТ выявил эффект трансфекции миметика miR-326 на пролиферацию клеток HepG2. б Анализ МТТ выявил эффект трансфекции ингибитора miR-326 на пролиферацию клеток SMMC-7721. c Анализ МТТ выявил влияние совместного культивирования с экзосомами, полученными из макрофагов M1, трансфицированных миметиком miR-326, на пролиферацию клеток HepG2. г Анализ МТТ выявил влияние совместного культивирования с экзосомами, происходящими из макрофагов M1, трансфицированных ингибитором miR-326, на пролиферацию клеток SMMC-7721. е Анализ образования колоний выявил влияние трансфекции миметика miR-326 на способность клеток HepG2 образовывать колонию. е Колонии клеток HepG2. г Анализ образования колоний выявил влияние трансфекции ингибитора miR-326 на способность клеток SMMC-7721 образовывать колонии. ч Численность колоний клеток SMMC-7721. я Анализ образования колоний выявил влияние совместного культивирования с экзосомами, полученными из макрофагов M1, трансфицированных миметиком miR-326, на способность клеток HepG2 образовывать колонии. j Количество колоний клеток HepG2, обработанных экзосомами. к Анализ образования колоний выявил влияние совместного культивирования экзосом, полученных из макрофагов M1, трансфицированных ингибитором miR-326, на способность клеток SMMC-7721 образовывать колонии. l Количество колоний клеток SMMC-7721, обработанных экзосомами. На панелях a и f * P <0,05 по сравнению с группой NC-миметиков; На панелях b и h * P <0,05 по сравнению с группой ингибиторов NC; На панелях c и j * P <0,05 по сравнению с контрольной группой, # P <0,05 по сравнению с группой имитаторов Exo-NC; На панелях d и l * P <0,05 по сравнению с контрольной группой, # P <0,05 по сравнению с группой ингибиторов Exo-NC. Данные измерений были представлены как среднее значение ± стандартное отклонение ( N =3), сравнения между несколькими группами проводились с помощью одностороннего дисперсионного анализа
В клетках SMMC-7721 нокдаун miR-326 усиливал пролиферацию клеток и способность образовывать колонии (рис. 3b, g, h). В клетках SMMC-7721, обработанных экзосомами, происходящими из макрофагов M1, способность к пролиферации и колониеобразованию уменьшалась. Экзосомы, полученные из макрофагов M1, трансфицированные ингибитором MiR-326, дополнительно способствовали пролиферации клеток и образованию колоний (рис. 3d, k, l). Предполагается, что miR-326, полученная из экзосом, происходящих из макрофагов M1, препятствует пролиферации клеток HCC.
Экзосомный белок miR-326, полученный из макрофагов M1, подавляет миграцию и вторжение клеток HCC
Затем исследовали влияние экзосомальной miR-326 на инвазию и миграцию клеток HCC. Было продемонстрировано, что в клетках HepG2 восстановление miR-326 ограничивает инвазию и миграцию (Fig. 4a-c). Экзосомы, происходящие из макрофагов M1, мешали клеткам HepG2 проникать и мигрировать. Инвазия и миграция еще больше ухудшались, когда клетки HepG2 совместно культивировали с экзосомами, полученными из макрофагов M1, трансфицированных miR-326 (рис. 4g – i).
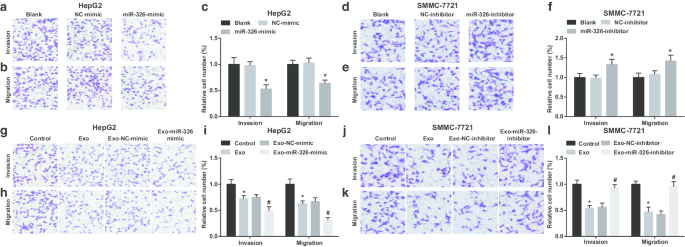
Экзосомный miR-326 макрофага M1 ингибирует миграцию и инвазию клеток HCC. а Анализ Transwell обнаружил эффект трансфекции миметика miR-326 на инвазию клеток HepG2. б Анализ Transwell обнаружил эффект трансфекции miR-326, имитирующий миграцию клеток HepG2. c Количество инвазий и миграций клеток HepG2. г Анализ Transwell выявил влияние трансфекции ингибитора miR-326 на инвазию клеток SMMC-7721. е Анализ Transwell выявил эффект трансфекции ингибитора miR-326 на миграцию клеток SMMC-7721. е Количество инвазий и миграций клеток SMMC-7721. г Анализ Transwell выявил влияние совместного культивирования с экзосомами, полученными из макрофагов M1, трансфицированных миметиком miR-326, на инвазию клеток HepG2. ч Анализ Transwell выявил влияние совместного культивирования с экзосомами, полученными из макрофагов M1, трансфицированных миметиком miR-326, на миграцию клеток HepG2. я Число инвазий и миграций клеток HepG2, совместно культивируемых с экзосомами, полученными из макрофагов M1. j Анализ Transwell выявил влияние совместного культивирования с экзосомами, происходящими из макрофагов M1, трансфицированных ингибитором miR-326, на инвазию клеток SMMC-7721. к Анализ Transwell выявил влияние совместного культивирования с экзосомами, происходящими из макрофагов M1, трансфицированных ингибитором miR-326, на миграцию клеток SMMC-7721. l Число инвазий и миграций клеток SMMC-7721, совместно культивируемых с экзосомами, полученными из макрофагов M1. На панели c * P <0,05 по сравнению с группой NC-миметиков; На панели f * P <0,05 по сравнению с группой ингибиторов NC; На панели i * P <0,05 по сравнению с контрольной группой, # P <0,05 по сравнению с группой имитаторов Exo-NC; На панели l * P <0,05 по сравнению с контрольной группой, # P <0,05 по сравнению с группой ингибиторов Exo-NC. Данные измерений были представлены как среднее значение ± стандартное отклонение ( N =3), сравнения между несколькими группами проводились с помощью одностороннего дисперсионного анализа
MiR-326 knockdown resulted in enhancements in SMMC-7721 cell invasion and migration (Fig. 4d–f). When treated with M1 macrophages-derived exosomes, SMMC-7721 cells were exhibited with decreased invasion and migration. However, SMMC-7721 cell invasion and migration were boosted upon co-culture with miR-326-inhibitor-transfected M1 macrophages-derived exosomes (Fig. 4j–l). It is implied that miR-326 derived from M1 macrophage exosomes impedes the invasion and migration of HCC cells.
M1 Macrophage-Derived Exosomal miR-326 Promotes Apoptosis of HCC Cells
When examining the effect of exosomal miR-326 on the cell cycle and apoptosis of HCC cells, PI single staining and Annexin V-FITC/PI double staining were applied. It was illustrated that miR-326 overexpression increased cells arrested at G0/G1 phase, reduced cells arrested at S and G2/M phases and raised apoptosis in HepG2 cells (Fig. 5a–d). Co-culturing with M1 macrophages-derived exosomes increased cells arrested at G0/G1 phase, reduced cells arrested at S and G2/M phases and raised cell apoptosis of HepG2 cells. Co-cultivation with exosomes from M1 macrophages transfected with miR-326-mimic further enhanced these effects (Fig. 5i–l).

M1 macrophage exosomal miR-326 promotes apoptosis of HCC cells. а , b Flow cytometry detected the effect of transfection of miR-326 mimic on HepG2 cell cycle; c , d flow cytometry detected the effect of transfection of miR-326 mimic on HepG2 cell apoptosis; е , f flow cytometry detected the effect of transfection of miR-326 inhibitor on SMMC-7721 cell cycle; г , ч flow cytometry detected the effect of transfection of miR-326 inhibitor on SMMC-7721 cell apoptosis; i , j flow cytometry detected the effects of co-culture of with miR-326 mimic-transfected M1 macrophage-derived exosomes on HepG2 cell cycle; к , l flow cytometry detected the effect of co-culture with miR-326 mimic-transfected M1 macrophage-derived exosomes on HepG2 cell apoptosis; m , n flow cytometry detected the effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on SMMC-7721 cell cycle; o , p , flow cytometry detected the effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on SMMC-7721 cell apoptosis. In panel b and d, *P <0.05 versus the NC-mimic group; In panel f and h, *P <0.05 versus the NC-inhibitor group; In panel j and l, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-mimic group; In panel n and p, *P <0.05 versus the controlgroup, #P <0.05 versus the Exo-NC-inhibitor group. Measurement data were depicted as mean ± standard deviation (N = 3), comparisons among multiple groups were conducted by one-way analysis of variance
In SMMC-7721 cells, miR-326 down-regulation reduced cells arrested at G0/G1 phase, elevated cells arrested at S and G2/M phases, and declined cell apoptosis (Fig. 5e–h). Untransfected M1 macrophages-derived exosomes increased cells arrested at G0/G1 phase, reduced cells arrested at S and G2/M phases, and heightened cell apoptosis. MiR-326-inhibitor-transfected M1 macrophages-derived exosomes degraded cells arrested at G0/G1 phase, elevated cells arrested at S and G2/M phases, and decreased cell apoptosis (Fig. 5m–p). Briefly, it is summarized that miR-326 derived from M1 macrophage exosomes arrests cell cycle in G0/G1 phase and induces cell apoptosis in HCC.
M1 Macrophage-Derived Exosomal miR-326 Declines CD206 and NF-κB Expression in HCC Cells
Next, the potential mechanism of miR-326 derived from M1 macrophage exosomes in the biological progress of HCC cells was explored. NF-κB is the key link between inflammation and cancer. Many regulatory proteins and miRNAs could inhibit the excessively activated NF-κB signaling to suppress cancer [16]. Such a beneficial effect may include the polarization of M2 macrophages into M1 macrophages. CD206 and NF-κB expression in HepG2 and SMMC-7721 cells was tested by RT-qPCR. It was suggested that miR-326 restoration decreased CD206 and NF-κB expression in HepG2 cells, while miR-326 knockdown enhanced CD206 and NF-κB expression in SMMC-7721 cells (Fig. 6a, b). Moreover, co-culture with M1 macrophage exosomes significantly reduced CD206 and NF-κB expression in HepG2 cells, while co-culture with M1 macrophage exosomes-overexpressing miR-326 further decreased CD206 and NF-κB expression. Treated with untransfected M1 macrophages-derived exosomes, CD206 and NF-κB expression was decreased in SMMC-7721 cells. Co-cultured with miR-326-inhibitor-transfected M1 macrophages-derived exosomes, SMMC-7721 cells were featured by heightened CD206 and NF-κB expression (Fig. 6c, d). It was concluded that miR-326 from M1 macrophage exosomes played a tumor suppressor by inhibiting NF-κB in HCC cells.

M1 macrophage exosomal miR-326 declines CD206 and NF-κB expression in HCC cells. а RT-qPCR detected the effect of transfection of miR-326 mimic on the expression of CD206 and NF-κB in HepG2 cells; б RT-qPCR detected the effect of transfection of miR-326 inhibitor on the expression of CD206 and NF-κB in SMMC-7721 cells. c RT-qPCR detected the effect of co-culture with miR-326 mimic-transfected M1 macrophage-derived exosomes on the expression of CD206 and NF-κB in HepG2 cells; г RT-qPCR detected the effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on the expression of CD206 and NF-κB in SMMC-7721 cells. In panel a, *P <0.05 versus the NC-mimic group; In panel b, *P <0.05 versus the NC-inhibitor group; In panel c, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-mimic group; In panel d, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-inhibitor group. Measurement data were depicted as mean ± standard deviation (N = 3), comparisons among multiple groups were conducted by one-way analysis of variance
miR-326 from M1 Macrophage Exosomes Inhibits HCC Tumor Growth In Vivo
Finally, the in vivo results were validated through tumor xenografts. As displayed, miR-326 overexpression decreased volume and weight of tumors in HepG2 cells (Fig. 7a–c). In mice transplanted with HepG2 cells co-cultured with exosomes, the treatment with M1 macrophage exosomes significantly reduced the tumor volume and weight of HepG2 cells, while treatment with M1 macrophages-overexpressing miR-326 further reduced the tumor volume and weight of HepG2 cells (Fig. 7g–i).

M1 macrophage exosomal miR-326 reduces the volume and weight of HCC tumor in vivo. а - c The effect of transfection of miR-326 mimic on the tumor volume and tumor of nude mice xenografted with HepG2 cells. г - е The effect of transfection of miR-326 inhibitor on the tumor volume and tumors of nude mice xenografted with SMMC-7721 cells. г - я The effect of co-culture with miR-326 mimic-transfected M1 macrophage-derived exosomes on the tumor volume and tumor of nude mice xenografted with HepG2 cells. j –l The effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on the tumor volume and tumor of nude mice xenografted with SMMC-7721 cells. In panel b and c, *P <0.05 versus the NC-mimic group; In panel e and f, *P <0.05 versus the NC-inhibitor group; In panel h and i, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-mimic group; In panel k and l, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-inhibitor group. Measurement data were depicted as mean ± standard deviation (n = 5), comparisons among multiple groups were conducted by one-way analysis of variance
In SMMC-7721 cells, miR-326 suppression increased volume and weight of tumors (Fig. 7d–f). Untransfected M1 macrophages-derived exosomes obstructed tumor growth in volume and weight of SMMC-7721 cells. Co-cultured with miR-326 inhibitor-transfected M1 macrophages-derived exosomes, SMMC-7721 cells were injected into mice and caused elevations in tumor volume and weight (Fig. 7j–l). Summarily, miR-326 derived from M1 macrophage exosomes depressed tumor growth of HCC in vivo.
Discussion
HCC is a common cancer that is characterized with high morbidity and mortality, difficult early diagnosis and treatment, poor prognosis and 5-year survival rate [17]. Recently, a study has highlighted that the lowly expressed lncRNA cox-2 declines the ability of M1 macrophages to suppress HCC cell growth, invasiveness, angiogenesis migration and promote apoptosis [18]. Hu et al. have discussed that miR-326 is obviously degraded in HCC tissues and cell lines, while down-regulated miR-326 is connected to the TNM stage, lymph node metastasis and differentiation of HCC patients [14]. It is customarily considered that HCC cells-derived exosomes can form a fertile environment to facilitate HCC cells growth, invasiveness and metastasis as well as development of drug resistance [19]. The current study was designed to explore the mechanism of exosomal miR-326 in regulating invasion and migration of HCC cells.
Our results indicated that miR-326 expression decreased in HCC cells but increased in exosomes. A recent study has pointed out that miR-326 expression is declined in HCC tissues [20]. Another study has presented miR-326 expression is notably reduced in HCC cell lines and tissues and its down-regulation predicts a poor prognosis in HCC [13]. It is reported that miR-326 acts a tumor-suppression role and is greatly depressed in HCC cells [21]. All these aforementioned evidences are in line with our findings. A study has purported that in comparison with the controls, miR-326 expression is raised in Tconv-derived exosomes which is observed in relapsing–remitting multiple sclerosis patients [11].
Other results emerge from our data that highly expressed exosomal miR-326 reduced cell proliferation, colony formation, migration and invasion as well as facilitated apoptosis of HCC cells in vitro and reduced the volume and weight of HCC tumor in vivo. It has been suggested previously that HCC cell growth can be suppressed via overexpression of miR-326, and HCC cell migration and invasion ability are markedly attenuated through elevating miR-326 [21]. It is reported that the up-regulated miR-326 expression suppresses HCC cell growth and invasiveness as well as stimulates cell apoptosis in vitro [14]. Besides that, a prior study has verified that overexpression of miR-326 declined tumor growth in vivo [13]. A study has revealed that ectopic expressed miR-326 markedly attenuates cell growth, and suppresses cellular migration and invasiveness in non-small cell lung cancer cell lines [22]. Moreover, it is found that miR-326 decreases profibrotic genes like MMP-9, implying its repressive function in cancer cell proliferation [23]. Also, it is presented that miR-326 represses Bcl-2 protein expression and elevates Bax expression so as to affect the apoptosis [24]. Similar to our findings, there are some miRNAs interacting with exosomes to play a role in HCC development. It is displayed that highly expressed exosomal miR-638 can repress the proliferation of HCC cells, involving the potential impact on carcinogenesis [25]. Another study also proves that HCC cells-derived exosomal miR-451a suppresses tumor angiogenesis via disrupting endothelial functions as apoptosis, tube formation, migration and permeability [26]. A prior research generally confirms that when treated with the overexpression of miR-744 exosomes, the proliferation of HCC cells is dramatically suppressed [27].
Conclusion
To briefly conclude, our study provides evidence that M1 macrophage-derived exosomal miR-326 suppresses proliferation, migration and invasion as well as advances apoptosis of HCC cells, supplying a new insight in a novel target therapy for HCC. Due to the limited sample size and limited known researches, the exact mechanism of miR-326 is not fully elucidated, and therefore, further large-scale studies are required to illustrate the underlying mechanism.
Сокращения
- HCC:
-
Hepatocellular carcinoma
- miRNAs:
-
MicroRNAs
- FBS:
-
Fetal bovine serum
- M/CSF:
-
Macrophage colony-stimulating factor
- PBS:
-
Phosphate buffered saline
- DMEM:
-
Dulbecco’s modified eagle medium
- PPAR:
-
Peroxisome proliferator-activated receptor
- ТЕМ:
-
Просвечивающий электронный микроскоп
- NTA:
-
Nanoparticle tracking analysis
- NC:
-
Negative control
- RT-qPCR:
-
Reverse transcription quantitative polymerase chain reaction
- BCA:
-
Bicinchoninic acid
- BSA:
-
Бычий сывороточный альбумин
- Exo:
-
Exosomes
- ANOVA:
-
Односторонний дисперсионный анализ
Наноматериалы
- Полный контроль терагерцовой поляризации с расширенной полосой пропускания через диэлектрические метапове…
- Изучение in vitro влияния наночастиц Au на клеточные линии HT29 и SPEV
- Эффект распыленного под косым углом электрода ITO в структурах перовскитных солнечных элементов MAPbI3
- Изготовление и определение характеристик нового композитного катализатора из углеродного нановолокна Tio2 дл…
- Повышение противоопухолевой эффективности и фармакокинетики буфалина с помощью пегилированных липосом
- Настройка режима скольжения-качения углеродных нанотрубок с помощью гидроксильных групп
- Оценка активности цитохрома P450 3A4, ингибируемого наночастицами золота, и молекулярных механизмов, лежащих в о…
- Крошечные наночастицы фторида редкоземельных элементов активируют рост опухолевых клеток за счет электриче…
- Механизм отказа переключения в программируемой ячейке металлизации на основе пероксида цинка
- Комбинированные нанолипосомы на основе ингибитора аутофагии (LY294002) и 5-фторурацила (5-FU) для повышения эффекти…