


Крошечные наночастицы фторида редкоземельных элементов активируют рост опухолевых клеток за счет электрических полярных взаимодействий
Аннотация
Локализованные внеклеточные взаимодействия между наночастицами и рецепторами трансмембранных сигналов вполне могут активировать рост раковых клеток. Здесь крошечный LaF 3 и PrF 3 наночастицы в суспензиях DMEM + FBS стимулировали рост опухолевых клеток в трех различных линиях клеток человека (A549, SW837 и MCF7). Распределение наночастиц по размерам, активация сигнальных путей AKT и ERK и тесты на жизнеспособность указали на механическую стимуляцию сайтов связывания лигандов интегринов и EGFR посредством синергетического действия ансамбля наночастиц крошечного размера (<10 нм). Хотя наночастицы крошечного размера могут быть хорошо связаны с активацией EGFR, взаимодействие интегрина с наночастицами остается многогранной проблемой. Теоретический мотив показывает, что в пределах необходимого масштаба pN-силы каждый сайт связывания адгезии лиганда может быть активирован диэлектрической наночастицей крошечного размера посредством электрического дипольного взаимодействия. Размер активной наночастицы оставался определяемым количеством поверхностных зарядов на сайте связывания лиганда и наночастице, а также разделяющим расстоянием между ними. Полярная составляющая электрической дипольной силы оставалась обратно пропорциональной второй степени размера наночастиц, показывая, что только диэлектрические наночастицы крошечного размера могут стимулировать рост раковых клеток посредством электрических дипольных взаимодействий. Работа способствует распознаванию различных стрессовых режимов цитоскелета раковых клеток.
Фон
Онкогенез - это многомерная проблема, связанная с геномными изменениями. Он также активируется взаимодействиями клетка-внеклеточный матрикс (ECM) между каркасами и структурами цитоскелета [1,2,3,4], выраженными посредством стресса механосенсоров, подобных интегринам, от многоклеточных сил, способных изменять геномное программирование [5]. Взаимодействие микроокружения опухоли с каркасами ECM обычно активирует белки фокальной адгезии клеточной мембраны и рецепторы трансмембранных сигналов (TSR), рецепторы эпидермального фактора роста (EGFR), фактор роста эндотелия сосудов (VEGFR) или рецепторы фактора роста нервов (NGFR). Механосенсоры регулируют рост опухолевых клеток посредством передачи сигналов между внеклеточным активным доменом клеток [6,7,8,9] и внутриклеточными филаментами F-актина, вызывая лавину реакций фосфорилирования.
Конформационные изменения белков и возбуждение путей TSR требует, чтобы активирующая сила лежала в диапазоне сил pN и, конечно же, ниже уровня nN [10]. Помимо случайного механического воздействия и силы активного химического сродства, эффективность связывания (сила связывания) между наночастицами (НЧ) и белками клеточной мембраны может модулироваться посредством коротких или дальнодействующих электрических полярных или других типов дисперсионных взаимодействий. На ограниченной площади поверхности НЧ только определенное количество белков может быть прикреплено на достаточно долгое время, чтобы оставаться биологически активными [11], и локальные взаимодействия в ограниченном пространстве с биологической средой были признаны ответственными за набор различных функциональных возможностей клеток. маршруты [12]. Следовательно, пути передачи сигналов от взаимодействия белок-NP сигнализируют о проблемах безопасности для NP [11, 13].
Поскольку благоприятный или неблагоприятный ответ в клетках от НЧ зависит от типа [11], связь между НЧ и биологическими метками должна устанавливаться в каждом конкретном случае [14, 15].
Противоречивые результаты воздействия НЧ на опухолевые клетки в отношении абляционной эффективности или эффективности роста опухоли или различных уровней токсичности НЧ [16, 17] также вызывают проблемы безопасности. Тем не менее, несмотря на прогресс, сегодня отсутствует информация о конкретных путях, с помощью которых НЧ взаимодействуют с эукариотическими клетками, что препятствует идентификации универсального терапевтического подхода к НЧ. Поскольку НЧ разного размера и разный химический состав поверхности обычно отклоняют клеточные ответы, включая связывание НЧ с мембранным рецептором и активность TSR, токсичность НЧ связана с морфологией поверхностно-активных веществ, состоянием электрического заряда, концентрацией и составом белков и наноматериалов в ECM. [18,19,20,21] и, наконец, прочность молекулярной связи между НЧ и фенотипами клеток [22].
Предыдущие исследования меланомы и карциномы шейки матки, подвергнутые воздействию кремнезема, золотых НЧ и углеродных нанотрубок, показали, что размер НЧ избирательно активирует рост опухолевых клеток [23,24,25,26,27]. Корреляция между сигналами TSR в линии раковых клеток человека SK-BR-3 и размером модифицированных НЧ золота и серебра продемонстрировала, что, хотя НЧ размером 2–100 нм реформировали передачу сигнала, огромная разница в апоптотической активности была достигнута, когда клетки взаимодействовали с 40 НЧ размером –50 нм [26]. Недавно было также высказано предположение, что изменение размера НЧ золота с 5 до 40 нм, возможно, регулирует скорость роста линий раковых клеток A549 и 95D. В частности, НЧ размером 5 нм подавляли любую пролиферацию обоих типов клеток, в то время как НЧ размером ~ 10 нм не влияли на рост клеток [27]. Аналогичным образом, клетки A549 и THP-1, подвергшиеся воздействию SiO 2 НЧ проявляли цитотоксичность, зависящую от размера, а также НЧ размером 15 нм также коррелировали с высокими уровнями цитотоксичности. Напротив, НЧ размером 60 нм обладают меньшей токсичностью. Наконец, НЧ размером 200 нм увеличивают рост стволовых клеток за счет активации ERK1 / 2, тогда как НЧ размером 2–4 мкм способны активировать различные пути передачи сигнала [28]. НЧ небольшого размера конъюгируют EGFR и включают пути передачи сигналов протеинкиназы B (AKT) и внеклеточной сигнальной киназы (ERK), которые вызывают воспаление клеток.
Наночастицы редкоземельных элементов (RE-NP) могут также взаимодействовать со специфическими доменами, такими как сайты связывания, зависящие от ионов металлов (MIDAS), приспособления к MIDAS (ADMIDAS), сайты связывания синергических ионов металлов (SyMBS) и сайты связывания лигандов (LABS). , расположенный в α ν β 3 субъединица или другие субъединицы интегринов [29, 30].
Точно так же RE-NPs влекут за собой дополнительную степень гибкости во взаимодействии опухолевых NPs [31,32,33,34]. В то время как НЧ церия (наноцерия) проявляют защитное действие против клеточных повреждений различными радикалами [35], низкие уровни концентрации модифицированных НЧ церия значительно ускоряют пролиферацию клеток гепатомы за счет снижения апоптоза за счет активации сигнальных путей AKT / ERK [36]. Обычно ансамбль НЧ, окружающий клетку, ответственен за цитоскелетное напряжение и в равной степени выдерживает химическое, нанотермодинамическое (Хилл) [37], энтропийное или электрическое дипольное взаимодействие между НЧ и механосенсорами. Однако до сих пор понимание взаимодействий между NP, TSR и клетками остается расплывчатым и недоступным.
В принципе, сильный ионный характер соединений RE должен стимулировать механосенсоры клеток посредством электрических взаимодействий. Кроме того, поскольку ионы РЗЭ широко используются в различных приложениях, очень важно изучить их потенциальный вклад в рост опухолевых клеток, что подразумевает соответствующие протоколы защиты общественного здоровья. Фторид лантана (LaF 3 ) и фторид празеодима (PrF 3 ) используются в люминесцентных лампах, радиационных цветных очках, волоконной оптике, эмалевых аппликациях и электродах. ЛаФ 3 разрабатывается в конкретных типах стекол, покрытий люминофорных ламп, водоподготовке и катализаторах. Он также является важным компонентом коммерческого фторидного стекла (ZBLAN), которое в смеси с фторидом европия используется для оптической связи и в качестве кристаллической мембраны в ионоселективных фторидных электродах с хорошим пропусканием в инфракрасном диапазоне. В равной степени PrF 3 также используется в угольных дуговых лампах для киноиндустрии, студийного освещения и прожекторов. Фторидные стекла, легированные празеодимом, также используются в одномодовых волоконно-оптических усилителях.
Таким образом, эта работа демонстрирует, что RE-NP крошечного размера обладают способностью стимулировать рост опухолевых клеток посредством электрических дипольных взаимодействий.
Статья состоит из трех разделов. Во-первых, распределение по размерам, взаимодействия и геометрия наночастиц анализируются с помощью динамического рассеяния света (DLS), атомно-силовой микроскопии (AFM), просвечивающей электронной микроскопии (TEM), дифракции рентгеновских лучей (XRD), двумерной быстрой микроскопии. Анализ с преобразованием Фурье (2D-FFT) и вакуумная ультрафиолетовая спектроскопия (VUV 110–180 нм). Далее, корреляция между уровнем роста трех различных линий раковых клеток человека (A549, SW837 и MCF7) с распределением по размерам и концентрацией LaF 3 и PrF 3 Создана НП. Наконец, в пределах необходимого предела силы в 1 пН для активации механосенсоров и последующего роста опухолевых клеток, жизнеспособность опухолевых клеток укладывается в теоретический мотив электрического дипольного взаимодействия между одним RE-NP и одним LABS. Работа способствует идентификации и классификации различных типов стресса цитоскелета и взаимодействий между НЧ и механосенсорными клетками раковых клеток.
Результаты
Размер и структура НП
Во-первых, DLS, AFM, TEM, XRD, FFT, VUV-спектроскопия и t статистику испытаний применяли для извлечения распределения по размерам RE-NP в жидких суспензиях (рис. 1, 2, 3, 4, 5, 6 и 7). Затем были использованы тесты жизнеспособности клеток и тесты вестерн-блоттинга (Wb) для определения активации конкретных механосенсоров RE-NP.
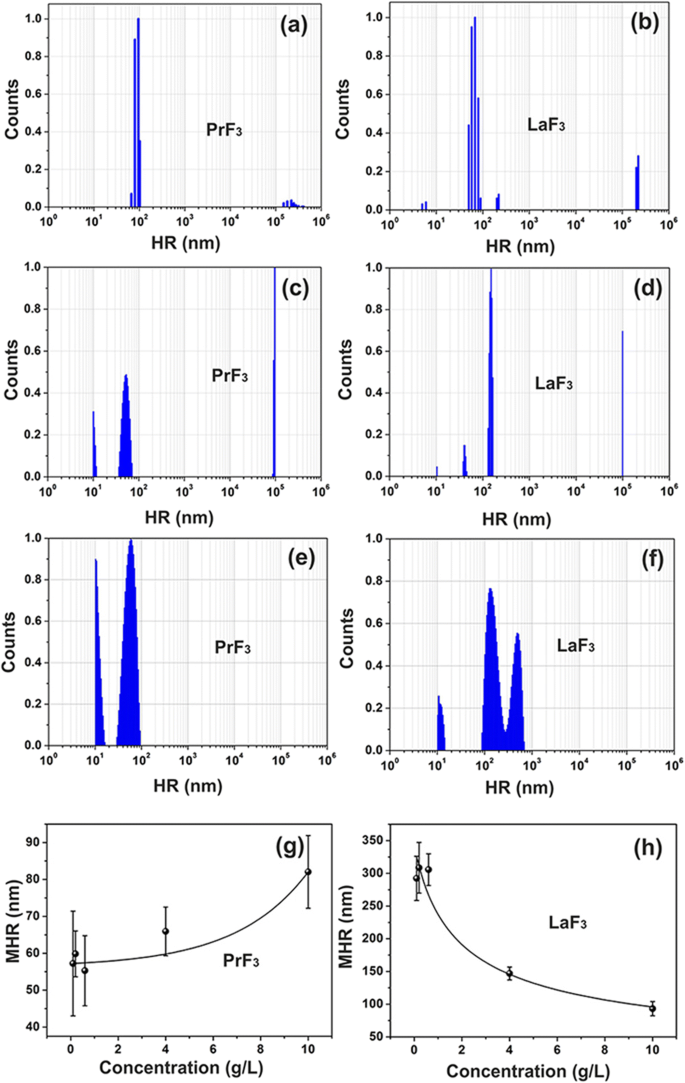
Спектры DLS-распределения суспензий РЗЭ по размерам. а, б ПрФ 3 и LaF 3 НЧ (5 г / л) в воде. c, d ПрФ 3 и LaF 3 НЧ (5 г / л) в DMEM + FBS. е, е ПрФ 3 и LaF 3 НЧ (0,1 г / л) в DMEM + FBS. г, ч Средний гидродинамический радиус (MHR) со стандартным отклонением PrF 3 и LaF 3 НЧ в DMEM + FBS при разных уровнях концентрации
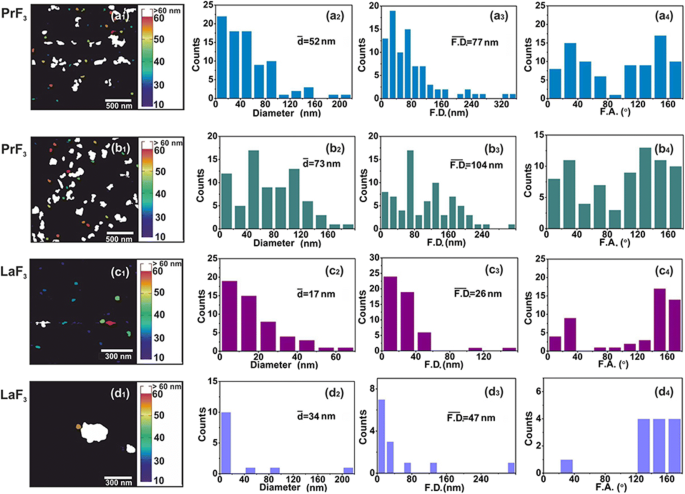
Цифровой АСМ ( x , y ) гистограммы размеров RE-NP. ( a1 – d1 ) PrF 3 (2 × 2 мкм 2 ) и LaF 3 (1 × 1 мкм 2 ) НЧ в суспензиях DMEM + FBS. ( a2 – d2 ) ( x , y ) гистограммы размеров среднего равного диаметра площади круга \ (\ left (\ overline {d} \ right) \) RE-NP. (a3 – d3) ( x , y ) размерные гистограммы диаметров области Ферета \ (\ left (\ overline {\ F.D.} \ right) \) RE-NPs. ( a4 – d4 ) Гистограммы угла Ферета (F.A.) относительно x -ось. РЗЭ-НЧ были ориентированы по двум основным направлениям между ± (44–60 °)
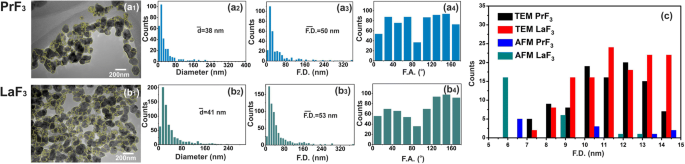
TEM digital ( x , y ) гистограммы размеров RE-NP. ( a1, b1 ) ПЭМ-изображения РЭ-НП. Желтыми линиями обозначены 2D границы между RE-NP. ( a2, b2 ) ( x , y ) гистограммы размеров среднего равного диаметра площади круга \ (\ left (\ overline {d} \ right) \) RE-NP. ( a3, b3 ) ( x , y ) гистограммы размеров диаметра области Ферета \ (\ left (\ overline {\ F.D.} \ right) \) RE-NP. ( a4, b4 ) Гистограммы угла Ферета (F.A.) относительно x - ось с преимущественными направлениями ± (44–60 °). ( c ) Размерная гистограмма диаметра Ферета малых и крошечных RE-NPs, извлеченных из изображений AFM и TEM для 4 мкм 2 области
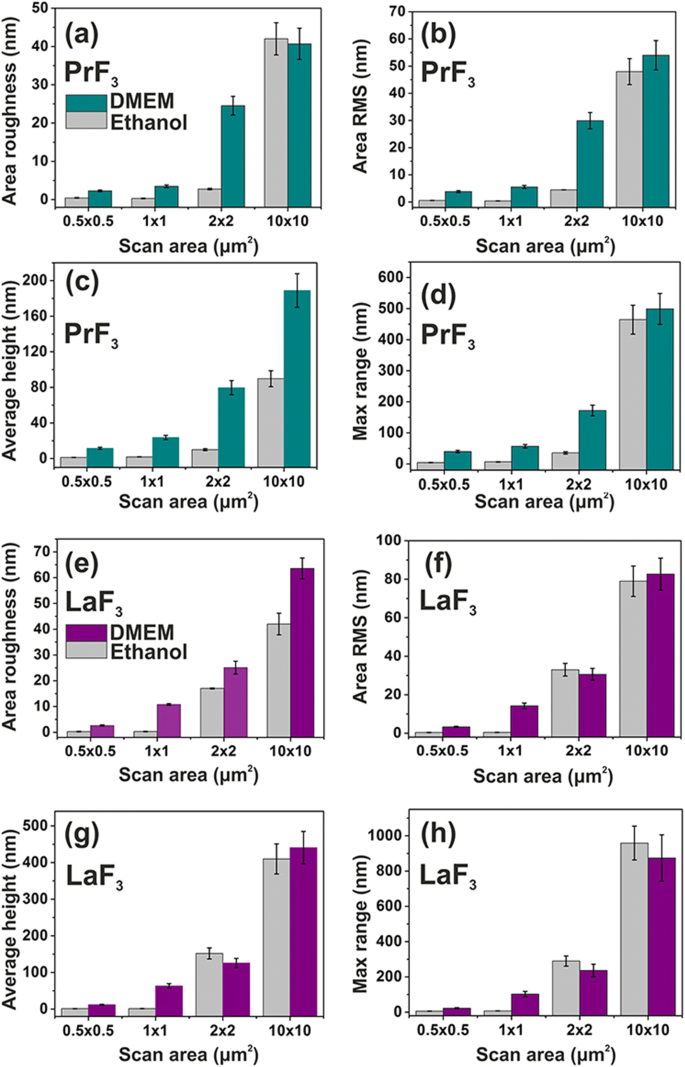
Гистограммы параметров поверхности (z-высота) PrF 3 и LaF 3 RE-NP на стеклянной подложке в DMEM + FBS и этаноле для различных областей сканирования. а , e Шероховатость поверхности. б , f Площадь RMS. c , г Средняя высота. г , ч Максимум z -диапазон

АСМ и оптические изображения высушенных RE-NP в DMEM + FBS. а - г Оптический ( a ) и изображения АСМ ( b - г ) DMEM + FBS, показывающий самоорганизующиеся структуры 5 мкм. е - ч ПЗС-матрица ( e ) и изображения АСМ ( f - ч ) ПрФ 3 НЧ в среде DMEM + FBS демонстрируют самоорганизующиеся дендритные структуры размером 500 нм. я - l ПЗС-матрица ( i ) и изображения АСМ при разном увеличении ( j - l ) из LaF 3 НЧ в DMEM + FBS демонстрируют 100 нм дендритные самоорганизующиеся структуры
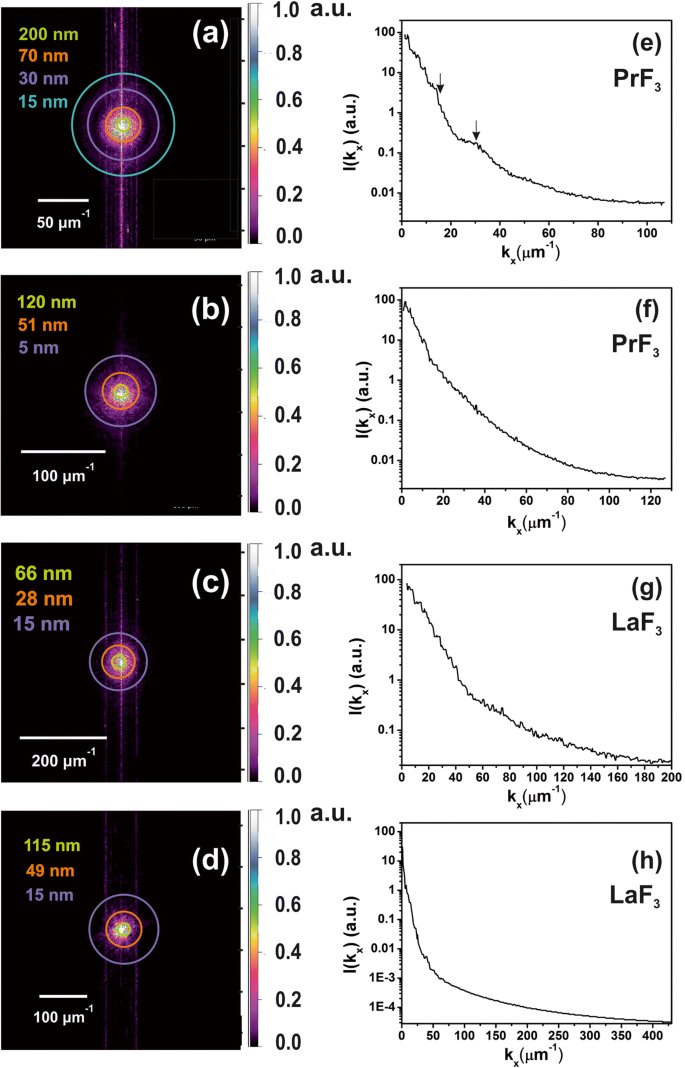
Спектры 2D-БПФ z -высокое распределение высушенных РЭ-НЧ в среде DMEM + FBS, свидетельствующее о наличии НЧ небольшого размера в жидких суспензиях. а , b z-высота распределения PrF 3 НЧ из изображений АСМ, рис. 2 (а1, б1). Маленький z -высоты (~ 5 нм) были идентифицированы в ( b ). c , d z-высота распределения LaF 3 НЧ из изображений АСМ, рис. 2 (c1, d1). е , f Спектры мощности z -высотные волновые векторы PrF 3 НП вдоль x -ось. г , ч Спектры мощности z -высотные волновые векторы LaF 3 НП вдоль x -ось
Спектры XRD a ПрФ 3 и b ЛаФ 3 НП. Средние диаметры поверхности PrF 3 и LaF 3 НЧ были ~ 23 ± 4 и ~ 15 ± 4 нм соответственно
DLS
Мутные гетерогенные смеси изначально были сформированы путем добавления RE-NP в среды для культивирования клеток, что свидетельствует о сложности взаимодействий между RE-NP в жидких суспензиях. Крошечные (<10 нм), маленькие (> 10 и <20 нм) и большие (> 20 нм) структуры RE-NP были идентифицированы как для PrF 3 и LaF 3 растворимые вещества (рис. 1a – d).
Значения среднего гидродинамического радиуса (MHR) РЗЭ-НЧ большого размера (55–83 нм для PrF 3 и 99–296 нм для LaF 3 ) следили прямо и обратно пропорционально уровню концентрации НЧ (0,1–10 кг м −3 ) в жидкой модификации Eagle’s по Дульбекко с фетальной бычьей сывороткой (DMEM + FBS), рис. 1g, h. Также MHR небольшого размера LaF 3 и PrF 3 НЧ оставались постоянными, 10,66 ± 0,74 нм и 10,64 ± 0,40 нм, соответственно, при различных уровнях концентрации RE-NP. MHR RE-NP не изменялась в течение как минимум 6 дней. После сушки суспензий РЗЭ-НЧ было невозможно повторно растворить порошок РЗЭ, поскольку крупные агломерации, стабилизированные сильными взаимодействиями, вынуждали осаждаться.
АСМ и ПЭМ-визуализация RE-NP и анализ поверхности
Для надежного распределения размеров и статистики RE-NP малых размеров в DMEM + FBS при 0,1 кг м −3 АСМ (области сканирования 1 × 1 и 2 × 2 мкм 2 ) и ПЭМ-изображение (рис. 2 (a1 – d1) и рис. 3 (a1, b1)). После переноса жидких капель РЗЭ-НЧ в DMEM + FBS на стеклянную подложку было идентифицировано относительно большое количество неагрегированных крошечных РЗЭ-НЧ [38] как по среднему размеру, так и по среднему диаметру Фере НЧ ( Рис. 2 (a2 – d2, a3 – d3) и рис. 3 (a2, b2, a3, b3)). Кроме того, гистограммы углового распределения диаметров Ферета AFM и TEM (для большего размера НЧ) показали, что оба РЗЭ-НЧ были ориентированы преимущественно в двух направлениях между ± (44–62 o ) относительно x -ось (рис. 2 (a4 – d4) и рис. 3 (a4, b4)).
Несмотря на это z -Распределение НЧ по высоте не дало никакой прямой информации об общем распределении НЧ по размеру, это удобный инструмент сравнения для первой оценки ( x , y ) по размеру, поскольку z-высота и ( x , y ) распределения остаются взаимосвязанными [38].
Средние параметры поверхности обоих PrF 3 и LaF 3 в высушенных суспензиях для различных областей сканирования АСМ также показаны на рис. 4. Маленькие z -значения высоты указывают на очень однородный z -высота распределения обоих RE-NP для малых 1 × 1 мкм 2 сканировать области. Напротив, как для RE-NP, так и для больших областей сканирования z - распределение по высоте было значительно шире. Низкий z -величины распределения по высоте на малых площадях сканирования отражают присутствие в жидких суспензиях мелких РЗЭ-НЧ. Значения параметров поверхности в среде DMEM + FBS были в среднем выше, чем в этаноле, что свидетельствует о сложном реакционном состоянии между белками и RE-NP, что согласуется с многогранной структурой на рис.5 и данными 2D-FFT (рис.6). ). В целом, LaF 3 НЧ показали интересный отклик в высушенных суспензиях и более широкие параметры шероховатости поверхности, чем PrF 3 НП.
БПФ
Цветные кольца были добавлены на выбранных радиусах в спектры 2D-FFT (рис. 6a – d). Циклы представляют различное распределение наночастиц по размеру в евклидовом пространстве, от крошечного размера, равного размеру пикселя (1,9–3,9 нм), до значительного размера ~ 2 мкм, который был верхним пределом сканирования наконечника АСМ в г -ось (рис. 6e – h). Деконволюция z -высота значения с радиусом наконечника АСМ обеспечивает фактическое разрешение в z -высота распределения ~ 5 нм. Спектры 2D-FFT продемонстрировали интенсивное распределение волновых векторов вблизи центра из-за среднего значения z -высота РЭ-НП ~ 44 нм. Шаблоны БПФ демонстрировали гало-структуру, которая постепенно размывается из-за полидисперсной широкой структуры крошечных размеров, идентифицированной в спектрах 2D-БПФ. Поскольку в спектрах появлялся только ореол без каких-либо дифракционных картин для обоих спектров 2D-FFT, регулярные самоорганизующиеся структуры отсутствовали. Характерные длины корреляции, полученные из кольцевых шаблонов 2D-БПФ PrF 3 и LaF 3 составляли ~ 51, 70 нм и 28, 49 нм, соответственно, что соответствует значениям MHR, извлеченным из спектров DLS.
XRD
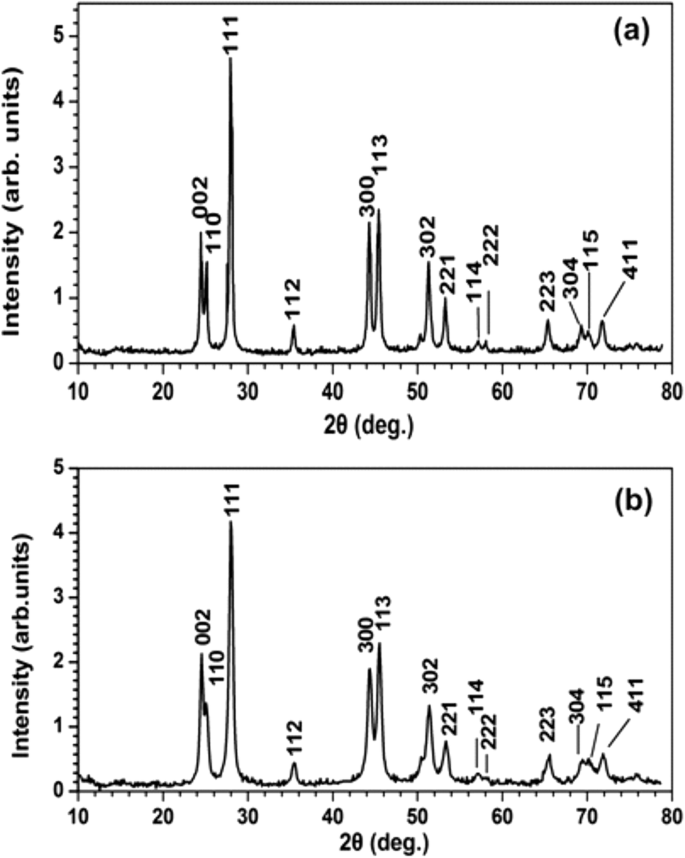
XRD-спектроскопия охарактеризовала кристаллическую структуру и предоставила дополнительную информацию о размере PrF 3 и LaF 3 НЧ (рис.7). Острые дифракционные пики, соответствующие стандартной структуре гексагональной фазы для обоих РЗЭ-НЧ, показывают высокое кристаллическое состояние агломерирующих фаз. Используя формулу Шеррера (\ (\ tau =\ frac {0.9 \ lambda} {\ beta \ cos \ left (\ theta \ right)} \ Big) \), средний средний диаметр окружности равной площади (MEAC) τ ПрФ 3 и LaF 3 НЧ составили ~ 23 ± 4 и ~ 15 ± 4 нм соответственно.
ВУФ-спектроскопия
Спектр ВУФ-пропускания гигроскопического PrF 3 Слой НЧ, нанесенный на CaF 2 субстрата от 125 нм (~ 10 эВ) до 190 нм (~ 6.5 эВ) показано на рис. 8. ВУФ-пики при 140–170 нм, ранее приписываемые переходам Pr 3+ трехвалентные ионы из земли 4f конфигурация электронного состояния для компонентов Старка 4f5d электронная конфигурация внутри YF 3 , LaF 3 , KY 3 F 10 и LiLuF 4 монокристаллической матрицы, и они перекрываются полосой поглощения воды ВУФ, показывая присутствие молекул связанной воды в PrF 3 и LaF 3 кристаллы.
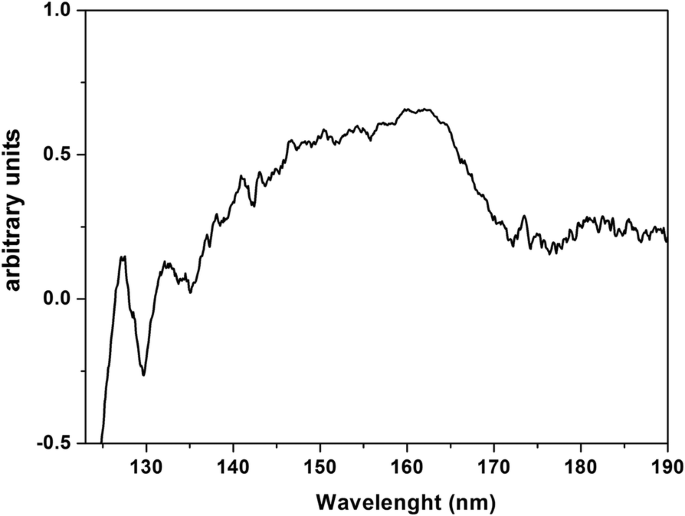
Спектр ВУФ пропускания PrF 3 НЧ в водных суспензиях, нанесенных на высушенный CaF 2 субстрат. Спектр указывает на прикрепление воды и улавливание в PrF 3 НП
Тест жизнеспособности
После анализа распределения по размерам и статистики RE-NP, тест жизнеспособности водорастворимых солей тетразолия (WST) был использован для мониторинга токсичности PrF 3 и LaF 3 NP для трех линий раковых клеток человека:A549, полученного из рака легких, SW837, полученного из рака толстой кишки, и MCF7, полученного из рака груди. Использовали три различные концентрации суспензий RE-NP (0,5, 1 и 5 мМ) в DMEM + FBS (A549, SW837) и в среде Roswell Park Memorial Institute с сывороткой плодов крупного рогатого скота (RPMI + FBS) (MCF7). Линии клеток первоначально помещали на 96-луночные планшеты и оставляли для прикрепления на ночь. Чтобы находиться в линейной области роста клеток и избежать насыщения (рис. 9а) на следующий день, свежая среда, содержащая PrF 3 и LaF 3 Были добавлены суспензии, и тесты на жизнеспособность были выполнены через 24 и 48 часов после добавления RE-NP или через 48 и 72 часа после начального момента посева клеток. Однако для трех концентраций и трех культуральных клеточных линий была обнаружена разница в избыточном росте при условии, что среда не заменялась и в культуру не добавлялись дополнительные RE-NP, что изменяет начальные условия эксперимента. Кроме того, было невозможно высеять на планшете концентрацию клеток менее ~ 5 × 10 4 . клеток на лунку, потому что слияние трех клеточных линий было слишком маленьким, чтобы гарантировать измеримый рост клеток. Оптимальная экспериментальная установка была настроена на ~ 5 × 10 4 клеток на лунку.
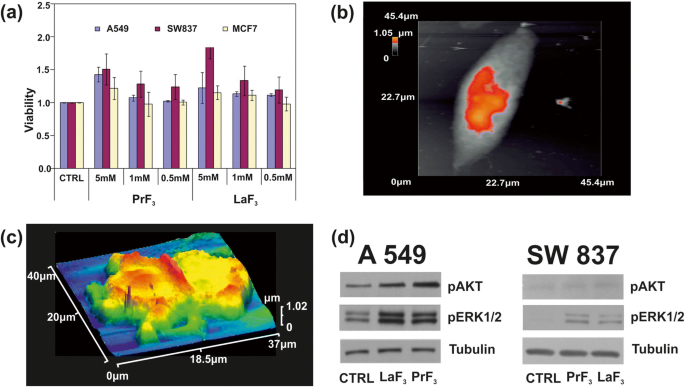
а Гистограммы анализа жизнеспособности WST трех различных линий раковых клеток (A549, SW837, MCF7), обработанных различными концентрациями PrF 3 и LaF 3 НЧ в биологических средах. б АСМ изображение одиночной раковой клетки A549. c АСМ изображение разделенной раковой клетки A549 в RE-NP в DMEM + FMS. г Анализ фосфорилирования Wb клеток A549, SW837 с путями AKT и ERK
При более высокой концентрации (5 мМ) для обеих суспензий РЭ был получен восходящий рост всех клеточных линий (рис. 9б, в). Среди них наибольший рост был у линейки SW837 (86%, LaF 3 ). Менее выраженный, но все же значимый, чрезмерный рост клеток (15%) был отмечен для линии клеток MCF7 при 5 мМ. А т тестовый статистический анализ ( p и Фишер Ф значения жизнеспособности опухолевых клеток показали, что рост опухолевых клеток был ненасыщенным через 24 часа; он следовал неизвестному физическому закону, связывающему жизнеспособность и концентрацию RE-NP (дополнительный файл 1).
Анализы фосфорилирования
Также проверяли статус фосфорилирования двух белков (фиг. 9d). С использованием специфических антител и анализов Wb в клеточных линиях A549 и SW837, выращенных в среде DMEM + FBS с 5 мМ LaF 3 и PrF 3 НЧ в течение 24 часов, была получена высокая активность фосфорилирования ERK1 / 2 и AKT в обработанных клетках по сравнению с контрольными клетками (CTRL).
Обсуждение
Относительная скорость роста раковых клеток возрастала при более высоких уровнях концентрации обоих RE-NP (фиг. 9a). Однако значения MHR RE-NP в DMEM + FBS отслеживались напрямую (PrF 3 ) и обратно пропорционально (LaF 3 ) до концентрации РЗЭ-НЧ 0,1–10 кг м −3 (Рис. 1ж, з). Следовательно, РЗЭ-НЧ со средним размером более ~ 55 нм не должны оказывать никакого влияния на рост клеток, и только НЧ крошечного размера могут принимать реальное участие в росте опухоли.
Размер и структура RE-NP
Идентификация крошечных RE-NP
Из экспериментальных данных были извлечены средний размер, распределение и статистические параметры RE-NP. Применяя t статистика теста для «нулевой гипотезы» о «среднем круге равной площади» НЧ в высушенном PrF 3 и суспензии DMEM + FBS, p значение диаметра НЧ между двумя случайно выбранными АСМ-изображениями составило ~ 0.001 (Дополнительный файл 2). Значение диаметра MEAC (63 нм) было достоверно извлечено из данных АСМ, и оно было сопоставимо со значением MHR из данных DLS (рис. 1g).
Напротив, значение диаметра MEAC из случайно выбранного LaF 3 образцы показали средний диаметр MEAC 26 нм с более высоким значением вероятности отклонения ( p =0,07), указывая на расходящееся поведение LaF 3 в жидких суспензиях. Несоответствие между диаметром MEAC и MHR из DLS (296 нм) (рис. 1) связано со сложностью взаимодействий в LaF 3 снова подвески. Действительно, для 2 × 2 мкм 2 Область сканирования наконечника АСМ, средняя z -высота была ~ 140 нм, что свидетельствует о наличии большого размера LaF 3 НЧ, перенесенные из жидких суспензий на подложку (рис. 4). Для «нулевой гипотезы» о «равных значениях диаметра MEAC из случайно выбранных образцов ТЕМ» p значения также были небольшими ( p =0,001). Для обоих RE-NP средние значения диаметра MEAC, извлеченные из объединенных данных ПЭМ и XRD для обоих PrF 3 и LaF 3 обозначен высокий p значения, p =0,29 и 0,06, соответственно, не позволяя, таким образом, корреляции между данными ПЭМ и XRD. Только ТЕМ, АСМ (PrF 3 ) и данные DLS были достаточно надежными для извлечения значений диаметра MEAC и ядра-оболочки (дополнительный файл 2).
Кроме того, неизотропное угловое распределение диаметров Ферета показало, что оба PrF 3 и LaF 3 структуры были сильно поляризованными диэлектриками, так как анизотропное угловое распределение указывает на сильные электрические полярные взаимодействия между нанокристаллами. Разнообразное поляризованное состояние LaF 3 был ответственен за снижение относительной эффективности агломерирующего состояния в суспензиях и увеличение параметров шероховатости поверхности в высушенных образцах.
Программный анализ частиц случайных изображений АСМ для 5 мкл и концентрации 0,1 кг м −3 идентифицировали ряд ~ 22 и ~ 11 РЗЭ-НЧ размером менее 15 нм и 10 нм ( p =0,001), соответственно, и ряд ~ 60 RE-NP из изображений ПЭМ ( p =0,001) на площади ~ 4 мкм 2 , подтверждая, таким образом, присутствие RE-NP крошечных размеров в суспензиях (рис. 3 (c), дополнительный файл 2), не обнаруженных с помощью DLS.
Структура и геометрия RE-NP
Распределение РЗЭ-НЧ по размерам расходится в суспензиях этанола и DMEM + FBS (рис. 4). Разнообразие обусловлено различными молекулярными взаимодействиями между адсорбированными белками, углеводами, электролитами и поверхностью RE-NP, что приводит к образованию очень сложных органических покрытий (короны), которые модулируют специфические взаимодействия RE-NP с клетками в Среда DMEM + FBS.
Взаимодействие между молекулами воды, захваченными гигроскопичными РЗЭ-НЧ, и DMEM + FBS также было жизненно важным для формирования ядра-оболочки. Он также оказал сильное влияние на белок и конформационные изменения в промежуточных взаимодействиях во время начальной фазы подготовки. Поскольку отношение поверхности к общей массе НЧ в суспензиях достигло высоких значений, эффективная стабильность, а также физико-химические, механические и текучие свойства РЗЭ-НЧ, включая способность абсорбировать белки, сильно варьировались [39,40,41]. .
Сравнительное распределение RE-NP по размерам в жидкости (DLS) и затвердевших суспензиях с помощью AFM и TEM показало, что RE-NP были инкапсулированы внутри органических форм, образующих диэлектрические структуры ядро-оболочка, где белковая оболочка окружает RE-ядро. АСМ, полученные от затвердевших RE-NP в суспензиях DMEM + FBS, нанесенных на стеклянные подложки, также указывают на образование многогранных RE-NP и белковых комплексов короны (рис. 5). В то время как высушенная среда образовывала регулярную самоорганизующуюся структуру кристаллических структур (рис. 5a – d), высушенные суспензии RE-NPs демонстрировали аморфную слоистую структуру с несколькими черными пятнами, видимыми даже с помощью цифровой камеры AFM (рис. 5e – l). . При более высоком оптическом увеличении дискретные скопления глобулярных форм, меньшие, чем в одной среде, также были обнаружены для обоих РЗЭ-НЧ в DMEM + FBS вместе со структурами дендритного типа, обе из которых демонстрируют сложность взаимодействий, что согласуется с результаты параметров поверхности (рис. 4). Даже при самом высоком разрешении АСМ (1 × 1 мкм 2 области), последняя полоса рис.5, в высушенных структурах для обоих RE-NP не было обнаружено изолированных агрегаций РЗЭ-НЧ в пределах разрешения ~ 5 нм. Сферический мицелий черного цвета длиной 1-2 мкм, показанный на оптических изображениях, представлял собой большие агломерирующие образования RE-NPs ядро-оболочка. Сложность реакций между RE-NP и DMEM + FBS была визуализирована посредством превращения длительно самоорганизующихся удлиненных структур в чистой DMEM + FBS в дендритные структуры.
Результаты указывают на картину одиночной ядерной структуры RE-NP, инкапсулированной внутри белковой оболочки. Эти структуры невозможно было обнаружить, поскольку они были окружены органическими веществами и электролитами, которые перекрестно реагировали с РЗЭ-НЧ. ВУФ-спектр PrF 3 показывает некоторые спектральные пики между 140 и 170 нм (рис. 8). Ионные переходы перекрываются полосой поглощения воды ВУФ, расширенной от 145 до 180 нм с максимумом при 168 нм. Только спектральные сигнатуры 4f6s электронная конфигурация с максимумами при 132 и 127 нм присутствовали в спектре. However, these bands could evince the presence of water in the high hygroscopic PrF3 suspensions. Water has a rich, structured absorption band in the VUV spectral range centred at 122 nm, revealing the presence of water molecules in the core-shell NPs.
Activation of Mechanosensors
Activation of Integrins by External Forces
The activation of oncogenic pathways by RE-NPs [24], besides the 3D structural nature of TSRs, is based on some Natural Evolution principles for sustaining the viability of cells. First, upon binding a specific external ligand in a LABS, conformational changes along the entire TSR spectrum underline a series of cascading pathways, triggering tumour cell growth (Fig. 9c, d). The transmission of signals advances through the plasma membrane via various protein chains. Signal transduction was via conformational transformations of integrins responding to a high affinity external force (Fig. 10a, b).

Simplified layout of integrin activation by NPs and signal transaction pathways. а Structure and conformational geometries of integrins at a low (A), medium (B) and high-affinity strengths (C). б AKT and ERK1/2 signal transaction pathways activated by RE-NPs via external integrin stimulation
Because of “life sustainability” and “survival laws” that prevents cancer cell growth by random “noise”, it is required that the strength of the external force should be within a bounded range of values and also the external strength stimulus should apply for a long period on a large number of mechanosensors in a cancer cell. The external strength that stimulates cancer must be slightly larger than the strength of the interatomic molecular forces under normal conditions. For a thermal energy of a ligand at room temperature (kT = 0.025 eV, T = 298 K), and for a regular thermal stress of molecular bonds of ~ 0.05 nm, the mean thermal force acting on the LABS stays for 1.2 × 10 −12 N. In principle, a force above ~ 10 × 10 −12 N acting coherently on the whole set of mechanosensors on a cell should activate signal transduction in tumour cells. Consequently, ignoring any thermal and mechanical stressing in the ECM normal conditions, integrin activation via electrical polar interactions between LABS and NPs has the potency to start signal transduction in cancer cells and to initiate tumorigenesis.
Integrin Structure and Geometry
An integrin receptor in the upright conformation state extends ∼ 20 nm upwards from the cell membrane [42] (Fig. 10a). For no contacts between the two α- and β-subunits, other than those in the headpiece near the ligand-binding pocket, the α- and β-subunits are well separated with their cytoplasmic tails extended out up to ∼ 8 nm [42]. A conic projection geometry (20 nm slant height, 5–10 nm diameter of its circular base), bounded by the α- and β-subunits, defines a projected area on the surface of cell’s membrane between ~ 19 and ~ 80 nm 2 , for a typical mean radius of a tumour cell R c ≈ 5 μm (equivalent surface area of a spherical cell \( {S}_c=4\pi {R}_c^2=3.14\ \mathrm{x}\ {10}^8\ {\mathrm{nm}}^2 \)). By dividing the area S c of a spherical tumour cell surface with the projected area of an integrin on a cell surface, an upper limit of the number of integrin receptors for these projected areas was n int = 1.6 x 10 7 and 3.9 × 10 6 соответственно. These numbers are compared with the mean number of integrins on a cell \( {\overline{N}}_{int}\approx 2\ \mathrm{x}\ {10}^5 \) and for an average interspacing of 45 nm between adjacent integrin receptors [43]. Nevertheless, \( {\overline{N}}_{int} \) might be larger because of an uneven surface structure, different separating distances between integrins and variable size of tumour cells (Fig. 9c), but the number of integrins on a cell membrane stand between n int and \( {\overline{N}}_{int} \).
Interaction of Mechanosensors with RE-NPs
ERK ½ and AKT Activation
The TEM images and the elemental mapping of F, La and Pr showed that RE-NPs were unable to penetrate inside the cell. They gathered around the A549 cell membrane (Fig. 11), confirming that an external force can stimulate cell growth because of TSRs activation [44]. The Pr atoms were distributed around the boundaries of the cell’s membrane. The small numbers of F, La and Pr identifications inside the cell were not associated with endocytosis of RE-NPs, but they were images of RE-NPs from the projections of the two cells hemispheres on cell’s equatorial cycle.

TEM images and elemental analysis of RE-NPs at the surface boundaries of A549 cells. а TEM image of small size LaF3 NPs surrounding the A549 cells. б Elemental analysis of F atoms in RE-NPs distributed around the cell. c Elemental analysis of La atoms. The low concentration of La atoms was associated with a rather small scattering efficiency of the X-rays. г - е The same as for (a - c ) for PrF3 RE-NPs
It was also evident that both RE-NPs were able to enhance AKT phosphorylation, especially in A549 cells (Fig. 9d), where the steady-state level of AKT pathway activity was higher for the SW837 cell line. The phosphorylation level for the MCF7 cell line was below the detection limit, in agreement with the relatively low levels of growth. High phosphorylation levels of ERK1/2 [36] and AKT were detected in A549 and SW837 cell lines. Cell growth was started once NPs with a proper size interact with the mechanosensors of the cells to provide the correct force for initiating cell growth [45, 46]. ERK and AKT pathways were frequently active in several cancer cell types via extracellular springing, as they were stimulated by the TSRs, upon a selective binding with various mitogenic ligands, or via the activation of the mechanosensory group. The interaction was responsible for a continuous intracellular stimulation that, according to the cell’s phenotype, driven the cancer cells to uncontrolled and endless growth. Viability tests were also run for 48 and 72 h, but the growth of all cell lines was saturated at 48 and 72 h after the initial moment of Cell plating.
Interaction of Cells with Ions
Likewise, as fluoride anions are the most reactive electronegative elements and, the mean radii extension of the unscreened 4f electronic configuration of La and Pr trivalent ions are relatively large, high electric surface charges could be developed via electric dipole interactions [47].
One crucial question stands whether a single ion binding on a specific site can activate tumour cell growth. Because the projected area of the 4f electronic configuration of a single RE ion is S 4f = 0.040 and 0.043 nm 2 (for an approximated spherical geometry of the 4f electronic configuration and a 4f mean orbital radii ~ r 4f =0.113 and 0.117 nm for Pr and La ions, respectively), a typical upper limit number of single RE ions, or other equivalent size ions, over the whole area of the cell membrane was ~ Sc / S 4f = N 4f ~7.9 × 10 9 RE ions; a number which is at least two orders of magnitude above the upper limit of the mean number of integrins on a tumour cell. As the relative overgrowth of cells was ascending with rising concentration (Fig. 9a), it is unlikely that tumour cell growth is triggered by a specific binding of single trivalent RE ions [48] on the ligand sites [49,50,51]. Indeed, the large number of RE ions should have saturated the cell’s growth and thus the viability of cells should have remain independent from the concentration of the RE ions.
Interaction of Integrins with RE-NPs
Within the requisite force range of few pN, and for efficient activation of integrins from NPs, the interaction between NPs and LABS should activate a large fraction of integrins of the cell for a long time. In the most extreme favoured case for cell growth, the number of NPs had to remain equal with the number of integrins on the cell’s surface, and the interactive force between LABS and NPs has to be attractive for obtaining a constant (long-term) action. A thin spherical shell of spherical NPs surrounding a tumour cell occupied a volume\( {V}_{sc}\approx 4\uppi {R}_c^2x \), where R c = 5 μm is the cell radius and x ≈ 20 nm is half the separating distance between adjacent integrin receptors and V sc ≈ 6.3 x 10 9 nm 3 . For justifying the requirement that each integrin receptor interacts only with one NP, a first estimation of the size of NPs to meet the above requirements for the whole set of integrins on a cell is obtained by dividing the volume of the spherical shell V sc with the number of integrins. A simple calculation for a cell radius 5 μm shows that the limits of radii of NPs activating the whole set of integrins within the spherical shell volume V sc ≈ 6.3 x 10 9 nm 3 covering the cell is obtained by divided the volume V sc with the number of integrins \( {\overline{N}}_{int}\approx 2\ \mathrm{x}\ {10}^5 \) and n int ≈ 1.6 x 10 7 . The volume of the spherical NPs stands for 3.15 × 10 4 and 3.93 × 10 2 nm 3 соответственно. Therefore, the radii of the NPs interacting with an integrin lay between ~ 20 and 5 nm. Allowing for one order of magnitude variations in the number of integrins \( {\overline{N}}_{int} \), the radii of the NPs interacting with integrins is between ~ 27 and ~ 3 nm respectively.
By also applying similar simple calculations and within the experimental limits of concentration levels of RE-NPs (0.1–10 kg m −3 ), the maximum numbers of PrF3 with MHR 55–83 nm and LaF3 with MHR 296–100 nm NPs (Fig. 1g, h) covering the surface of a tumour cell V sc stood for 4.1 × 10 4 –2.1 × 10 4 and 17.1 × 10 2 –1.5 × 10 4 НП. These values are placed well below the number of integrins on the cell surface. For rising concentrations of PrF3 and LaF3 from 0.1 and 10 kg m −3 , the number of PrF3 and LaF3 NPs in the suspensions must go up for either descending or ascending size of NPs. As viabilities of cancer cells are raised at higher concentration levels, it is unlikely that 55–296 nm sized RE-NPs are responsible for cancer cell mitosis under the current experimental configuration.
Also, from the DLS data, the size of both RE-NPs between 10 and 20 nm remained constant (10.6 nm) at different RE concentrations. The number of RE-NPs with this size covering the cell surface is between 3.7 × 10 5 and 1.5 × 10 6 . This number is comparable with the mean number of integrins \( {\overline{N}}_{int}\approx 2\ \mathrm{x}\ {10}^5 \) on a cell surface. Therefore, only small size RE-NPs have the potency to stimulate cancer cell growth by stimulating all the integrins on a cell surface, in agreement with the experimental observations (Figs. 1g, h and 9a).
The number of tiny sizes RE-NPs with MEAC diameter (TEM) from 2 to 10 and 10 to 15 nm on the cell surface (S c = 314 μm 2 ) stands for 1.3 × 10 4 and 1.8 × 10 4 RE-NPs, respectively. Those values stayed one order of magnitude below \( {\overline{N}}_{int}\approx 2\ \mathrm{x}\ {10}^5 \) and therefore tiny size RE-NPs had also the potency to justify the experimental results of rising viability values with concentration (Fig. 9a). Also, the rough surface of tumour cell (Fig. 9c) is able to form cavities, where small size RE-NPs are trapped, triggering thus cell’s mechanosensors. Most important, only tiny size RE-NPs have the potency to activate integrin receptors via electrical dipole interactions (vide infra).
Interaction of EGFR with RE-NPs
An upper limit of small size NPs capable of stimulating cell’s overgrowth via the EGFR was set previously to 14 nm [52], but a realistic size of NPs stimulating the EGFR should be < 5 nm [53] (Fig. 12). The area number density of EGFR on the surface of tumour cells stands for ~ 1.4 × 10 −4 nm −2 and the total number of EGFR on the surface S c of cells remains between ~ 4.2 x 10 4 and 10 5 [54,55,56]. RE-NPs with 5–10 nm size stayed for a number of 34 NPs (Fig. 3). Extrapolating this number to the surface of a cell S c , the total number of RE-NPs remained at ~ 10 4 NPs, a number which matches the number of EGFR receptors on a A549 cell. Therefore, the EGFR have the potency to be activated synergistically also by a number of tiny size RE-NPs.

AKT and ERK1/2 signal transduction pathways activated by RE-NPs via EGFR stimulation. EGFR is activated only by tiny size ~ 5 nm NPs
Electric Dipole Interaction Between RE-NPs and LABS
The above experimental results are supported by the hypothesis of cancer cell growth from LABS stimulation by tiny size core-shell RE-NPs via electrical dipole interactions, Appendix.
Indeed, the mean electrical dipole force \( \left\langle {\overrightarrow{F}}_{V_2}\right\rangle \)acting on LABS from a core-shell RE-NP includes two terms (Fig. 13d and Appendix, Eq. A22). The first radial term is inversely proportional to the forth power of separating distance r 1 between the RE-NPs and LABS and is also proportional to the size of NP. The second polar term is inversely proportional to both the separating distance r 1 and the square power of the size of NP,
$$ \left\langle {\overrightarrow{F}}_{V_2}\right\rangle =-\frac{G_1{N}_2{N}_1\ d{e}^2}{4{\varepsilon}_0\ {r}_1\ }\theta \left(\ 3G\frac{b}{r_1^3}{\widehat{r}}_1+\frac{\theta }{2{b}^2}{\widehat{\theta}}_1\right)\kern0.75em (1) $$
а Electrical dipole interaction between one core-shell RE-NP and one LABS. б , c RE core-shell NP near a MIDAS (b ) and ADMIDAS (c ) adhesion sites. г Locus area (green) of the size of RE-NPs and separating distance between a LABS and a core-shell RE-NP for two electrical charging states
В формуле. 1, G и G 1 are the geometrical factors of NPs, describing either core-shell or core spherical structures, Appendix, Eqs. A6 and A14; N 1 , N 2 are the numbers of surface electrons on the a RE-NPs and LABS surfaces; г и b are the effective characteristic spatial extension of atomic orbitals of LABS, ~ 0.1 nm, and the radius of RE-NP; е и ε 0 are the electron charge and the vacuum permittivity and \( \theta =\frac{d}{r_1}<0.01\ rad \). Because the core of the RE-NPs is a crystalline semiconductive material, an inherent large number of surface and volume defective sites were accountable for a high density of pseudo-electron energy levels that allowed the electrons to move freely within the core volume [46]. Consequently, a core-shell structure had the potency to be highly polarised. Therefore, LABS can be activated efficiently by core-shell RE-NPs via electrical dipole interactions at close separating distances. The high polarised efficiency of the core nucleus was confirmed experimentally via the selective orientation of NPs along two distinct directions (Fig. 2(a4–d4) and Fig. 3(a4, b4)).
The polar interaction force is also proportional to the geometrical factor G 1 , Appendix, Eq. A14. Typical values of dielectric constants of the culture media, shell configuration and RE core components stand for ε 1 = 78, ε 2 = 10 and ε 3 = 15. When the ratio of core-shell to core radii b/a sets within 1 and 50, the geometrical factors G , G 1 retain almost constant values (G = 0.2, G 1 = 0.01) and they are self-same for both a spherical core (b/a = 1) and a spherical core-shell. Any permanent or induced polarisation of an open or closed a-I-MIDAS domain forming the LABS domain has its origin on six coordinated water oxygen atomic orbitals with Mn 2+ or Mg 2+ ions, arranged in a spherical geometric configuration [7] (Fig. 12a–c).
As the electrical dipole force in Eq. 1 stands for the vector sum of a radial (first term) and a polar component (second term), the last term prevails over the first one provided that
$$ {r}_1>\sqrt[3]{6G}b\sim b\kern0.75em (2) $$In this case, a LABS is activated from the polar force component for all (b/a) ratios and, most important this term is inversely proportional to the second power of the size of NPs, in agreement with the experimental results that only tiny or small size LaF3 NPs activated cancer cell proliferation.
The prevailed polar force term for different r 1 и b values and for different Ν 1 , Ν 2 charging states activating the LABS/MIDAS stay within the limits [57,58,59,60].
$$ {10}^{-12}N<\frac{G_1{N}_2{N}_1\ d{e}^2}{8{\varepsilon}_0\ {r}_1{b}^2\ }{\theta}^2<{10}^{-9}N\kern1em (3) $$Inequality 3 relates the size b of the RE core-shell NPs, the separating distance r 1 and the number of the bound or free electrons Ν 2 , Ν 1 on the surface of the two dipoles. The locus of points (r 1 , b ) satisfying the inequality 3 for different surface charge states Ν 1 , Ν 2 is bounded by the black, red and blue lines (Fig. 13c). As there was no specific assumptions for the type of RE-NP, results can be equally applied for any type of polarised NPs.
When the algebraic product of the number of the surface electrons N 1 и N 2 (bound or free) on the LABS and the RE-NP, respectively, was N 1 N 2 = 2, the locus of RE-NPs size and separating distance for integrin activation was < 1 nm. At higher charging states, N 1 N 2 =10 4 , the locus area spans a wider RE-NPs size and separating distance area set of values, from 0.5 nm–19 nm to 2.5–15 nm, respectively.
From the above analysis, it is found that only tiny or small size NPs can activate LABS at a certain separating distance r 1 and the electrical dipole interaction strength decays inversely proportional to the second power of the size of NPs. From Fig. 13c and for a charging state with N 1 N 2 = 5 x 10 4 , the size of NPs capable to activate LABS is bounded by the limits
$$ 2.5\ \mathrm{nm}Выводы
Cancer is a complex disease. Tumours are highly heterogeneous, and cell growth, among other factors, depends on dynamical interactions between cells and the continually changing extracellular matrix. Besides random genomic mutations, signal transductions in cells, activating cell growth can be triggered by mechanical, thermodynamic and electrical polar interactions between the microenvironment of the extracellular cell matrix and the membrane’s mechanosensors. Here, we demonstrated that tumour cell proliferation in three different human cancer cell lines (A549, SW837, MCF7) had the potency to be activated by a synchronised and synergetic activation of EGFR or via electrical dipole interactions between tiny size RE-NPs and the LABS of integrins on a cell.
Because the prerequisite force for integrin activation should stand between 10 −12 and 10 −9 N, the size of the active RE-NPs causing cell growth should be within certain limits. Cancer activation is specified by both the electrical surface charges on the LABS and the NPs and by their separating distance. This electric dipole activating force follows an inversely proportional square power law of the radius of NPs, evidencing that only tiny or small size RE-NPs have the potency to stimulate cancer cell growth via electrical dipole interactions, in agreement with the experimental results.
Methods
Synthesis of RE-NPs
PrF3 NPs were synthesised via co-precipitation. Briefly, 4 g of Pr2 О 3 were added to 110 mL of 10% nitric acid in a polypropylene glass beaker together with 3 g of NaF under stirring. The mixture was heated to 50 °С and stirred for 45 min until a clear light-green solution appeared. Then it was filtered. The pH of the mixture adjusted to 4 by adding 25% of ammonium hydrate. Next, the mixture was stirred again for 20 min. Finally, the precipitated NPs washed with distilled water by centrifugation.
LaF3 NPs were also synthesised by applying the same protocol in a mixture of La2 О 3 (4 g) and NaF (3 g). From both preparations, an aliquot of the suspensions containing NPs was air-dried for structural analysis and the remaining part kept as water suspension for the biological studies.
The suspensions of NPs were prepared in complete DMEM+FBS cell culture medium by adding water suspended NPs directly to the medium to a final concentration of 5 mM. Then, starting from the 5 mM stock solution, some subsequent dilutions using DMEM as a solvent were prepared to a final NPs concentration of 1 mM and 0.5 mM, respectively.
Size Distribution of RE-NPs
XRD
The crystal structure and the size of PrF3 and LaF3 NPs were characterised by XRD spectroscopy, with an X-ray diffractometer (Shimadzu XRD-7000S) in the 2θ range from 10° to 80° using the graphite monochromatised Cu-Ka radiation (1.5406 Å). The weighted average of τ for all peaks was used in the statistics. Weighting, besides β, took into account the relative intensity of every peak of the XRD spectra. The corresponding errors incorporate the reading error (0.3 mrad) and the standard error of the mean (se = σ / √ Ν ).
DLS
The size distribution and the MHR of RE-NPs in water and DMEM+FBS suspension were determined for comparison by DLS at 632.8 nm and right angles at 37 °C with a multi-angle dynamic and static light scattering instrument (PHOTOCOR-FC). The values of the MHR (Stokes radius) and the size distribution of NPs were calculated from the autocorrelation spectra and the Stokes-Einstein relation with the DynaLS software. Because the intensity of scattered light in pure DMEM+FBS was 20 times lower than with RE-NPs additives, the level of aggregating proteins in pure DMEM+FBS was negligible compared with mixed suspensions of RE-NPs in DMEM+FBS medium. MHR and RE-NPs size distribution and size errors were obtained by fitting and processing the data from the DLS instrument with the DynaLS software that allows the MHR to be calculated in different spectral domains of the main size distributions, from 10–10 2 to 10 2 –10 3 nm, Additional file 2.
AFM
Because size distribution below 15 nm was close to the low limit range of DLS, AFM was also applied to evaluate small size distribution. At low concentration of RE-NPs in liquid suspensions and slow drying rates of droplets on glass substrates, the deposits reflected the size distribution in the liquid suspensions [37]. Following the dispersion of RE-NPs in ethanol or DMEM+FBS, a drop of suspension was placed on a clean glass substrate using a micropipette, and then it was dried in air at room temperature for AFM imaging and analysis (diInnova, Bruker). AFM was performed in the tapping mode, in ambient conditions with a phosphorus-(n)-doped silicon cantilever (Bruker, RTESPA-CP), having a nominal spring constant of 40 nN/nm and operating at a resonance frequency of 300 kHz. Surface areas of various sizes (0.5 × 0.5–50 × 50 μm 2 ) were imaged with high spatial resolution (512 px × 512 px) at a scanning rate of 0.2 Hz to identify domains with different size distributions via “scan area filtering” [37]. From the morphological analysis by the SPM LabAnalysis V7 software, the particle’s size distribution, shape and aggregation stage were determined.
The size of NPs for different scanning areas was also noticeable by the particle analysis chromatic bar (Fiji integrated ROI colour coder based on MEAC diameter) (Fig. 2(a1–d1)). The AFM image was transformed into a binary image using an appropriate z -height threshold. Every pixel of the processed image contained information not only for the z -height in the pixel area but also for the presence of particles in the pixel area. x -histograms of MEAC and Ferret diameter (Fig. 2(a2–d2, a3–d3)) were extracted by using the “Image J 1.51n Fiji distribution software”, with the correct z -height threshold values. The size resolution per pixel was 3.9 and 1.9 nm for PrF3 and LaF3 соответственно.
The particle identification, the noise extraction and the particle area data were processed by the “Particle Analyser function” of Fiji software (Fig. 2(a1–d1)). The particle diameter histograms were also analysed. Both the equal area circle diameter (Fig. 2(a2–d2)) and Feret diameter or “calliper diameter” (maximum diameter of a particle among all directions) (Fig. 2(a3–d3)), whose direction was the Feret angle (Fig. 2(a4–d4)), were analysed. The mean equal area circle diameter and the mean Feret diameter were calculated taking into account all particles identified. The associated errors incorporated the actual pixel size in every AFM image and the standard error of the mean (se = σ / √ Ν ).
А т test was performed for every set of AFM images based in the “null hypothesis” that the mean particle diameter was the same for all the AFM images between randomly selected figures (Fig. 2(a1–b1, c1, d1)). p value (probability that the null hypothesis based on t distribution is not valid) is shown in Additional file 2.
TEM
The same technique was followed for calculating the above parameters in TEM imaging (Fig. 3(a1–b4)). Atomic resolution TEM (Hitachi HT7700 Exalens) imaged either extracellular or intracellular RE-NPs attachment on the A549 cells fixed in glutaraldehyde. Elemental analysis of F, La and Pr were also carried out (Oxford Instruments X-Max 80T).
2D-FFT
Additional information on the NPs size distribution in the (x , y ) plane was also extracted from the 2-D Fourier transform of AFM images of NPs using the relation
$$ I\left({k}_x,{k}_y\ \right)=\iint f\left(x,y\right)\exp \left(i{k}_xx\right)\exp \left(i{k}_yy\right) dxdy $$где f ( x , y ) is a size function at a point (x, y ), k x , k y are the associated wavevectors in the inverse Eukledian space at the same point and I ( к x , k y ) is the “spectral density” of the function f ( x , y ) at the point k x , k y . For most applications, f ( x , y ) is the z -height of the NPs at the point (x, y ) and z = f ( x , y ).
For a set of discrete data, such as the digitised AFM images, the 2D-FFT was used instead of 2D Fourier transform in the continuous space. For a m × n X-matrix (pixels of an AFM image), the 2D-FFT transform takes the form
$$ \kern1em {Y}_{p+1,q+1}=\sum \limits_{j=0}^{m-1}\sum \limits_{k=0}^{n-1}{\omega}_m^{jp}{\omega}_n^{kq}{X}_{j+1,k+1\kern1.25em } $$where \( {\omega}_m^{jp}={e}^{2 pi/m},{\omega}_n^{kq}={e}^{2 pi/n} \) are the associated frequencies. Then, an appropriate shift along the y -axis was performed and the integers m, n, p, q, k were translated into lengths and inverse lengths respectively by a multiplication with the pixel’s size of the image.
Water Trapping in RE-NPs
VUV Spectroscopy
To appraise the state of water in RE-NP’s complexes during the initial stage of suspension preparation, the adsorption of water molecules on the surface of the hygroscopic PrF3 NPs was identified with a laboratory-made VUV (110–180 nm) absorption spectrometer. It consists of a hydrogen lamp operating in a longitudinal stabilised discharge mode at 10 kV, a stainless steel vacuum chamber and a VUV monochromator (Acton VM502), equipped with a solar blind photomultiplier (Thorn EMI 9412 CsTe) and a laboratory-made data collection system. Thin layers of PrF3 NPs suspensions in water were prepared and dried on 1-mm-thick VUV-grade CaF2 substrates by applying the “drop-casting method”. Then, the CaF2 substrates were placed in the optical path between the hydrogen lamp and the VUV monochromator in a vacuum. The stainless steel 316 vacuum chamber was evacuated initially to 10 − 7 mbar using two turbomolecular pumps at a differential pumping configuration (Edwards EXT 100/200, pumping speed 150 ls −1 ). However, a high outgassing rate of PrF3 sets an upper limit to the background pressure in the vacuum chamber ~ 8.5 × 10 −5 мбар. The relatively low background pressure of both compounds irreversibly damages the VUV optics and the turbomolecular pump after few hours of operation and therefore it sets certain experimental constraints, preventing an equivalent registration of LaF3 spectrum because of high outgassing rates and a low background operating pressure (< 10 −4 mbar). The experimental data (light transmitted through the sample film on CaF2 window) were fitted to a logarithmic response for calculating the transmittance.
Cell Culture and Growth Assay
Cell Growth
The A549 and SW837 cell lines were maintained in DMEM+FBS, whereas the MCF7 lines were in RPMI+FBS. Both media supplemented with 10% fetal bovine serum (FBS), 1 × penicillin, 1 × streptomycin and 2 mM l-glutamine. Cells were incubated at 37 °C, 5% CO2 in a humidified atmosphere.
The WST viability test was used to monitor the intrinsic toxicity of PrF3 and LaF3 NPs for three human cancer cell lines, A549, SW837 and MCF7. For the viability assay, three different concentrations of RE solubles (0.5, 1 and 5 mM) in DMEM+FBS (A549, SW837) and RPMI+FBS (MCF7) were used. The initial number of cells seeded in the 96-well plates was ~ 5 × 10 4 ячеек / лунка. This amount of cells was plated 24 h prior to the RE-NPs treatment of cells in order to allow enough time for the cells to attach properly to the plate (wells) and to attain the optimum growing conditions. Subsequently, the viability test was performed 24 h after RE-NPs addition, or 48 h after the initial cell cultures were placed in the wells. As we did not observe any cell reduction, but on the contrary cell-overgrowth, especially with the SW620 cell line at 5 mM, the cell confluence quickly reached 80–90% of its initial value after 24 h of the addition of RE-NPs or 48 h from the initial plating.
Five microliters of WST solution was added to each well and the plate was incubated for 1 h during the growth state. The absorbance at 450 nm of each well was measured using a microplate reader (Biorad, x Mark). Each experimental point for each cell line and each RE suspension was extracted from two samples and triplicated every 2 days (total of 108 samples).
F test was used for every set of cell viability measurements. Here, the “null hypothesis” was that the relative to the CTRL “mean viability value was the same at different concentrations within the same cell line”. With this null hypothesis, an unknown law connecting tumour cell viability and RE-NPs concentration was identified. p value (probability the null hypothesis to be rejected) was also tested from the F distribution Additional file 1.
Western Blotting and Antibodies
Total proteins were extracted with 60 μL of radioimmunoprecipitation assay (RIPA) lysis buffer (20 mM Tris-HCl (pH 7.5); 150 mM NaCl, 1 mM Na2 EDTA; 1 mM EGTA; 1% NP-40; 1% sodium deoxycholate; 2.5 mM sodium pyrophosphate; supplemented with proteases inhibitors 1 mM β-glycerophosphate; 1 mM Na3 VO4 1 μg/ml; leupeptin) and the Wb assay was performed according to standard protocols (Fig. 9b). Briefly, total proteins (50 μg) were separated by SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membrane. Blots were incubated overnight at 4 °C with appropriate primary antibodies. The antibodies used were tubulin code sc-8035, from Santa Cruz (final concentration 1:1000 in blocking buffer); p-ERK (E-4) code sc-7383, from Santa Cruz (final concentration 1:500 in blocking buffer); and p-AKT (Thr308) code 9275S, from Cell Signaling (final concentration 1:1000 in blocking buffer).
Wb bands are collected from different blots showing quality control of antibodies specificity. Numbers at the top of the phosphorylation images show grey scale levels from 0 (black) to 168 (grey) (maximum value ), indicating activation at a non-saturated mode.
Сокращения
- 2D-FFT:
-
Two-dimensional fast Fourier transform
- ADMIDAS:
-
Adjacent MIDAS
- AFM:
-
Атомно-силовая микроскопия
- AKT:
-
Protein kinase B
- CTRL:
-
Control cells
- DLS:
-
Динамическое рассеяние света
- DMEM:
-
Модифицированный носитель Орла, созданный Дульбекко
- ECM:
-
Cell-extracellular matrix
- EGFR:
-
Epidermal growth factor receptors
- ERK:
-
Extracellular signal-regulated kinase
- F.A.:
-
Feret angle
- F.D.:
-
Feret area diameters
- FBS:
-
Fetal bovine serum
- LABS:
-
Ligand adhesion binding site
- MEAC:
-
Mean equal area circle
- MHR:
-
Mean hydrodynamic radius
- MIDAS:
-
Metal ion-dependent adhesion sites
- NGFR:
-
Nerve growth factor receptor
- NP:
-
Наночастицы
- RE-NPs:
-
Rare-earth nanoparticles
- RIPA:
-
Radioimmunoprecipitation assay
- RMS:
-
Среднеквадратичное значение
- RPMI:
-
Roswell Park Memorial Institute medium
- SDS-PAGE:
-
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis
- SyMBS:
-
Synergistic metal ion binding sites
- ТЕМ:
-
Просвечивающая электронная микроскопия
- TSR:
-
Transmembrane signal receptors
- VEGFR:
-
Vascular endothelial growth factor
- VUV:
-
Vacuum ultraviolet
- Wb:
-
Western blot assays
- WST:
-
Water-soluble tetrazolium salts
- XRD:
-
Рентгеновская дифракция
Наноматериалы
- О полупроводниковых наночастицах
- Биосовместимые наночастицы FePO4:доставка лекарств, стабилизация РНК и функциональная активность
- Изучение in vitro влияния наночастиц Au на клеточные линии HT29 и SPEV
- Модифицированный гиперразветвленный полиглицерин как диспергатор для контроля размера и стабилизации нано…
- Стимуляция роста клеток SH-SY5Y наночастицами золота, модифицированными 6-меркаптопурином и проникающим в нейро…
- Наночастицы маггемита действуют как нанозимы, улучшая рост и устойчивость к абиотическому стрессу у Brassica napus
- Излучающие в ближнем инфракрасном диапазоне Cr3 + / Eu3 + со-легированные галлогерманатом цинка стойкие люминесц…
- Нанокомпозиты на основе оксида графена, украшенные наночастицами серебра в качестве антибактериального аге…
- Золотые наночастицы круглой формы:влияние размера и концентрации частиц на рост корней Arabidopsis thaliana
- Синтез биметаллических вольфрам-медных наночастиц in situ с помощью реактивной радиочастотной (RF) тепловой плаз…