


Влияние инженерных наночастиц на высвобождение экзополимерных веществ из морского фитопланктона
Аннотация
Разработанные наночастицы (ENP), продукты современных нанотехнологий, могут потенциально воздействовать на морскую среду, создавая серьезные угрозы для морских экосистем. Однако клеточные ответы морского фитопланктона на ENP все еще не установлены. Здесь мы исследуем четыре разных вида диатомовых водорослей ( Odontella mobiliensis , Skeletonema grethae , Phaeodactylum tricornutum , Thalassiosira pseudonana ) и одна зеленая водоросль ( Dunaliella tertiolecta ) для высвобождения их внеклеточных полимерных веществ (EPS) при модельных обработках ENP:диоксид титана 25 нм (TiO 2 ), Диоксид кремния 10–20 нм (SiO 2 ) и диоксида церия 15–30 нм (CeO 2 ). Мы нашли SiO 2 ENP могут значительно стимулировать высвобождение EPS из этих водорослей (200–800%), в то время как TiO 2 Воздействие ENP вызывало наименьшее высвобождение. Кроме того, увеличение внутриклеточного Ca 2+ концентрация может быть вызвана ENPs, предполагая, что процесс высвобождения EPS опосредуется через Ca 2+ сигнальные пути. При лучшем понимании клеточного механизма, опосредованного ЭПС-индуцированным высвобождением ЭПС, можно разработать потенциальные превентивные меры и меры безопасности для смягчения негативного воздействия на морскую экосистему.
Фон
Технические наночастицы (ENP), размер которых варьируется от 1 до 100 нм (по крайней мере, в одном измерении), используются в производстве многочисленных потребительских товаров, включая чернила и краски для принтеров, моющие средства, бактерициды, покрытия, косметику, солнцезащитные лосьоны, шины, компьютерное строительство и доставка лекарств. Учитывая многообещающее применение ЕПС, финансирование Национальной инициативы в области нанотехнологий (NNI) только в США в 2017 году приблизилось к 1,4 млрд долларов США [1,2,3]. Установление фундаментальных знаний в наномасштабе было основным направлением исследовательского сообщества в области нанотехнологий на первом этапе. По состоянию на 2009 год эти новые знания лежали в основе примерно четверти триллиона долларов на мировом рынке, из которых около 91 миллиарда долларов приходилось на американские продукты, включающие наноразмерные компоненты [4]. С быстрым развитием нанотехнологий неизбежно, что ЕПС в конечном итоге найдут свой путь к водным системам. Основная проблема с ENP с точки зрения их потенциальной токсичности (например, потенциал для образования активных форм кислорода, ROS) в окружающей среде связана с их большой и уникальной поверхностной реактивностью. Однако фактическое воздействие на морскую экосистему остается в значительной степени неизвестным из-за сложных экологических и биологических факторов природных вод и разнообразия ЕПС [1, 5, 6]. Предыдущие исследования показали, что ENP могут нанести значительный ущерб морской экосистеме, основанной на водорослях [7, 8]. Было показано, что морские организмы (особенно фитопланктон) взаимодействуют с ENP, что приводит к негативным последствиям [9,10,11]. С потенциальным увеличением использования нанотехнологий в различных областях, все больше и больше ENP могут попадать в водную среду, поэтому клеточные реакции морского фитопланктона на ENP требуют дальнейшего внимания [12,13,14,15,16,17,18,19,20, 21].
Большинство морских микробов, как авто-, так и гетеротрофных, обычно способны продуцировать экзополимерные вещества (EPS), которые имеют различные функциональные роли и физические свойства в морской экосистеме, действуя как ингибиторы роста, стимуляторы роста, токсины, поглотители металлов или как субстраты для гетеротрофный цикл [22,23,24,25,26]. EPS, выделяемый фитопланктоном и бактериями в океане, представляет собой богатые полисахаридами анионные коллоидные биополимеры, которые имеют решающее значение для образования морских гелей, морского снега и биопленок, а также для удаления коллоидов и микроэлементов и обеспечения защиты от различных экологических угроз. в том числе ЕПС [7, 15, 19, 20, 25, 27]. Кроме того, секреция ЭПС считается естественной реакцией, когда фитопланктон испытывает различные стрессы [8].
Ca 2+ является обычным вторичным мессенджером, участвующим во множестве внутриклеточных сигнальных путей. Было продемонстрировано, что Ca 2+ необходим для хемотаксиса, подвижности и адгезии диатомовых водорослей Amphora coffeaeformis [28]. Повышенный уровень свободного внутриклеточного Ca 2+ уровни, как известно, приводят к активации протеинкиназы С, которая участвует во многих внутриклеточных сигнальных путях [29]. Поскольку высвобождение EPS тесно связано с подвижностью и адгезией диатомовых водорослей, было предложено, чтобы Ca 2+ -опосредованный процесс секреции контролирует высвобождение EPS из диатомовых водорослей [30] и является прямым доказательством, подтверждающим Ca 2+ передача сигналов, экзоцитоз и корреляция Ca 2+ передача сигналов с экзоцитозом описана в нашем предыдущем исследовании [31]. Прошлые исследования также показали, что взаимодействие с ENP может изменять внутриклеточный Ca 2+ пути, которые необходимы для передачи сигналов клетками [29, 32,33,34]. Специфический внутриклеточный Ca 2+ изменения концентрации важны в процессах передачи сигналов и секреции клеток; однако нет сообщений о диоксиде титана (TiO 2 ), диоксид кремния (SiO 2 ) или диоксид церия (CeO 2 ) для изменения внутриклеточного Ca 2+ уровень фитопланктона.
В 2013 году Quigg et al. [8] обобщили прямые и косвенные токсические эффекты ENP на водоросли. В наших предыдущих экспериментах было показано, что ENP способствуют агрегации EPS [35]. В связи с этим EPS может либо усугубить, либо снизить прямую токсичность, вызванную ENP, по отношению к водным организмам [7, 15, 36]. Однако о прямых измерениях высвобождения САП из фитопланктона в условиях стресса ЕПС никогда не сообщалось. Целью данного исследования является изучение высвобождения ЭПС из четырех различных видов диатомовых водорослей ( Odontella mobiliensis , Skeletonema grethae , Phaeodactylum tricornutum , Thalassiosira pseudonana ) и одна зеленая водоросль ( Dunaliella tertiolecta ) в рамках процедур ЕПС. Понимая основные механизмы вызванного ENP высвобождения ЭПС в фитопланктоне, принятие превентивных мер и мер безопасности может смягчить потенциально пагубное воздействие на морские организмы.
Результаты и обсуждения
Характеристика ENP
Динамическое лазерное рассеяние (DLS) использовалось для характеристики размеров следующих ENP, взвешенных в чистой воде:TiO 2 , SiO 2 , и главный исполнительный директор 2 . Гранулометрический состав TiO 2 составлял от 7 до 66 нм. , От 9 до 66 нм в SiO 2 и от 12 до 70 нм в CeO 2 . Некоторые большие размеры могут быть связаны с агломерацией или агломерацией, в то время как преобладающий размер TiO 2 составляет 25 нм, SiO 2 составляет от 10 до 20 нм, а CeO 2 составляет от 15 до 30 нм, что соответствует информации производителя (рис. 1).
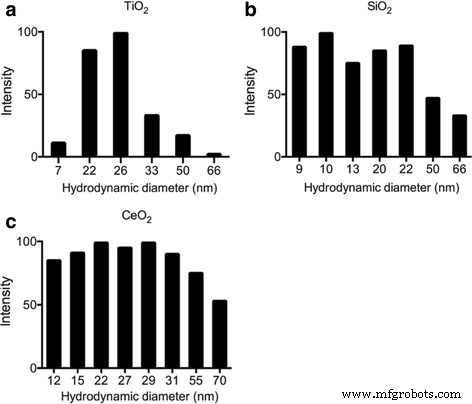
Характеристика ЕПС по оценке DLS a TiO 2 , b SiO 2 , и c Генеральный директор 2 в среде L1 после обработки ультразвуком, показывая их распределение по размерам. Конечная концентрация ENP в образце DLS составляет 1 мкг / мл, время измерения - 3 минуты сразу после обработки ультразвуком
ENPs индуцируют внутриклеточный Ca 2+ Концентрация в фитопланктоне
Изучить, могут ли ENP вызывать повышение внутриклеточного Ca 2+ концентрации, клетки фитопланктона (OD 600 =0,8) загружали красителем Fluo-4AM и подвергали воздействию 1 мг / мл 25 нм TiO 2 , 10–20 нм SiO 2 , и 15–30 нм CeO 2 Соответственно. Изменение внутриклеточного Ca 2+ Концентрация, представленная интенсивностью флуоресценции в клетках фитопланктона, отслеживалась в течение 150 с. На рис. 2a – e показано, что 1 мг / мл трех соответствующих ENP увеличивают Ca 2+ . концентрация в SiO 2 примерно на 50–300%, TiO 2 примерно на 40%, а CeO 2 примерно на 150–200% при неизменных условиях контроля (среда L1). Результаты показывают, что ENP могут индуцировать значительный внутриклеточный Ca 2+ ответы фитопланктона и предполагают, что фитопланктон реагирует на различные ENP через Ca 2+ сигнальные пути. Наши данные указывают только на незначительные изменения внутриклеточного Ca 2+ уровни, когда TiO 2 присутствует, что может быть связано со значительной гибелью клеток фитопланктона от TiO 2 -индуцированная токсичность [37, 38]. В нашем предыдущем исследовании TiO 2 вызывает повышение внутриклеточного Ca 2+ концентрации [34] наряду со значительным апоптозом клеток [39]. Однако SiO 2 неожиданно показал наиболее очевидный внутриклеточный Ca 2+ увеличивается для всех видов фитопланктона, а CeO 2 может запускать только промежуточный внутриклеточный Ca 2+ повышение концентрации. Предыдущее исследование показало потенциал высоких CeO 2 концентрации (> 50 мг / мл) для индукции внутриклеточного окислительного стресса и повышения внутриклеточного Ca 2+ уровни, хотя эффекты были небольшими, и подтвердили наш вывод [40]. Мы также измерили дзета-потенциал каждого ENP в искусственной морской воде, чтобы устранить потенциальный эффект, который может вызвать поверхностный заряд; однако стоимость была низкой. Измерения показали, что ЕПС считаются приблизительно нейтральными [41] (Дополнительный файл 1:Дополнительные данные). Это послужило первым отчетом, в котором было обнаружено, что разные ENP индуцируют внутриклеточный Ca 2+ концентрация изменяется в конкретном фитопланктоне, что в конечном итоге открывает новые возможности для будущих исследований.
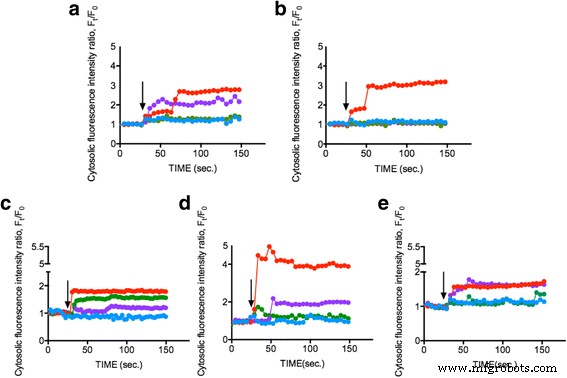
Измерение внутриклеточного Ca 2+ концентрация после стимуляции разными ENP. Различные клетки фитопланктона a Dunaliella tertiolecta , b Thalassiosira pseudonana , c Skeletonema grathae , d Phaeodactylum tricornutum и e Odontella mobiliensis были обработаны TiO 2 25 нм (зеленый), SiO 2 10–20 нм (красный), CeO 2 15–30 нм (фиолетовый) с концентрацией 1 мг / мл и контроль (синий). Черная стрелка указывает момент времени, когда были применены EPN (30 с). Измерения показывают репрезентативные данные в среднем по 20 отдельным ячейкам
Выбросы ЭПС в фитопланктоне, вызванные ЕПД
Ферментно-связанный анализ лектина (ELLA) был использован для оценки количества высвобождения EPS из клеток фитопланктона при стимуляции TiO 2 , SiO 2 , и главный исполнительный директор 2 ENP, диапазон концентраций от 1 мкг / мл до 5 мг / мл на основе предыдущих исследований для TiO 2 [42, 43] и главный исполнительный директор 2 [44,45,46]. Секреция EPS была нормализована к общему количеству ДНК фитопланктона (дополнительный файл 1:дополнительные данные), чтобы иметь равную базу для сравнения. По сравнению с контролем мы обнаружили, что 10–20 нм SiO 2 может увеличить выпуск EPS до 550% в Dunaliella , 500% в Thalassiosira , 1000% в Skeletonema , 400% в Odontella и 900% в Phaeodactylum (Рис. 3). Когда виды фитопланктона подвергались воздействию TiO 2 , сильного влияния на секрецию ЭПС не наблюдалось, так как только Skeletonema и Phaeodactylum показали существенные изменения. Таким образом, данные о выпуске EPS согласуются с нашим внутриклеточным Ca 2+ результаты концентрации. TiO 2 не оказывает значительного влияния на продукцию EPS, подобно тому, как внутриклеточный Ca +2 концентрации показали очень предельные изменения из-за токсичности TiO 2 к фитопланктону. Производство и остатки АФК могут привести ко многим осложнениям, таким как апоптоз фитопланктона [47,48,49]. В СЕО 2 лечение, результаты показали незначительный эффект у Dunaliella , Скелетонема , Одонтелла и Phaeodactylum . Однако SiO 2 показал наиболее значительную индукцию EPS у Thalassiosira pseudonana (около 600%) и Skeletonema grethae (около 1000–1500%). Эти данные показывают, что разные ENP могут вызывать специфическое высвобождение EPS из фитопланктона и внутриклеточного Ca 2+ изменения также соответствуют результатам выпуска EPS. Оценивая изменения внутриклеточного Ca 2+ концентрации, очевидно, что существует прямая связь в Ca 2+ клеточные пути, в которых ENPs вызывают секрецию EPS из фитопланктона. Это наблюдение согласуется с нашими предыдущими исследованиями, основанными на Phaeocystis Выпуск EPS [31]. Результаты являются прямым доказательством того, что фитопланктон может обнаруживать и различать ENP, которые реагируют различным высвобождением EPS, регулируемым Ca 2+ клеточные пути.
Использование ELLA позволило нам определить высвобождение EPS через взаимодействия фитопланктона с ENP. Наши результаты показывают, что секреция EPS значительно увеличилась, поскольку фитопланктон взаимодействовал с SiO 2 для Dunaliella tertiolecta , Thalassiosira pseudonana и Skeletonema grethae . Похоже, что эти диатомовые водоросли способны распознавать SiO 2 частицы. Однако у Phaeodactylum tricornutum , сильной секреции ЭПС не обнаружено. Эта разница представляет собой высвобождение ЭПС, вызванное зависимостью ЭПС от видов фитопланктона и концентрации ЭПС (рис. 3). В предыдущем исследовании разливы нефти вызывали крупные выбросы морских микробов EPS, которые были предложены для противодействия негативным последствиям разливов нефти [50]. Кроме того, Боглаенко и Тансель обнаружили, что SiO 2 частицы были способны эффективно удалять масляные агрегаты [51]. Наше открытие обеспечивает новый потенциальный механизм, в котором SiO 2 с низкой токсичностью частицы могут вызывать высвобождение EPS из определенного фитопланктона, потенциально способствуя удалению разливов нефти за счет агрегации EPS. Никогда не сообщалось, что диоксид церия нарушает морские экосистемы, основанные на фитопланктоне. Результаты здесь показали CeO 2 ЕПС могут повлиять на весь фитопланктон здесь, за исключением Thalassiosira pseudonana. Генеральный директор 2 ЕПС могут, как SiO 2, обладают способностью увеличивать выбросы EPS из определенного фитопланктона для смягчения воздействия нефти.
Выводы
Взаимодействие ЕПС и морской среды становится все более важным из-за текущих и будущих выбросов наноматериалов. Здесь мы демонстрируем усиление секреции ЭПС как одного из основных эффектов ЭПС на фитопланктон. Мы также предоставляем доказательства того, что разные фитопланктоны могут по-разному реагировать на различные стрессы ENP, регулируя Ca 2+ пути. Однако для полной оценки ЕПС в отношении морской экосистемы потребуются дальнейшие исследования, чтобы предоставить подробные знания и понимание взаимодействий между наноматериалами и морскими организмами.
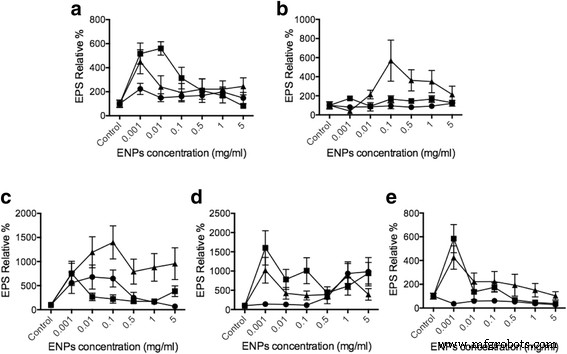
Выпуск EPS вызван различными ENP. Различные клетки фитопланктона a Dunaliella tertiolecta , b Thalassiosira pseudonana , c Skeletonema grathae , d Phaeodactylum tricornutum и e Odontella mobiliensis были обработаны TiO 2 (кружки), SiO 2 (треугольники), CeO 2 (квадраты) соответственно с концентрациями 5 мг / мл и 1 мг / мл, 0,5 мг / мл, 0,1 мг / мл, 10 мкг / мл, 1 мкг / мл ( n =3)
Методы
Культура фитопланктона
Пакетные культуры Odontella mobiliensis (CCMP597), Dunaliella tertiolecta . (UTEX999), Skeletonema grethae (CCMP775), Phaeodactylum tricornutum (UTEX646), Thalassiosira pseudonana (Коллекция культур морского фитопланктона Provasoli - Guillard, West Boothbay Harbor, MN, США) выращивали в морской среде L1 (Sigma, MO, USA) в цикле 14:10 (свет:темнота) при 100 мкмоль м -2 s −1 и 24 ° C в аксенических условиях. Фазу роста культуры определяли путем подсчета клеток с помощью гемоцитометра.
Наночастицы и их характеристика
Все ЕПС, TiO 2 , SiO 2 , Главный исполнительный директор 2 (Sigma-Aldrich, Миссури, США) перед использованием обрабатывали ультразвуком в чистой воде. Перед тестированием ENP были восстановлены фильтрованной средой L1 (Sigma, Миссури, США). Размер ENP был независимо подтвержден с помощью гомодинного динамического лазерного рассеяния (DLS). Вкратце, образцы морской воды повторно фильтровали через мембрану Millipore 0,22 мкм (предварительно промытую 0,1 н. HCl) и выливали непосредственно в пять 10-мл рассеивающих ячеек, которые затем помещали в гониометр лазерного спектрометра Brookhaven BI-200SM (Brookhaven Instruments, Нью-Йорк, США). Автокорреляционная функция флуктуаций интенсивности рассеяния, обнаруженных под углом 45 °, обрабатывалась в режиме реального времени автокоррелятором Brookhaven BI 9000AT, а распределение частиц по размерам рассчитывалось методом CONTIN (Provencher, 1982). Результаты для каждого образца были собраны в трех экземплярах сразу после обработки ультразвуком. Калибровку спектрометра DLS проводили с использованием стандартных суспензий монодисперсных латексных микросфер (Polysciences, PA, США).
Режим ЕПС
Клетки фитопланктона культивировали в 96-луночном планшете со средой L1 в течение 24 ч. Клетки обрабатывали исходными растворами ENP:5 мг / мл и 1 мг / мл, 0,5 мг / мл, 0,1 мг / мл, 10 мкг / мл, 1 мкг / мл TiO 2 , SiO 2 , и главный исполнительный директор 2 (Sigma-Aldrich, Миссури, США) или среду L1 (контроль) в течение 48 часов. Супернатант, содержащий секретированный EPS, собирали и кратко центрифугировали при 4000 об / мин для удаления остаточных ENP. Этот протокол был адаптирован из нашей предыдущей публикации [34]. Диапазон концентраций, используемый здесь, не предназначен для представления или имитации текущих уровней ENP в окружающей среде, но направлен на оценку полного потенциального воздействия ENP на морской фитопланктон и исследование связанных клеточных механизмов. В качестве многообещающего появляющегося наноматериала ЕПС еще не достигли своей полной коммерческой мощности. Подробная оценка их полного экологического воздействия крайне необходима до того, как ЕПС выйдут на рынок коммерческих товаров и товаров для дома, чтобы вывести больше ЕПС в океан.
Ферментно-связанный анализ лектина (ELLA)
Супернатант, содержащий секретированный полисахарид, собирали и ненадолго центрифугировали при 1700 rcf (Megafuge 1.0R) для удаления остаточных ENP. Затем супернатант инкубировали в 96-луночном планшете (Nunc MaxiSorp, VWR, CA, USA) в течение ночи при 4 ° C. Затем 96-луночный планшет промывали PBST (PBS + 0,05% Tween-20) и PBS, а затем блокировали 1% BSA. 96-луночный планшет снова промывали PBST и PBS и инкубировали с лектином (Конканавалин A, ConA) (Sigma-Aldrich, Миссури, США), конъюгированным с пероксидазой хрена (HRP; 5 мг / мл) (Sigma-Aldrich, Миссури). , США), при 37 ° С в течение 1 ч. Субстрат, 3,39,5,59-тетраметилбензидин (TMB; Sigma-Aldrich, Миссури, США), добавляли в каждую лунку при комнатной температуре с последующим добавлением H 2 SO4 (Sigma-Aldrich, Миссури, США) для прекращения реакции. Оптическую плотность измеряли при 450 нм на PerkinElmer VICTOR3 (Массачусетс, США). Этот протокол был адаптирован из нашей предыдущей публикации [34, 52].
Определение ДНК
Осадок, содержащий фитопланктон, собирали и получали набор ZR-96 Quick-gDNA (ZYMO Research, Калифорния, США). Вкратце, для разрушения клеток фитопланктона и протекания через колонку для связывания ДНК использовали 4-кратный буфер для лизиса, в конце которого элюировали элюирующим буфером. Концентрации ДНК измеряли с помощью NanoDrop ND-1000 (Thermo, CA, USA). Протокол был адаптирован из протокола изготовленного набора.
Измерения внутриклеточного Ca 2+ Концентрации, вызванные ЕПС
Затем клетки фитопланктона загружали красителем Fluo-4AM (1 мМ) (Kd =335 нМ, λEx =494 нм и λEm =506 нм, ThermoFisher, CA, USA) в течение 60 мин [31]. После нанесения красителя клетки фитопланктона промывали, инкубировали со средой L1 и обрабатывали 1 мг / мл TiO 2 , SiO 2 , и главный исполнительный директор 2 соответственно. Все эксперименты по передаче сигналов кальция проводились на микроскопе Nikon (Nikon Eclipse TE2000-U, Токио, Япония). Протокол и условия были адаптированы из предыдущих публикаций [31, 34].
Дзета-потенциал измерения ЕПС
Для измерения поверхностных зарядов ENP дзета-потенциал (ζ) ENP измеряли с помощью Zetasizer Nano ZS, Malvern в присутствии искусственной морской воды при 25 ° C. После сбора данных для каждого образца зарегистрированные значения были усреднены.
Статистический анализ
Данные представлены в виде средних значений ± стандартное отклонение. Каждый эксперимент проводился независимо не менее трех раз. Гистограммы построены с помощью GraphPad Prism 6.0. (GraphPad Software, Inc., Сан-Диего, Калифорния, США).
Наноматериалы
- Полупроводниковые наночастицы
- Плазмонные наночастицы
- Влияние pH на желтые красители из сада
- Механизм образования упорядоченных плотно упакованных сверхрешеток наночастиц, отложенных из газовой фазы …
- Наночастицы как насос оттока и ингибитор биопленки для омоложения бактерицидного действия обычных антибиот…
- Биофлавоноиды, содержащие генистеин, наночастицы хитозана, нацеленные на фолатные рецепторы, для усиления пр…
- Нанотехнологии:от системы визуализации in vivo к контролируемой доставке лекарств
- Влияние метода синтеза наночастиц манганита La1 - xSr x MnO3 на их свойства
- Токсичность наночастиц CoFe2O4, покрытых ПЭГ, с лечебным эффектом куркумина
- Влияние кислотного пептизирующего агента на соотношение анатаз-рутил и фотокаталитические характеристики н…