


Понижающая регуляция микроРНК-342-5p или повышающая регуляция Wnt3a ингибирует ангиогенез и поддерживает стабильность атеросклеротических бляшек у мышей с атеросклерозом
Аннотация
Доказательства продемонстрировали, что микроРНК-342-5p (miR-342-5p) участвует в атеросклерозе (AS), но мало что известно о его внутренних регуляторных механизмах. Здесь мы стремились изучить влияние miR-342-5p, нацеленного на Wnt3a, на образование уязвимых бляшек и ангиогенез AS. ApoE - / - мышей кормили кормом с высоким содержанием жира в течение 16 недель, чтобы воспроизвести модель уязвимых бляшек AS. Выявлена экспрессия miR-342-5p и Wnt3a в тканях аорты при AS. Целевое отношение между miR-342-5p и Wnt3a было подтверждено. Более того, ApoE - / - Мышам вводили антагомир miR-342-5p и вектор сверхэкспрессии Wnt3a для проверки их функций в отношении уровней липидов в сыворотке, цитокинов, связанных с воспалительным и окислительным стрессом, стабильности аортальных бляшек и ангиогенеза в бляшках мышей AS. Экспрессия miR-342-5p была усилена, а экспрессия Wnt3a деградировала в тканях аорты мышей AS, а miR-342-5p непосредственно нацелена на Wnt3a. Повышающая регуляция Wnt3a или понижающая регуляция miR-342-5p снижали содержание липидов в крови, уровни воспалительного и окислительного стресса, уязвимость бляшки ткани аорты и ингибировали ангиогенез в бляшке аорты мышей AS. Функциональные исследования показывают, что истощение miR-342-5p может стабилизировать бляшки ткани аорты и уменьшить ангиогенез в бляшках у мышей AS за счет восстановления Wnt3a.
Введение
Атеросклероз (АС) - это возрастное заболевание артерий, характеризующееся утолщением, стенозом, затвердением и образованием атеросклеротических бляшек артерий [1]. Это основная причина смерти и заболеваемости в развитых странах [2]. Гистопатологические исследования атеросклеротических поражений человека показали, что развитие и разрыв бляшек характеризуются увеличением липидного / некротического ядра, уменьшением количества гладкомышечных клеток, инфильтрацией макрофагов и снижением содержания коллагена [3]. Ключевые клеточные элементы AS включают гиперлипидемию, образование пенистых клеток, дифференцировку в макрофаги, привлечение моноцитов и индуцированное воспаление [4]. Хотя многие лекарства для лечения АС широко используются в клинике, некоторые подгруппы пациентов все еще подвержены высокому риску инфаркта миокарда, ишемии миокарда, сердечной недостаточности и инсульта [5]. Следовательно, дальнейшее изучение потенциальных молекулярных механизмов может предоставить больше доказательств для лечения АС.
Одна микроРНК (miRNA) может одновременно регулировать несколько генов-мишеней [6]. miR-342-5p, как выяснилось, содержится в импринтированном кластере miRNA 14q32, действует как инновационная молекула Notch, расположенная ниже по течению [7], и модулирует множественные ангиогенные пути, такие как трансформирующий передачу сигналов фактора роста β и фактор роста эндотелия сосудов [8]. Родственные иммуномодулирующие микроРНК, такие как miR-342-5p, играют множество важных ролей в регуляции прогрессирования атеросклероза [9]. Более того, предполагается, что некоторые miRNAs участвуют в разрешении AS, такие как miR-155 и miR-217 [10, 11]. В исследовании сообщается, что miR-342-5p действует как новый модулятор активации макрофагов при AS [12]. Другое исследование показало, что происходящая из макрофагов miR-342-5p способствует AS и увеличивает воспалительную стимуляцию макрофагов [13]. Ку и др. . обнаружили, что экспрессия Wnt3a отрицательно модулируется miR-342-5p при аноректальных мальформациях [14], указывая тем самым, что существует целевое отношение между miR-342-5p и Wnt3a. Передача сигналов Wnt играет важную роль во время эмбриогенеза для модуляции клеточной полярности, клеточной пролиферации, формирования оси и апоптоза [15]. Wnt3a, ключевой компонент гена мезодермы, играет критическую роль в эмбриональном развитии [16]. Было показано, что анализ транскриптома бляшек макрофагов под контролем эпигенома во время регрессии AS обнаруживает активацию пути передачи сигналов Wnt [17]. Более того, исследование показало, что Wnt3a модулирует адгезию и миграцию гладкомышечных клеток сосудов, которые вносят вклад в патогенез AS и рестеноза [18]. Таким образом, в этом исследовании впервые изучалось влияние miR-342-5p, нацеленного на Wnt3a, на образование уязвимых бляшек и ангиогенез AS.
Материалы и методы
Этика
С животными обращались гуманно с использованием утвержденных процедур в соответствии с рекомендациями Руководства по уходу и использованию лабораторных животных Национального института здоровья. Протокол был одобрен Комитетом по уходу за животными и их использованию Народной больницы провинции Цинхай (этический номер:201870726).
Экспериментальные животные
Мужской ApoE - / - мышей и мышей C57BL / 6J (степень отсутствия определенных патогенов) в возрасте 8 недель были доступны в компании Beijing Vital Laboratory Animal Technology (Пекин, Китай). Мышей (5–6 мышей в клетке) содержали с 12-часовым / 12-часовым циклом день / ночь с неограниченным доступом к пище и воде.
Создание мышей-моделей AS
ApoE - / - мышей кормили кормом с высоким содержанием жира в течение 16 недель для создания модели бляшек, уязвимых к AS. Мышей C57BL / 6 J использовали в качестве нормальной группы с натуральными напитками и едой. ApoE - / - у мышей были блестящие светлые волосы и выпадение волос на спине через 12 нед. Дуга аорты и брахиоцефальная артерия у 3 модельных мышей были рассечены для окрашивания гематоксилином-эозином (HE), и на интиме не было значительных отложений бляшек. Еще 3 моделированные мыши были идентифицированы снова через 4 недели, и окрашивание HE показало, что на интиме дуги аорты были очевидные отложения бляшек, что указывает на успех создания модели.
Группировка и лечение мышей
ApoE - / - Мыши с уязвимой к AS бляшкой были разделены на 6 групп по 12 мышей в каждой группе:группа AS, группа отрицательного контроля (NC) (которым вводили физиологический раствор в ApoE - / - мышей), группа агомира miR-342-5p (инъецированная агомиром miR-342-5p для сверхэкспрессии экспрессии miR-342-5p в ApoE - / - мышей), группа антагомира miR-342-5p (инъецированная антагомиром miR-342-5p для снижения экспрессии miR-342-5p в ApoE - / - мышей), группа сверхэкспрессии (oe) -Wnt3a (инъецированная вектором oe-Wnt3a для усиления экспрессии Wnt3a в ApoE - / - мышей) и группа miR-342-5p агомир + oe-Wnt3a (инъецированная агомиром miR-342-5p и вектором oe-Wnt3a для усиления экспрессии miR-342-5p и Wnt3a в ApoE - / - мышей). Мышей C57BL / 6 J как нормальной группе кормили нормальной диетой. Корм с высоким содержанием жира содержал 20% жира и 0,25% холестерина. Агомир miR-342-5p, антагомир miR-342-5p и вектор oe-Wnt3a были приобретены в Sangon (Шанхай, Китай). Вектор oe-Wnt3a, агомир miR-342-5p, антагомир miR-342-5p растворяли в 0,2 мл физиологического раствора и вводили мышам в дозе 40 мг / кг через хвостовую вену каждые две недели. Через 8 недель были взяты образцы крови из глазных яблок, а затем мышей умерщвляли для сбора артериальных тканей [19].
В предварительном эксперименте ApoE - / - мышам с AS вводили 10 мг / кг, 20 мг / кг, 40 мг / кг агомира miR-342-5p, антагомира miR-342-5p или вектора oe-Wnt3a (один раз в две недели; всего 4 раза). Затем уровни экспрессии β-катенина определяли с помощью количественной полимеразной цепной реакции с обратной транскрипцией (RT-qPCR).
Сбор и обработка образцов
Перед взятием образцов мышей не кормили в течение 12 часов и анестезировали ингаляцией эфира, а образцы крови собирали из глазных яблок. Грудь мышей вскрывали, грудную аорту отделяли до конца брюшной аорты и удаляли весь сосуд. После очистки свободным от РНК физиологическим раствором с фосфатным буфером (PBS) ткани заключали для окрашивания HE, масляного красного O, окрашивания красным Сириусом и иммуногистохимического окрашивания. Некоторые сосудистые ткани сохраняли при -80 ° C для RT-qPCR и вестерн-блоттинга.
Определение уровня липидов в крови
Автоматический биохимический анализатор (Roche, Базель, Швейцария) был адаптирован для определения общего холестерина (TC), триглицеридов (TG), холестерина липопротеинов низкой плотности (LDL-C) и холестерина липопротеинов высокой плотности (HDL-C) в сыворотке. Обнаружение было реализовано в соответствии со спецификацией наборов (NanJing JianCheng Bioengineering Institute, Нанкин, Китай).
Иммуноферментный анализ (ELISA)
Определение содержания цитокинов в сыворотке:использовались наборы ELISA для коммерческого интерлейкина (IL) -5, IL-12p70, фактора некроза опухоли альфа (TNF-α) и интерферона (IFN) -γ. Наконец, значение оптической плотности (OD) каждой лунки было протестировано с помощью считывающего устройства для микропланшетов при 450 нм.
Определение повреждения, вызванного окислительным стрессом:содержание малонового диальдейда (MDA) и активность супероксиддисмутазы (SOD) в сыворотке крови проверяли с помощью набора MDA (значение OD проверяли на спектрофотометре при 532 нм) и набора SOD (значение OD определяли с помощью считывающего устройства для микропланшетов при 450 нм). нм). Наборы ИФА для IL-5, IL-12p70, TNF-α, IFN-γ, MDA и SOD были приобретены у MultiSciences (Lianke) Biotechnology Corporate Limited (Ханчжоу, Чжэцзян, Китай).
Окрашивание HE, окрашивание Oil Red O и окрашивание Sirius Red
После фиксации и заделки образцы разрезали на последовательные срезы толщиной 4 микрометра. Срезы депарафинизировали и гидратировали, окрашивали гематоксилином и эозином, дифференцировали, дегидратировали, очищали ксилолом, сушили и герметизировали нейтральной камедью. Ядро было синим, а другие ткани, такие как цитоплазма и соединительные ткани, были красными разных оттенков. Образование налета наблюдали с помощью флуоресцентного микроскопа. Стенки артерий срезов, окрашенных HE, отбирали под микроскопом, а результаты экспериментов собирали с помощью цифровой камеры. Программный модуль анализа изображений Image Pro Plus6.0 (IPP6.0) использовался для расчета площади налета в поперечном сечении каждого среза и площади стены, а также их соотношения.
Для окрашивания О масляным красным отбирали срезы размером 4–5 мкм. Срезы сушили при избыточной температуре в течение 20 мин и инкубировали со 100% изопропанолом в течение 5 мин. Затем срезы инкубировали с 0,5% -ным окрашивающим раствором масляным красным О в печи при 60 ° C в течение 8 минут, промывали в 85% изопропаноле в течение 3 минут, окрашивали гематоксилином в течение 1 минуты, очищали и герметизировали. Результаты окрашивания масляным красным О показали, что липид был красным или оранжевым, а ядро было светло-голубым. Программное обеспечение IPP6.0 использовалось для расчета площади жировой ткани и площади бляшки в бляшке среза ткани. Содержание липидов =O-положительная область окрашивания масляным красным / площадь бляшки × 100%.
Окрашивание сириусом красным:срезы депарафинизировали и гидратировали, окрашивали в течение 10 минут окрашивающим раствором целестинового синего, окрашивали раствором сириус-красного в течение 20 минут и контрастировали в течение 10 минут гематоксилином. Наконец, срезы обезвоживали градиентом этанола, очищали ксилолом и герметизировали нейтральной смолой. Площадь коллагена в бляшке среза ткани рассчитывалась с помощью программного обеспечения IPP6.0. Площадь коллагена =площадь положительного окрашивания по Сириусу красным / площадь бляшки × 100%. Был рассчитан процент липидов и коллагена в зоне бляшки.
Гематоксилин, эозин и краситель Сириус были доступны от China Pharmaceutical Group Shanghai Chemical Reagent Co. Ltd. (Шанхай, Китай). Масляный красный порошок O был куплен у Sigma-Aldrich Chemical Company (Сент-Луис, Миссури, США).
Количественная полимеразная цепная реакция с обратной транскрипцией (RT-qPCR)
Ткани аорты добавляли к реагенту для экстракции общей РНК Trizol (Invitrogen, Carlsbad, California, USA), а затем гомогенизировали для экстракции общей РНК и комплементарной ДНК. Все праймеры ссылались на последовательность, предоставленную Genbank, разработанную с помощью Primer 5.0 и синтезированную Shanghai Sangon Biotechnology Co. Ltd. компания, Хильден, Германия); Wnt3a:вперед:5'-AGGTAAGCTACTCCCTCAACTA-3 ', назад:5'-CTGAAGCACCCTCTCATGTATC-3'; -актин:прямой:5'-GCACCACACCTTCTACAATGAGC -3 ', обратный:5'-TCGTTGCCAATAGTGATGACC-3'; β-катенин:вперед:5'-TCAAGAGAGCAAGCTCATCATTCT-3 ', назад:5'-CACCTTCAGCACTCTGCTTGTG-3'. После реакции компьютер анализировал пороговый цикл (Ct). Относительное отношение miR-342-5p к U6 использовалось в качестве его выражения, относительное отношение Wnt3a к β-актину использовалось в качестве его выражения, и относительное соотношение было рассчитано как 2 −ΔΔCt метод.
Вестерн-блоттинг
Общий белок извлекали из тканей аорты. Концентрацию белка измеряли методом бицинхониновой кислоты. Был проведен электрофорез в полиакриламидном геле. Затем белок переносили на мембрану из поливинилиденфторида и получали целевую полосу. Мембрану герметично закрывали в 5% обезжиренном молоке на 1 ч, добавляли первичные антитела Wnt3a (1:500), β-катенин (1:1000, Santa Cruz Biotechnology, Inc., Санта-Крус, Калифорния, США), CD34 (1 :2500, Abcam, MA, США) и β-актина (1:2000, Beyotime Biotechnology Co., Шанхай, Китай) при 4 ° C в течение ночи. Мембрану промывали трис-буферным физиологическим раствором с Твин 20 (pH =7,5, 10 ммоль / л Трис-HCl, 100 ммоль / л NaCl и 0,2% Твин-20) в течение 10 мин × 3 раза, а затем добавляли вторичное антитело. (1:1000, ZSGB-Bio, Пекин, Китай) на 2 ч. Программное обеспечение ImageJ было адаптировано для оценки значения серого в полосах и количественного определения экспрессии белка.
Иммуногистохимическое окрашивание
Срезы размером 4–5 мкм помещали на предметные стекла, покрытые полилизином 100 мг / л и фиксированные ацетоном. Эндогенная пероксидаза блокировалась бычьим сывороточным альбумином. На ткани капали антитело MOMA-2 (1:200), α-SMA (1:200) и CD34 (1:200, Abcam Inc., Кембридж, Массачусетс, США), а также добавляли рабочий раствор вторичного антитела ( 1:1000). Ткани проявляли диаминобензидином, контрастировали гематоксилином (1 мин), обезвоживали, очищали, герметизировали и наблюдали под микроскопом. Для каждого иммуногистохимического среза были выбраны три различных поля зрения. Программное обеспечение IPP6.0 было выполнено для количественного анализа. Положительное иммуногистохимическое окрашивание MOMA-2 и α-SMA, соответственно, указывает на то, что макрофаги и гладкомышечные клетки в основном расположены в цитоплазме от желтого до коричневого цвета. Отдельно рассчитывали процентное содержание макрофагов и гладкомышечных клеток, которые объединяли с процентным содержанием липидов и коллагена в бляшке для расчета индекса уязвимости бляшки. Индекс уязвимости бляшки =(положительный процент макрофагов + положительный процент липидов) / (положительный процент коллагена + положительный процент гладкомышечных клеток) [20]. Плотность микрососудов (MVD) оценивали путем измерения экспрессии CD34 и количественно определяли как количество микрососудов / мм 2 .
Анализ двойного люциферазного репортерного гена
Целевой ген miR-342-5p был проанализирован на сайте биологического прогнозирования (http://www.microRNA.org). Двойной анализ репортерного гена люциферазы использовали для проверки того, является ли Wnt3a целевым геном для miR-342-5p. Последовательность дикого типа или мутантную 3'-нетранслируемую область Wnt3a (3'-UTR) клонировали в вектор GP-miRGLO (GenePharma, Шанхай, Китай). Репортер (0,5 мкг) и агомир 1, 10 или 100 пМ miR-342-5p трансфицировали в эндотелиальные клетки аорты мыши (№ 506, MingzhouBio, Нинбо, Китай) на 48 часов для проверки активности люциферазы с использованием двойной системы анализа люциферазы. (Промега, Висконсин, США).
Статистический анализ
Все данные были интерпретированы с помощью программного обеспечения SPSS 21.0 (IBM Corp. Armonk, NY, США). Данные измерений были указаны как среднее ± стандартное отклонение. Различия между двумя группами были сформулированы t -тест, а для нескольких групп - односторонний дисперсионный анализ (ANOVA) с последующим тестом множественных сравнений Тьюки. Статистическая значимость была установлена P значение <0,05.
Результаты
miR-342-5p увеличивает и уменьшает Wnt3a в тканях аорты ApoE - / - мыши и miR-342-5p напрямую нацелены на Wnt3a
Гены-мишени микроРНК (miRNA) связаны с функциями, связанными с атеросклерозом. miR-342-5p, Wnt3a и β-катенин тестировали в тканях аорты мышей модели AS с помощью RT-qPCR и вестерн-блоттинга. Было обнаружено, что по сравнению с нормальной группой miR-342-5p был повышен, в то время как Wnt3a и β-катенин были снижены в группе AS (оба P <0,05). По сравнению с группой NC, miR-342-5p была усилена, а Wnt3a и β-катенин уменьшены в группе агомира miR-342-5p (обе P <0,05), в то время как miR-342-5p был снижен, Wnt3a и β-катенин были повышены в группе антагомира miR-342-5p (оба P <0,05). Экспрессия Wnt3a и β-катенина была повышена в группе oe-Wnt3a по сравнению с группой NC (обе P <0,05). По сравнению с группой агомира miR-342-5p, экспрессия Wnt3a и β-катенина была повышена в группе агомира miR-342-5p + oe-Wnt3a ( P <0,05) (рис. 1A – D). Кроме того, в предварительном эксперименте была протестирована экспрессия β-катенина при обработке различных концентраций агомира miR-342-5p, антагомира miR-342-5p и oe-Wnt3a, и результаты показали (дополнительный файл 1:рис. S1 ) чем выше концентрация агомира miR-342-5p, тем ниже экспрессия β-катенина; чем выше концентрация антагомира miR-342-5p, тем выше экспрессия β-катенина; и чем выше концентрация oe-Wnt3a, тем выше экспрессия β-катенина.
miR-342-5p увеличивается, а Wnt3a уменьшается в тканях аорты ApoE - / - мыши и miR-342-5p непосредственно нацелены на Wnt3a. А Экспрессия miR-342-5p в ткани аорты мышей обнаружена с помощью RT-qPCR. Б Экспрессия мРНК Wnt3a в ткани аорты мышей, обнаруженная с помощью RT-qPCR ( n =12). В , D Экспрессия белков Wnt3a и β-катенина в тканях аорты мышей, протестированных с помощью вестерн-блоттинга ( n =12). E Сайт связывания miR-342-5p в 3'-UTR Wnt3a. F агомир miR-342-5p дозозависимо снижал относительную активность в клетках, трансфицированных 3'-UTR Wnt3a ( N =3). G Относительная активность люциферазы в клетках с 3'-UTR Wnt3a дикого типа и мутантным после трансфекции агомиром miR-342-5p или скремблом ( N =3). * P <0,05 по сравнению с нормальной группой, # P <0,05 по сравнению с группой NC. & P <0,05 по сравнению с группой агомира miR-342-5p. Данные измерений были указаны как среднее ± стандартное отклонение. Сравнения между двумя группами были сформулированы t -тест, в то время как сравнения между несколькими группами оценивали с помощью однофакторного дисперсионного анализа с последующим тестом множественных сравнений Тьюки. АС - атеросклероз; NC, отрицательный контроль
miRNAs могут ингибировать трансляцию специфических генов путем связывания с их информационной РНК 3'UTR. Сайт Bioinformatics предсказал, что существует целевое отношение между miR-342-5p и Wnt3a (рис. 1E). Анализ двойного репортерного гена люциферазы сообщил, что в эндотелиальных клетках аорты мышей, трансфицированных вектором Wnt3a 3'UTR, значение люциферазы в рениллах / светлячках дозозависимо снижалось при трансфекции агомиром miR-342-5p со значительным снижением с 10 до 100. pM miR-342-5p агомир и 64% снижение произошло в группе агомира 100 pM miR-342-5p по сравнению с группой NC. Это указывает на присутствие сайта-мишени miR-342-5p в 3'UTR Wnt3a. Однако значение активности люциферазы у ренилл / светлячков не изменилось в группе с мутациями Wnt3a (Fig. 1F, G). Таким образом, можно подтвердить, что Wnt3a является прямым геном-мишенью для miR-342-5p, а miR-342-5p / Wnt3a может регулировать прогрессирование AS.
Влияние повышающей регуляции Wnt3a или понижающей регуляции miR-342-5p на уровни липидов в ApoE - / - Мыши
Кроме того, чтобы исследовать, влияет ли нацеливание miR-342-5p и регулирование сигнального пути Wnt3a на уровни липидов у мышей AS, использовали автоматический биохимический анализатор для наблюдения за изменением уровней липидов. Результаты показали, что (рис. 2A – D), в отличие от нормальной группы, содержание TC, TG и LDL-C было повышено, а содержание HDL-C было снижено в группе AS (все P <0,05). По сравнению с группой NC, содержание TC, TG и LDL-C было повышено, а содержание HDL-C было снижено в группе агомира miR-342-5p (все P <0,05), в то время как содержание TC, TG и LDL-C было снижено, а содержание HDL-C было увеличено в группе антагомира miR-342-5p и группе oe-Wnt3a (все P <0,05). Что касается группы агомира miR-342-5p, содержание TC, TG и LDL-C было снижено, а содержание HDL-C было повышено в группе miR-342-5p agomir + oe-Wnt3a (все P <0,05). Эти результаты подтверждают, что miR-342-5p и Wnt3a регулируют уровень липидов в крови мышей AS, и дополнительно иллюстрируют взаимосвязь направленной регуляции между miR-342-5p и Wnt3a. Сверхэкспрессия Wnt3a могла бы обратить вспять эффекты, вызванные сверхэкспрессией miR-342-5p, у мышей AS.
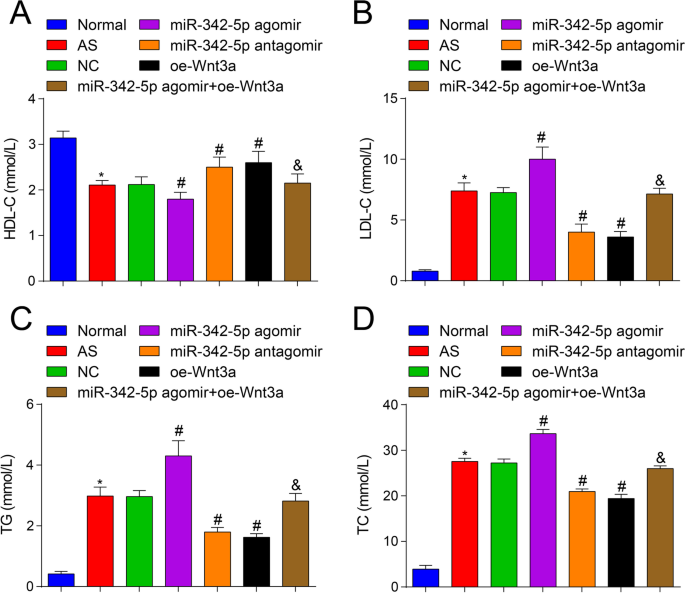
Влияние повышающей регуляции Wnt3a или пониженной регуляции miR-342-5p на уровни липидов в ApoE - / - мышей. А Сравнение содержания ХС-ЛПВП в сыворотке группы мышей. Б Сравнение содержания ХС-ЛПНП в сыворотке крови мышей. В Сравнение содержания ТГ в сыворотке крови мышей. D Сравнение содержания TC в сыворотке крови мышей. нет =12. * P <0,05 по сравнению с нормальной группой, # P <0,05 по сравнению с группой NC. & P <0,05 по сравнению с группой агомира miR-342-5p. Данные измерений были указаны как среднее ± стандартное отклонение. Сравнения между несколькими группами оценивали с помощью однофакторного дисперсионного анализа с последующим тестом множественных сравнений Тьюки. АС - атеросклероз; NC, отрицательный контроль
Влияние сверхэкспрессии Wnt3a или низкой экспрессии miR-342-5p на цитокины, связанные с воспалительным и окислительным стрессом, в сыворотке ApoE - / - Мыши
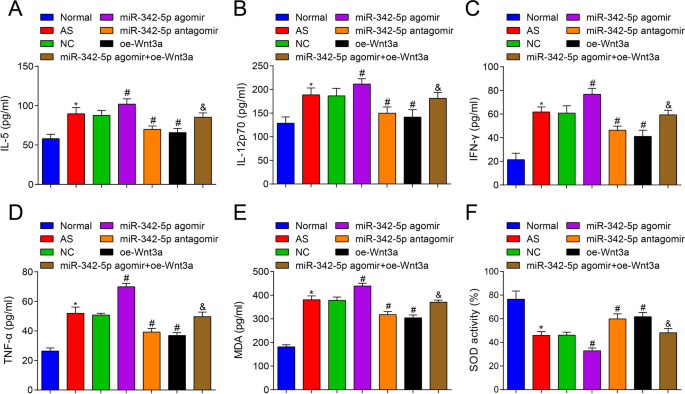
Затем содержание цитокинов в сыворотке мышей AS было протестировано с помощью ELISA, и результаты показали, что (рис. 3A – D) по сравнению с нормальной группой, IL-5, IL-12p70, IFN-γ и TNF-α были увеличены. в группе AS (все P <0,05). По сравнению с группой NC, содержание IL-5, IL-12p70, IFN-γ и TNF-α было повышено в группе агомира miR-342-5p (все P <0,05), тогда как содержание IL-5, IL-12p70, IFN-γ и TNF-α деградировало в группе антагомира miR-342-5p и группе oe-Wnt3a (все P <0,05). В отличие от группы агомира miR-342-5p, содержание IL-5, IL-12p70, IFN-γ и TNF-α было снижено в группе агомира miR-342-5p + oe-Wnt3a (все P <0,05). Намекнули, что нацеленная регуляция miR-342-5p сигнального пути Wnt3a дополнительно регулирует уровень родственных цитокинов в сыворотке мышей AS.
Влияние сверхэкспрессии Wnt3a или низкой экспрессии miR-342-5p на цитокины, связанные с воспалительным и окислительным стрессом, в сыворотке ApoE - / - мышей. А Сравнение содержания ИЛ-5 в сыворотке крови мышей. Б Сравнение содержания IL-12p70 в сыворотке крови мышей. В Сравнение содержания IFN-γ в сыворотке крови мышей. D Сравнение содержания TNF-α в сыворотке крови мышей. E, Сравнение содержания МДА в сыворотке группы мышей. F Сравнение активности СОД в сыворотке крови мышей. нет =12. * P <0,05 по сравнению с нормальной группой, # P <0,05 по сравнению с группой NC. & P <0,05 по сравнению с группой агомира miR-342-5p. Данные измерений были указаны как среднее ± стандартное отклонение. Сравнения между несколькими группами оценивали с помощью однофакторного дисперсионного анализа с последующим тестом множественных сравнений Тьюки. АС - атеросклероз; NC, отрицательный контроль
Кроме того, были протестированы содержание MDA и активность SOD в сыворотке мышей, и было обнаружено, что (рис. 3E, F) по сравнению с нормальной группой содержание MDA было повышено, а активность SOD была снижена в группе AS (оба P <0,05). По сравнению с группой NC, содержание MDA было увеличено, а активность SOD была снижена в группе агомира miR-342-5p (обе P <0,05), в то время как содержание MDA было снижено, а активность SOD была увеличена в группе антагомира miR-342-5p и группе oe-Wnt3a (все P <0,05). По сравнению с группой агомира miR-342-5p, содержание MDA было снижено, а активность SOD была увеличена в группе miR-342-5p агомир + oe-Wnt3a (обе P <0,05). Таким образом, было получено заключение, что истощение miR-342-5p и восстановление Wnt3a ингибируют окислительный стресс у мышей AS.
Влияние истощения miR-342-5p или восстановления Wnt3a на содержание липидов и коллагена в аортальной бляшке ApoE - / - Мыши
Чтобы изучить влияние Wnt3a, нацеленного на miR-342-5p, на площадь бляшек в ткани аорты мышей, было проведено окрашивание HE, и результаты показали, что (рис. 4A, B), за исключением нормальной группы, бляшки AS образовывались во всех разделы других групп. В группе AS площадь бляшки была большой, фиброзный колпачок был тоньше, липидное ядро было увеличено, в бляшке появилось больше пенистых клеток и выпадения кристаллов холестерина, внутренняя стенка артерии и мышечный слой были утолщены, и бляшка была более плотной. нестабильный. Ситуация в группе NC была аналогична таковой в группе AS. В группе miR-342-5p antagomir и oe-Wnt3a площадь бляшки была небольшой, интима артерии была гладкой, а фиброзные капсулы имели небольшое количество и становились тоньше. В бляшке не было разрыва, но наблюдались пенистые ячейки разного размера. Кристалл холестерина был асимметрично распределен и частично кальцифицирован, количество гладкомышечных клеток и коллагеновых волокон увеличивалось, а бляшка имела тенденцию к стабильности. По сравнению с группой NC, площадь бляшки была увеличена, а поражения AS усугубились в группе агомира miR-342-5p ( P <0,05), в то время как площадь бляшки была уменьшена в группе антагомира miR-342-5p и группе oe-Wnt3a с уменьшенными поражениями AS (обе P <0,05). По сравнению с группой агомира miR-342-5p площадь бляшки была уменьшена в группе агомира miR-342-5p + oe-Wnt3a ( P <0,05).
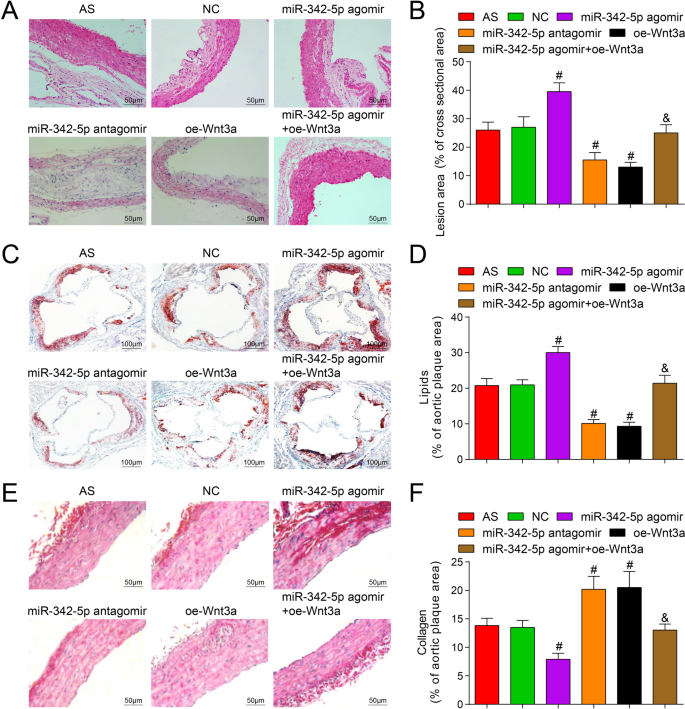
Влияние истощения miR-342-5p или восстановления Wnt3a на содержание липидов и коллагена в бляшке аорты ApoE - / - мышей. А Результаты окрашивания HE аорты у мышей (масштабная линейка:50 мкм). Б Сравнение площади бляшки аорты в каждой группе мышей. В Результаты окрашивания аортальным маслом Red O во всех группах мышей (масштабная линейка:100 мкм). D Сравнение содержания липидов в ткани аорты мышей. E Результаты окрашивания аорты сириусом красным в каждой группе мышей (масштабная линейка:50 мкм). F Сравнение содержания коллагена в тканях аорты мышей. нет =12. # P <0,05 по сравнению с группой NC. & P <0,05 по сравнению с группой агомира miR-342-5p. Данные измерений были указаны как среднее ± стандартное отклонение. Сравнения между несколькими группами оценивали с помощью однофакторного дисперсионного анализа с последующим тестом множественных сравнений Тьюки. АС - атеросклероз; NC, отрицательный контроль
Окрашивание масляным красным О и окрашивание сириус-красным были использованы для обнаружения эффекта miR-342-5p, нацеленного на Wnt3a, на содержание липидов и коллагена в бляшках ткани аорты мышей, и результаты показали, что (рис. 4C – F) масляный красный Окрашивание показало красный жир и синее ядро, а окрашивание Сириус красным показало красные коллагеновые волокна и синее ядро. По сравнению с группой NC, содержание липидов было повышено, а содержание коллагена было снижено в группе агомира miR-342-5p, а также содержание липидов было уменьшено, а содержание коллагена было накоплено в группе антагомира miR-342-5p и oe-Wnt3a. группа (все P <0,05). Что касается группы агомира miR-342-5p, содержание липидов было снижено, а содержание коллагена увеличено в группе агомира miR-342-5p + oe-Wnt3a (обе P <0,05). Результаты экспериментов полностью иллюстрируют, что нацеленная регуляция miR-342-5p сигнального пути Wnt3a оказывает регулирующее влияние на содержание липидов и коллагена в аортальной бляшке мышей AS.
Влияние понижающей регуляции miR-342-5p или повышающей регуляции Wnt3a на макрофаги и гладкомышечные клетки в бляшке аорты ApoE - / - Мыши
Степень АС прямо пропорциональна содержанию одноядерных макрофагов [21]. VSMC являются основными клетками в среднем слое артерий и необходимы для поддержания целостности артериальной стенки. VSMCs участвуют в реконструкции артериальной стенки и играют важную роль в AS на различных стадиях [22]. α-SMA - специфический маркер гладкомышечных клеток [23]. В этом исследовании антитело к маркеру макрофагов (MOMA-2) было использовано для мечения макрофагов, и была применена иммуногистохимия для определения экспрессии MOMA-2 и α-SMA, соответственно.
Под микроскопом положительное иммуногистохимическое окрашивание MOMA-2 и α-SMA, соответственно, указывает на то, что макрофаги и гладкомышечные клетки в основном расположены в цитоплазме от желтого до коричневого цвета. MOMA-2 immune positive indicated that macrophages was mainly located in the cytoplasm with yellow to brown. Determined by immunohistochemistry, it was manifested that versus the NC group, percentage of plaque macrophages (MAMO-2) positive staining was raised and percentage of positive smooth muscle cells was decreased in the miR-342-5p agomir (both P < 0.05). Percentage of plaque macrophages (MAMO-2) positive staining was reduced and percentage of positive smooth muscle cells was increased in the miR-342-5p antagomir group and the oe-Wnt3a group (all P < 0.05). In comparison with the miR-342-5p agomir group, percentage of plaque macrophages (MAMO-2) positive staining was depressed and percentage of positive smooth muscle cells was raised in the miR-342-5p agomir + oe-Wnt3a group (both P < 0.05) (Fig. 5A–D). It was implied that miR-342-5p targeted regulation of Wnt3a signaling pathway could regulate aggregation of macrophages and smooth muscle cells in arterial tissue plaques of AS mice.

Effects of high expression of Wnt3a or poor expression of miR-342-5p on aortic plaque vulnerability of ApoE −/− мышей. A Immunohistochemical staining of MOMA-2 in each group of mice (scale bar:50 μm). B Quantitative analysis of figure A. C Immunohistochemical staining of α-SMA in each group of mice (scale bar:50 μm). D Quantitative analysis of figure C . E Comparison of plaque vulnerability index in aortic tissues of AS mice. n = 12. # P < 0.05 vs. the NC group. &P < 0.05 vs. the miR-342-5p agomir group. Measurement data were indicated as mean ± standard deviation. Comparisons among multiple groups were assessed by one-way ANOVA followed by Tukey’s multiple comparisons test. AS, atherosclerosis; NC, negative control
Effects of High Expression of Wnt3a or Poor Expression of miR-342-5p on Aortic Plaque Vulnerability of ApoE −/− Mice
Plaque vulnerability index was calculated:(positive percentage of macrophages + positive percentage of lipids)/(positive percentage of smooth muscle cells + positive percentage of collagen). In relation to the NC group, the plaque vulnerability index was raised in the miR-342-5p agomir group (P < 0.05) and decreased in the miR-342-5p antagomir group and the oe-Wnt3a group (both P < 0.05). Versus the miR-342-5p agomir group, the plaque vulnerability index was decreased in the miR-342-5p agomir + oe-Wnt3a group (P < 0.05) (Fig. 5E). Briefly, miR-342-5p targeted regulation of Wnt3a signaling pathway-mediated vulnerability of plaques in arterial tissues of AS mice.
Effects of Low Expression of miR-342-5p or Overexpression of Wnt3a on Angiogenesis in Aortic Plaque of ApoE −/− Mice
Antibodies against endothelial cell marker CD34 can detect blood vessel density [24]. By immunohistochemistry and Western blot. Versus the NC group, MVD was heightened in the miR-342-5p agomir group and attenuated in the miR-342-5p antagomir group and the oe-Wnt3a group (all P < 0.05). In comparison with the miR-342-5p agomir group, MVD was decreased in the miR-342-5p agomir + oe-Wnt3a group (P < 0.05) (Fig. 6A–C). Collectively, miR-342-5p targeting and regulating Wnt3a signaling pathway directly participated in regulating the density of neovascularization in plaques of AS mice.

Effects of low expression of miR-342-5p or overexpression of Wnt3a on MVD in aortic plaque of ApoE −/− мышей. A Immunohistochemical staining of CD34 in each group of ApoE −/− mice (scale bar:50 μm). B Comparison of MVD in aortic plaque in ApoE −/− мышей. C Comparison of CD34 protein expression in ApoE −/− мышей. n = 12. # P < 0.05 vs. the NC group. &P < 0.05 vs. the miR-342-5p agomir group. Measurement data were indicated as mean ± standard deviation. Comparisons among multiple groups were assessed by one-way ANOVA followed by Tukey’s multiple comparisons test. AS, atherosclerosis; NC, negative control
Discussion
AS is an unpredictable disease involving forms of chronic inflammation and vascular remodeling, and is the major cause of mortality and morbidity globally [25]. A previous study has discussed that miR-342-5p is implicated in regulating the progression of AS [26]. Also, it was mirrored that Wnt pathway is involved in facilitating the occurrence and development of diabetic AS [27]. As the related mechanisms of miR-342-5p and Wnt3a in AS remain to be excavated, our study was to inquire the effect of miR-342-5p targeted Wnt3a on formation of vulnerable plaque and angiogenesis of AS.
Our study revealed that highly expressed miR-342-5p and lowly expressed Wnt3a were found in aortic tissues of AS mice. A study has presented that macrophage-derived miR-342-5p is dramatically raised in early atherosclerotic lesions in ApoE −/− mice [13]. Another study has presented that miR-342-5p is markedly elevated in atrial fibrillation patients [28]. It has been reported that Wnt3a deficiency irreversibly injures hematopoietic stem cell self-renewal and results in defects in progenitor cell differentiation [29]. A study has shown that depletion of Wnt3a leads to defective cardiac function [30]. It has been revealed that Wnt3a expression in hippocampus of Alzheimer’s disease mice is remarkably decreased [31]. Another result from our study is that Wnt3a was directly targeted by miR-342-5p in AS mice. It has been reported that miR-342-5p can target the 3'-UTR of Wnt3a and negatively regulate its expression [14].
In addition, our study has suggested that TC, TG, LDL-C, IL-5, IL-12p70, IFN-γ, TNF-α and MDA contents were increased in serum, and HDL-C content and SOD activity were decreased. In addition, plaque area, lipid content, collagen content and MVD were enhanced as well as MOMA-2 expression was raised and α-SMA expression was decreased in AS mice. IFN-γ is a soluble cytokine with many functions, including anti-fibrosis, anti-proliferation, immunomodulation, apoptosis and anti-viral activities [32]. It has been revealed that glutamine treatment markedly raises SOD activity and reduces MDA content as well as increases Wnt3a protein levels in Alzheimer’s disease [31]. A study has revealed that the plasma levels of TC, TG and LDL-C are notably elevated and HDL-C is markedly reduced in AS [33]. A study has reported that TEMPOL supplementation, which has a value in suppressing metabolic disorders and raising atherosclerotic plaque stability, enhances plaque collagen content and reduces lipid content [34]. Zhou et al. noted that OPCRR treatments dramatically reduces the serum lipid profiles including TC, TG and LDL-C as well as and raises the HDL-C, also decreases MDA content as a product of lipid peroxidation and, moreover, declines serum levels of TNF-α in AS [35]. It has been presented that atherosclerotic samples have obviously reduced expression of α-SMA [36]. A study has presented that raised MVD is found in diseased aortas and especially in ruptured atherosclerotic plaque [37]. Furthermore, our study revealed that poor expression of miR-342-5p and overexpression of Wnt3a decreased he lipid levels, cytokine contents, oxidative stress response, plaque area and lipid content as well as increased collagen content, depleted MOMA-2 expression and restored α-SMA expression in aortic tissues in AS mice. It has been suggested previously that miR-342-5p is found to be positively linked to LDL-C and TNF-α serum levels and has an inverse correlation with HDL-C in coronary artery disease (CAD) patients [12]. Another study has verified that depletion of miR-342-5p inhibits AS [13]. Additionally, an experiment has presented that low serum level of Wnt1 is related to raised TG and LDL-C in premature CAD patient [38]. In addition, a study has showed that up-regulated Wnt3a, enhanced SOD content and decreased MDA content are found in the curcumin groups in Parkinson's disease rats [39].
Conclusion
In brief, our study for the first time discovered the mechanism of miR-342-5p/Wnt3 axis in AS and revealed that depleting miR-342-5p could reduce formation of vulnerable plaque and angiogenesis in AS mice via restoring Wnt3a, which may be a potential candidate for treatment of AS (Additional file 2:Fig. S2). miR-342-5p may have a synergistic effect with other miRNAs in atherosclerotic vascular disease, but due to time and funding constraints, we did not conduct further relevant discussions, which is also a limitation of this study.
Availability of data and materials
The original contributions presented in the study are included in the article/Supplementary Material, and further inquiries can be directed to the corresponding author.
Abbreviations
- miR-342-5p:
-
MicroRNA-342-5p
- A:
-
Atherosclerosis
- α-SMA:
-
α-Smooth muscle actin
- MVD:
-
Microvessel density
- miRNA:
-
MicroRNA
- oe:
-
Overexpression
- NC:
-
Negative control
- PBS:
-
Phosphate-buffered saline
- TC:
-
Total cholesterol
- TG:
-
Triglyceride
- LDL-C:
-
Low-density lipoprotein cholesterol
- HDL-C:
-
High-density lipoprotein cholesterol
- ELISA:
-
Enzyme-linked immunosorbent assay
- IL:
-
Interleukin
- TNF-α:
-
Tumor necrosis factor alpha
- IFN:
-
Interferon
- MDA:
-
Malondialdeyde
- SOD:
-
Superoxide dismutase
- RT-qPCR:
-
Reverse transcription quantitative polymerase chain reaction
Наноматериалы
- Характеристика отклонений частоты кристаллов кварца:допуск по частоте, стабильность частоты и старение
- Мониторинг состояния увеличивает и поддерживает эффективность производства бумаги
- Создание и визуализация Cyclocarbon
- Графен в динамиках и наушниках
- Понижающая регуляция микроРНК-342-5p или повышающая регуляция Wnt3a ингибирует ангиогенез и поддерживает стабиль…
- Экспериментальные исследования стабильности и естественной конвекции наножидкости TiO2-вода в корпусах с раз…
- Миниатюрные растягиваемые и высокоскоростные линейные суперконденсаторы
- Исследование первых принципов стабильности и STM-изображения борофена
- Потенциальная токсичность наночастиц диоксида титана для печени, мозга и эмбрионов у мышей
- Гидротермальный синтез наночастиц In2O3, гибридные двойники, гексагональные дисковые гетероструктуры ZnO для по…