


Роль пути апоптоза в цитотоксичности, вызванной свежими и выдержанными наночастицами оксида цинка
Аннотация
Наночастицы оксида цинка (ZnO NP) используются в широком спектре приложений, включая промышленность, коммерческие продукты и область медицины. Многочисленные механистические исследования токсичности НЧ ZnO были выполнены на нетронутых (свежих) НЧ. Однако цитотоксичность, вызванная трансформированными (старыми) НЧ ZnO, и лежащие в основе механизмы остаются неясными. Здесь мы наблюдали физико-химическую трансформацию НЧ ZnO, происходящую с течением времени, с последующей оценкой цитотоксичности свежих и старых НЧ. Мы обнаружили, что свежие НЧ ZnO вызывают более высокий уровень апоптоза, чем их старые аналоги. Соответственно, данные секвенирования РНК для стареющего гибрида человека и хомяка, обработанного ZnO NP ( A L ) клетки показали, что сигнальные пути p53, PI3k-Akt, FoXO, глутатиона, ErbB, HIF-1, окситоцина и Jak-STAT были обогащены, но не были пути апоптоза. Количественные результаты ПЦР выявили значительно более высокий уровень мРНК IL1B и CD69 в группах, получавших свежие NP, по сравнению с группами, получавшими старые ZnO NP и хлорид цинка. Приведенные выше результаты показали, что более низкая цитотоксичность старых ZnO NP частично объясняется их пониженной способностью вызывать апоптоз. Регуляция транскрипции множественных сигнальных путей, активируемых старыми НЧ, может помочь в построении клеточного гомеостаза. В совокупности наши результаты подчеркивают влияние процесса старения (трансформации окружающей среды) НЧ ZnO на их токсичность и биологические последствия.
Введение
С быстрым развитием нанотехнологий в последние десятилетия наночастицы (НЧ) нашли применение в различных областях, включая промышленность, повседневную жизнь человека и наномедицину [1, 2]. Инвентаризация потребительских товаров нанотехнологий (ИПЦ) показывает 30-кратное увеличение в период с 2005 по 2015 годы количества нанопродуктов, включая 762 товара для здоровья (фитнеса), 72 продукта питания (напитки) и 23 товара для детей [2]. Растущее применение НЧ в потребительских товарах и в различных областях увеличивает вероятность попадания НЧ в окружающую среду, что вызывает опасения с точки зрения безопасности в связи с их потенциальным неблагоприятным воздействием. НЧ оксида цинка (ZnO) являются одними из наиболее часто используемых НЧ, а его глобальный годовой объем производства достиг почти 3400 тонн [3, 4]. Некоторые вещества, которые ранее считались биологически инертными, могут стать токсичными в виде наночастиц. Все большее количество исследований подтверждают, что НЧ ZnO могут представлять значительный риск для клеток млекопитающих и животных, вызывая значительную токсичность [5,6,7].
Различные стратегии, включая покрытие, функционализацию поверхности и модификацию состояния окисления, были использованы для ослабления потенциальной токсичности НЧ путем изменения их физических и химических свойств (таких как растворение, агломерация и нарушение клеточных мембран) [8,9,10, 11]. Хотя эти модификации НЧ в определенной степени ослабляют их токсическое действие, использование НЧ не всегда безопасно, особенно при определенных условиях воздействия и средах [12,13,14]. На самом деле, многие виды НЧ нестабильны и склонны к «старению» или «трансформации окружающей среды» после преднамеренного или непреднамеренного выброса в естественную среду [14,15,16,17]. В последние годы была проделана большая работа по изучению процесса экологической трансформации АЭС; однако исследования токсических эффектов «трансформированных (старых)» НЧ все еще очень ограничены, не говоря уже об их токсических механизмах.
Как типичный представитель нестойких НЧ, НЧ ZnO обладают очень высокой реакционной способностью и склонны к изменению физических и химических свойств и состояния присутствия после попадания в окружающую среду или проглатывания животными, что может значительно повлиять на их токсикологические эффекты [17 , 18]. Например, исследования показали, что процесс сульфидирования НЧ ZnO изменяет заряд, гидрофобность и состояние агрегации, что приводит к адсорбции НЧ в сульфидном состоянии в человеческой слюне, потом и жидкости бронхоальвеолярного лаважа. Кроме того, белок, адсорбированный НЧ ZnO, образует особую белковую корону, которая обычно влияет на его биологический эффект [19]. Фосфаты в физиологических растворах могут превращать НЧ ZnO в метастабильный ZnHPO 4 и Zn 3 (ЗП 4 ) 2 в течение 5–10 ч [20]. Процесс полной трансформации НЧ ZnO (≤ 3 мкг / мл) в системе воздействия in vitro на Т-лимфоциты человека (37 ° C, среда для культивирования клеток RPMI1640, содержащая 10% FBS в течение 24 ч) исследовали с использованием синхротронного излучения X- спектроскопия поглощения лучей вблизи края структуры (XANES) [21]. Вышеупомянутые исследования предполагают недооценку рисков для окружающей среды и здоровья НЧ ZnO исключительно путем оценки биологических эффектов чистых (свежих) НЧ ZnO. В свете этой проблемы существует острая необходимость всестороннего понимания процессов старения и экологической трансформации ЯЧ [22].
Наше предыдущее исследование показало, что НЧ ZnO, выдержанные в течение 40–120 дней в сверхчистой воде, претерпевают физико-химические превращения и превращаются в Zn 5 (CO 3 ) 2 (ОН) 6 , Zn (OH) 2 , и Zn 2+ [23]. Интересно, что старые НЧ ZnO проявляли более низкую цитотоксичность, чем свежие аналоги [23], но механизмы токсичности такого рода вариаций неясны. В настоящем исследовании мы намеревались изучить основные причины различной цитотоксичности между свежими и старыми НЧ ZnO. Систематически применялись НЧ ZnO с двумя различными размерами частиц (20 нм и 90–200 нм). Анализы цитотоксичности показали, что старые НЧ ZnO вызывают менее выраженные морфологические аномалии и относительно более высокую жизнеспособность клеток, чем их свежие аналоги. Данные секвенирования РНК показали, что апоптотические гены были обогащены в свежих клетках, обработанных ZnO NP, тогда как эти гены гораздо меньше подвергались воздействию старых обработок ZnO NP. Кроме того, клетки, подвергшиеся воздействию старых НЧ ZnO, показали пониженный уровень расщепленного белка каспазы-3, что дополнительно указывает на более высокую эффективность свежих НЧ ZnO в индукции апоптоза в культивируемых клетках. В сочетании с нашими предыдущими выводами это исследование показало, что снижение цитотоксичности старых ZnO NPs объясняется их ослабленной способностью запускать апоптоз клеток.
Материалы и методы
Наночастицы и реагенты
Коммерчески доступные нанопорошки ZnO (НЧ ZnO), средний размер которых, по данным производителя, составляет 20 нм (чистота 99,5%, почти сферическая) и 90–200 нм (чистота 99,9%, неправильная морфология), были приобретены в компании Nanostructured &Amorphous Materials (Хьюстон, Техас). ). Если не указано иное, все реагенты и химические вещества, использованные в этом исследовании, были приобретены у Sigma-Aldrich (Шанхай, Китай).
Дисперсия, старение и характеристика наночастиц
Исходные суспензии НЧ ZnO (1 мг / мл) были приготовлены суспендированием сухих нанопорошков в Milli- Q H 2 O (Millipore, 18 МОм см) и стерилизовали автоклавированием (120 ° C, 30 мин), а затем хранили при 25 ° C в течение периода естественного старения от 0 до 60 дней. Естественно трансформированные НЧ ZnO в течение 0 и 60 дней были обозначены как свежие и старые НЧ соответственно. Для обеспечения надлежащего диспергирования свежие и выдержанные суспензии обрабатывали ультразвуком (100 Вт) в течение 30 минут в ультразвуковой ванне перед характеризацией или инкубацией с клетками. Морфология, размер частиц и агрегация свежих и состаренных НЧ ZnO были охарактеризованы с помощью просвечивающей электронной микроскопии (ТЕМ, JEOL JEM-2010, Токио, Япония). Кристаллическую структуру свежих и состаренных наночастиц ZnO определяли с помощью дифракции рентгеновских лучей (XRD, PANalytical B.V., Шанхай, Китай) путем сравнения с аутентичными стандартами. Детали процесса естественного старения и характеристики наночастиц ZnO были описаны ранее [23].
Культура клеток и обработка НЧ ZnO
А L линия клеток, своего рода гибридные клетки человека и хомяка, образованные путем слияния gly2A В этом исследовании использовали мутант яичника китайского хомячка (СНО) и человеческие фибробласты. Эти гибридные клетки содержали стандартный набор хромосом CHO-K1 и одну копию хромосомы человека 11 и были культивированы в среде Ham's F12 (Гиклон, Гранд-Айленд, штат Нью-Йорк) с добавлением фетальной бычьей сыворотки (8%, Гиклон, Гранд-Айленд, штат Нью-Йорк). ), гентамицин (25 мкг / мл) и глицин (2 × 10 –4 M) при 37 ° C в увлажненном 5% CO 2 инкубатор [24]. Исходные суспензии свежих и состаренных НЧ ZnO диспергировали в течение 30 минут обработки ультразвуком (100 Вт) для предотвращения агломерации, затем разбавляли до соответствующих концентраций средой для культивирования клеток для экспонирования клеток. Клетки, содержащиеся в среде для культивирования клеток без НЧ, служили контролем в каждом эксперименте.
Анализ для определения цитотоксичности
А L клетки на логарифмической фазе роста культивировали на предметных стеклах в 35-мм чашках Петри (6 × 10 4 клеток / чашку) в течение 24 часов до стимуляции с последующей обработкой 2 мл культуральной среды, содержащей 1, 5, 10, 12, 15 и 20 мкг / мл свежих или старых НЧ ZnO 72 часа. По истечении времени обработки изображения морфологии клеток получали с помощью микроскопа Leica DM4B (Leica, Германия). ZnCl 2 был включен в качестве эталона ионов цинка для сравнения цитотоксичности с НЧ ZnO.
Набор для подсчета клеток (CCK-8) (APExBIO, Шанхай, Китай) использовали для определения жизнеспособности клеток. В деталях, A L клетки высевали в 96-луночные планшеты (4 × 10 3 клеток / лунку) средой для культивирования клеток в течение 24 часов и обработанной средой, содержащей различные концентрации ZnCl 2 , свежие и состаренные НЧ ZnO в течение 24, 48 и 72 ч соответственно. Для рабочего раствора объем добавленных НЧ из исходной суспензии составлял менее 5% от общего объема питательной среды в каждой лунке. После завершения времени обработки культуральную среду аспирировали, и клетки инкубировали со 100 мкл рабочего раствора CCK-8 в течение 2 часов при 37 ° C, следуя инструкциям производителя. Затем оптическую плотность регистрировали при 450 нм с использованием флуоресцентного ридера Spectra Max M2 (Molecular Devices, Wokingham, Berks, UK). Жизнеспособность клеток рассчитывалась как процент поглощения в лунках, при этом каждая концентрация НЧ нормировалась на поглощение контрольных клеток (100%).
Извлечение РНК, обратная транскрипция и количественная ПЦР
А L клетки на логарифмической фазе роста высевали в чашку Петри диаметром 35 мм (6 × 10 4 клеток / чашку) средой для культивирования клеток в течение 24 часов. Затем среду заменяли 2 мл культуральной среды, содержащей 12 мкг / мл ZnCl 2 . , свежие и выдержанные НЧ ZnO в течение 72 ч. По истечении времени обработки культуральную среду аспирировали и клетки промывали 3 раза PBS. Затем в каждую чашку добавляли 1 мл реагента Trizol (Invitrogen, Carlsbad, CA, USA) для экстракции общей РНК в соответствии с инструкциями производителя. Концентрацию и чистоту общей РНК, полученной после экстракции, количественно определяли с помощью спектрофотометра Q5000UV-Vis (Quawell, США). После количественной оценки проводили обратную транскрипцию с использованием набора TransGene RT-PCR (TransGene Biotech, Пекин, Китай) для получения кДНК из матрицы РНК в соответствии с протоколами производителя. Полученные образцы кДНК были количественно определены с помощью спектрофотометра Q5000 UV-Vis и затем проанализированы с использованием SYBR-Green в качестве флуоресцентного красителя (TransGene Biotech, Пекин, Китай) в системе Roche RT-PCR (Applied Biosystems) [25].
Ген домашнего хозяйства, кодирующий глицеральдегид-3-фосфатдегидрогеназу ( Gapdh ) использовался в качестве внутреннего контроля для оценки Il-1α , Ил-1β , Caspase 3 , CD69 , июн и MT1 Экспрессия мРНК. Результаты были выражены как относительное соотношение экспрессии между целевым геном и Gapdh . . Последовательности праймеров, использованные в этом исследовании, представлены в таблице 1.
Анализ данных секвенирования РНК
Общие образцы РНК A L клетки из контрольной группы, возрастная группа, обработанная ZnO NP, и ZnCl 2 группы, получавшие лечение, были секвенированы BangFei Bioscience (Пекин, Китай). Вкратце, общая РНК A L клетки экстрагировали, следуя протоколам ТРИЗОЛ, до изопропонального осаждения. Затем образцы РНК ресуспендировали в буфере для экстракции перед секвенированием. Необработанные данные секвенирования РНК анализировали с использованием пакета R Deseq2 [Eric1]. Диаграмма Венна была сгенерирована R-пакетом VennDiagram [Eric1.2]. Значительно измененные гены использовали для дальнейшего анализа обогащения путей. Эксперименты проводились в трех независимых повторностях. Гены рРНК, митохондриальные гены и гены размером менее 40 п.н. были исключены из анализа.
Данные секвенирования РНК, эталонные серии GSE97852, GSE60159 и GSE39444, были получены из Gene Expression Omnibus [Eric 2, 3, 4]. График анализа обогащения генетического набора был создан R (версия 3.6.2) с использованием пакета fgsea [Eric 5]. Гены апоптоза с 1,5-кратным значительным изменением & p значение <0,05 использовалось для дальнейшего анализа. Тепловая карта с деревом генов была создана R-пакетом «ComplexHeatmap» [Эрик 6]. Среднее сцепление использовалось в качестве метода кластеризации, а Евклидова использовалась в качестве метода измерения расстояния. Анализ обогащения путей проводился с использованием STRING2.0 [Eric 7].
Вестерн-блоттинг
А L клетки на логарифмической фазе роста высевали в чашку Петри диаметром 60 мм (1,5 × 10 5 клеток / чашку) средой для культивирования клеток в течение 24 часов. Затем среду заменяли 4 мл культуральной среды, содержащей 12 мкг / мл свежих или выдержанных НЧ ZnO, в течение 24 ч. В конце периода воздействия культуральную среду аспирировали, а затем клетки промывали 3 раза PBS и лизировали на льду лизисным буфером RIPA (Beyotime, Китай) для сбора клеточных белков. Равные количества клеточных белков разделяли на 12% SDS-PAGE гелях и затем переносили на мембрану из поливинилиденфторида (PVDF) (Roche, Swiss). Вкратце, после 2-часового блокирования 5% обезжиренным молоком в TBST при 25 ° C мембраны затем инкубировали с первичным антителом в соответствующих разведениях (согласно протоколам производителя) при 4 ° C в течение ночи с последующей инкубацией с конъюгированным с HRP вторичным антителом. антитела (1:5000, Promega, Мэдисон, США) в течение 2 ч при 25 ° C. Наконец, иммунную метку выявляли с помощью раствора повышенной хемилюминесценции (ECL) (BOSTER, Китай). Первичные антитела против про / расщепленной каспазы-3 и против актина были приобретены у Cell Signaling Technology и ImmunoWay соответственно.
Статистика
Статистический анализ проводился на основе результатов, полученных как минимум в трех независимых экспериментах. Все данные были представлены в виде средних значений ± стандартное отклонение (SD) и статистически сравнивались с использованием одностороннего дисперсионного анализа (ANOVA). На всех участках p значения <0,05 были отмечены * и считались статистически значимыми.
Результаты
Характеристика НЧ ZnO
Чтобы определить различия в подробных физико-химических характеристиках между свежими и состаренными НЧ ZnO, мы сначала наблюдали морфологию НЧ с помощью ПЭМ (рис. 1А). Наши результаты показали, что свежие НЧ ZnO размером 20 нм были кристаллами, близкими к сферической, а свежие НЧ ZnO размером 90–200 нм были нерегулярно стержневидными / кубическими кристаллами. Размер отдельных частиц соответствовал размеру, предоставленному производителем. Очевидно, НЧ ZnO размером 20 нм и 90–200 нм были склонны к агрегации в сверхчистой воде. Кроме того, независимо от формы и размера исходных наночастиц, микроструктура наночастиц ZnO размером 20 нм и 90–200 нм резко изменилась с прозрачной кристаллической структуры на аморфное или пластинчатое / игольчатое состояние после выдержки в течение 60 дней. Кроме того, кристаллическая природа и фазовая чистота как свежих, так и состаренных наночастиц были определены с помощью дифракции рентгеновских лучей (XRD) с Cu Kα-излучением ( λ =0,15418 нм) приближаются к температуре 25 ° C, как показано на рис. 1B. Рентгенограмма свежих НЧ ZnO показала, что образцы состояли из кристаллической структуры вюрцита и не было идентифицировано характерных пиков примесей, что свидетельствует о высоком качестве свежих НЧ. Для старых НЧ рентгенограмма показала новообразование Zn 5 (CO 3 ) 2 (ОН) 6 (номер карты 00-011-0287) и Zn (OH) 2 (номер карты 00-003-0888) твердые фазы, свидетельствующие о химическом превращении НЧ ZnO (20 и 90–200 нм) в процессе старения.
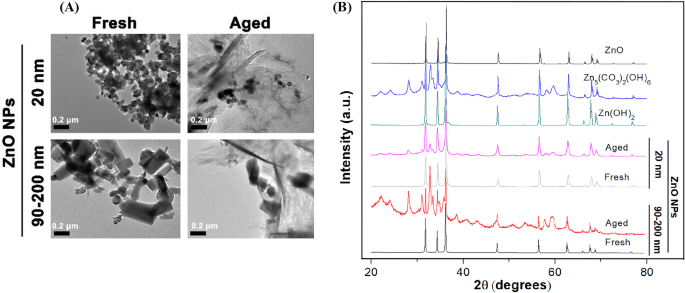
Физико-химические характеристики свежих и выдержанных НЧ ZnO. А Репрезентативные микрофотографии свежих и старых НЧ (100 мкг / мл, 20 и 90–200 нм) в Milli- Q вода с использованием ПЭМ низкого разрешения, B Рентгенограммы свежих НЧ, состаренных НЧ, ZnO, Zn (OH) 2 и Zn 5 (CO 3 ) 2 (ОН) 6 ссылки в сушеном виде
Морфологическое наблюдение A L Клетки, подвергшиеся действию свежих и старых НЧ ZnO
Обработка НЧ приводит к заметному изменению формы или морфологии клеток in vitro [26]. Следовательно, A L клетки, подвергнутые воздействию свежих или старых ZnO NPs в концентрации 10 и 15 мкг / мл в течение 72 часов, исследовали под стереоскопическим микроскопом. Как показано на рис. 2, морфология клеток в контрольной группе оставалась нормальной. Клетки прилипали хорошо, большая часть прикреплялась в течение 2 часов. Большинство клеток были веретенообразными или многоугольными, при этом несколько недавно делящихся клеток показали более прозрачную цитоплазму и лучшую дисперсию в процессе прикрепления. Обработка 12 мкг / мл свежих НЧ ZnO (20 нм и 90–200 нм) в течение 72 часов значительно изменила морфологию клеток. Хотя большинство клеток прикрепилось в течение 3-5 часов, они не могли хорошо распространяться, а некоторые клетки стали округлыми и потеряли многоугольную форму. Когда концентрация ZnO NP была увеличена до 15 мкг / мл, обработанные клетки атрофировались и не могли прилипать, что свидетельствует о значительно более низкой жизнеспособности их клеток, чем у клеток, обработанных 10 мкг / мл. Эти результаты показали, что LC100 для свежих НЧ ZnO, вероятно, составляет менее 15 мкг / мл при 72-часовой обработке. Напротив, морфология клеток в группах с возрастом 20 нм и 90–200 нм, обработанных NP (15 мкг / мл), существенно не пострадала, и большинство выживших клеток могло прилипать и распространяться, при этом наблюдались менее полумертвые клетки, что свидетельствует о том, что старые НЧ ZnO гораздо менее цитотоксичны, чем свежие НЧ ZnO.
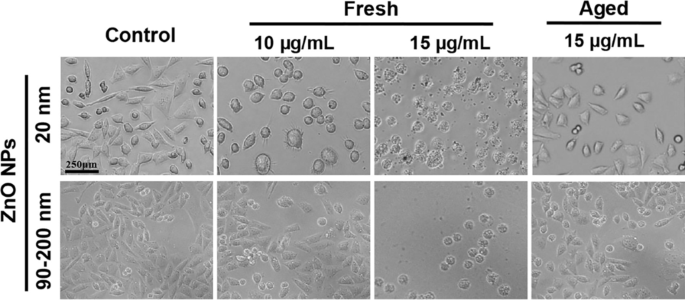
Изменения морфологии в A L клетки после воздействия свежих или старых ZnO NP в течение 72 часов в дополнительной среде Ham’s F12, а неэкспонированные клетки использовали в качестве контрольных групп. А L морфологию клеток наблюдали с помощью оптического микроскопа при 10-кратном увеличении
Старые НЧ ZnO вызывают более низкую цитотоксичность, чем свежие НЧ
Для дальнейшего изучения разницы в цитотоксичности свежих и старых ZnO NP, мы исследовали жизнеспособность клеток с помощью наборов CCK-8. Как показано на рис. 3, инкубация A L клетки с градиентными дозами свежих и старых НЧ ZnO (от 0 до 20 мкг / мл, 20 нм и 90–200 нм) в течение 24 ч, 48 ч или 72 ч показали дозозависимое снижение жизнеспособности клеток. Никакой очевидной цитотоксичности не наблюдалось при обработке клеток НЧ ZnO ≤ 10 мкг / мл. Когда дозировка свежих и состаренных НЧ ZnO повышалась до 12 и 15 мкг / мл, жизнеспособность клеток проявляла тенденцию к уменьшению в зависимости от времени. Очевидно, жизнеспособность клеток в старых группах, обработанных NP, была значительно выше, чем в группах, обработанных свежими NP. Кроме того, ZnCl 2 -обработка также снижает жизнеспособность клеток в зависимости от дозы и времени, тогда как цитотоксичность ZnCl 2 было намного меньше, чем у свежих и старых НЧ ZnO.
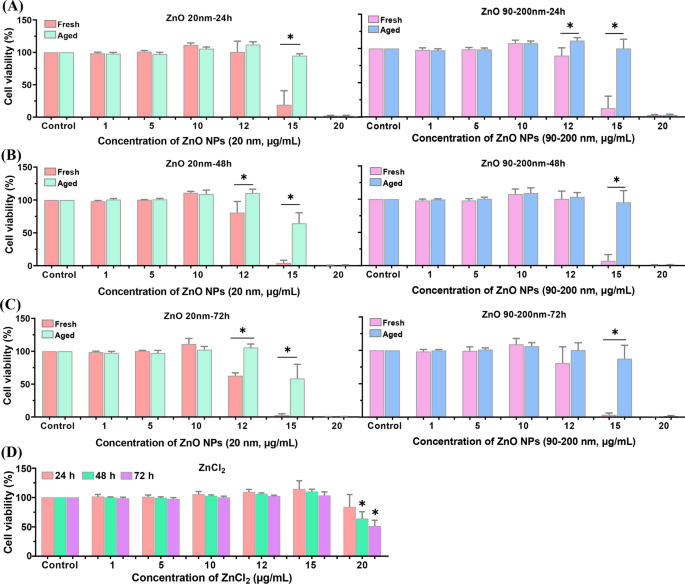
Жизнеспособность клеток, индуцированная свежими и старыми НЧ ZnO в A L клетки. А L клетки инкубировали с различными концентрациями свежих и старых НЧ ZnO (20 и 90–200 нм) в течение 24 часов ( A ), 48 ч ( B ) и 72 часа ( C ). D А L клетки подвергались воздействию различных концентраций ZnCl 2 на разное время. Данные были основаны на ≥ 3 независимых экспериментах и выражены как среднее ± стандартное отклонение * p <0,05
Обработка свежими НЧ ZnO активировала пути апоптоза и повысила экспрессию апоптотических генов
Чтобы раскрыть основные механизмы, приводящие к более низкой цитотоксичности старых НЧ, мы проанализировали данные секвенирования РНК как для свежих, так и для старых НЧ ZnO. Как показано на фиг. 4A, B, после обработки свежими НЧ ZnO в клетках Jurkat активировался путь апоптоза ( p =0,017) и ячейки HMDM ( p =0,041). Гены апоптоза: ANXA1 , CYLD , TNFSF10 , IER3 , CDKN1A , ИЮН , SAT1 , PMAIP1 , CD38 и ISG20 были значительно обогащены свежими обработанными ZnO NP клетками Jurkat. Гены апоптоза: CD38 , TNFRSF12A , CCNA1 , BMP2 , PPP2R5B , EREG , IFNGR1 , CD44 , CD14 , GNA15 , GCH1 , TIMP1 , BTG2 , IL1B , IL1A , BTG3 , BCL2L11 , SC5D и SPTAN1 были значительно обогащены свежими обработанными ZnO NP клетками HMDM (фиг. 4C, D). Поскольку клетки Джуркета (Т-лимфоциты периферической крови) и клетки HMDM (макрофаги, происходящие из моноцитов человека) являются разными типами клеток, то их способ запуска апоптоза может быть разным. В целом, эти результаты показали, что воздействие свежих НЧ ZnO может активировать различные пути апоптоза в различных типах клеток.
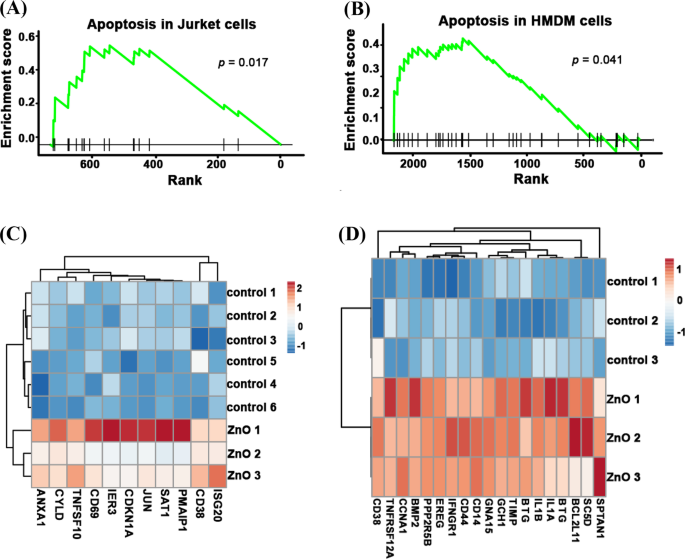
Путь апоптоза был обогащен данными РНК-seq свежих обработанных ZnO NP клеток Jurket и HMDM. Оценка обогащения значительно экспрессируемых генов пути апоптоза свежих обработанных ZnO NP клеток Jurket ( A ) и ячейки HMDM ( B ). Тепловая карта экспрессии апоптотического гена свежих обработанных ZnO NP клеток Jurket ( C ) и ячейки HMDM ( D ) и их контрольные группы
Старые НЧ ZnO не регулируют экспрессию апоптотических генов в виде свежих НЧ ZnO
Наши данные о секвенировании РНК для стареющих ZnO NP, обработанных A L клетки показали, что p53, PI3k-Akt, FoXO, глутатион, ErbB, HIF-1, окситоцин и сигнальный путь Jak-STAT были обогащены (рис. 5A). Гены апоптоза, обогащенные клетками Jurket и HMDM, не подвергались значительному влиянию в старых клетках, обработанных ZnO NP (фиг. 5B). Для дальнейшего подтверждения результатов мы проверили экспрессию родственных генов с помощью ПЦР в реальном времени. Мы обнаружили, что некоторые из генов апоптоза: BMP2 , PMAIP1 , IL1α , CD69 , CCNA1 , CD38 и IL1β не обнаруживались в старых обработанных НЧ ZnO A L клетки (данные не показаны), вероятно, потому, что большинство этих генов экспрессируются в клетках иммунной системы. Другие активируемые гены апоптоза ( IL1α , IL1β и CD59 ), наблюдаемые в группах, обработанных свежими ZnO NP, существенно не изменились в уровнях экспрессии при обработке старых ZnO NP. В то время как MT1 , которые служат в качестве положительного контроля, были значительно увеличены в уровне экспрессии, экспрессия каспазы 3 существенно не изменилось (рис. 5C). Эти данные свидетельствуют о том, что старые НЧ ZnO, в отличие от своих свежих аналогов, менее эффективны в активации генов пути апоптоза в A L ячеек.
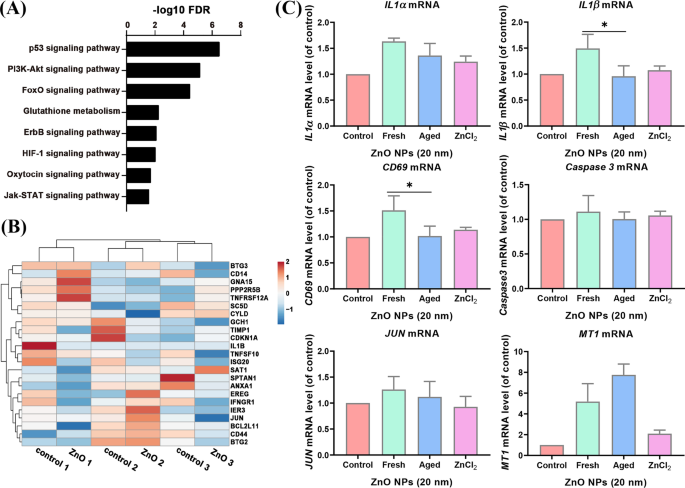
Путь апоптоза не был обогащен данными РНК-seq для стареющих ZnO NP-обработанных A L клетки. ( А ) Анализ онтологии генов обогащенных путей из старых ZnO NP-обработанных A L клетки. ( B Тепловая карта экспрессии апоптотических генов у пожилых ZnO NP-обработанных A L ячейки и контрольная группа. ( C ) Экспрессия выбранных апоптотических генов и контрольных генов ( MT1 ) в свежем и выдержанном ZnO, обработанном NP A L ячейки
Свежие, но не выдержанные НЧ ZnO повышают уровень экспрессии активированного белка каспазы 3
Обнаружение экспрессии гена каспазы 3 сам по себе не может прямо указывать на активацию пути апоптоза. Для дальнейшего анализа того, может ли обработка НЧ ZnO изменить уровень апоптотических белков, с помощью Вестерн-блоттинга исследовали экспрессию расщепленного белка каспазы 3, обычно используемого биомаркера для индикации активации апоптоза клеток [27]. Как показано на рис. 6, по сравнению с контрольной группой, обработка свежими НЧ ZnO (20 нм) увеличивала клеточный уровень расщепленного белка каспазы 3 в 1,31 ± 0,023 раза, что было значительно выше, чем у старых 20 нм НЧ ZnO- обработанная группа (1,12 ± 0,039 раза). Когда размер частиц свежих НЧ ZnO был увеличен до 90–200 нм, экспрессия расщепленного белка каспазы 3, индуцированная свежими НЧ, увеличивалась в 1,46 ± 0,078 раза, что значительно выше, чем у старых НЧ (в 1,07 ± 0,075 раза). . Эти данные дополнительно иллюстрируют более высокую эффективность свежих НЧ ZnO в индукции апоптоза клеток по сравнению с их старыми аналогами.
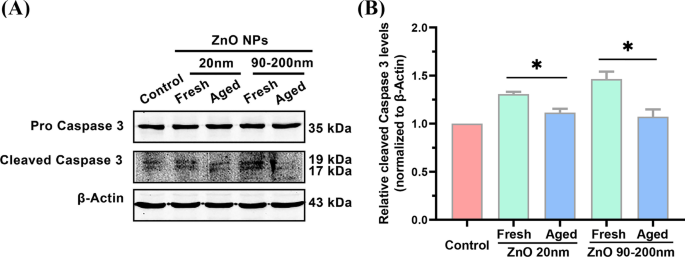
Уровни апоптоза в A L клетки инкубировали со свежими и выдержанными НЧ ZnO (20 и 90–200 нм). Вестерн-блоттинг ( A ) и количественной оценки ( B ) уровней расщепленного белка каспазы 3 при инкубации клеток с 12 мкг / мл свежих и старых НЧ ZnO (20 и 90–200 нм) в течение 72 часов. Данные были основаны на ≥ 3 независимых экспериментах и выражены как среднее ± стандартное отклонение * p <0,05
Обсуждение
Сообщалось, что НЧ ZnO претерпевают физико-химические превращения в Zn 5 (CO 3 ) 2 (ОН) 6 с выпуском Zn 2+ в процессе естественного старения [23, 28]. Однако цитотоксичность, вызванная трансформированными (старыми) НЧ ZnO, и лежащие в основе механизмы остаются неясными. Здесь, чтобы раскрыть механизм различной цитотоксичности между свежими и старыми НЧ ZnO, был проведен анализ секвенирования РНК и тест ОТ-ПЦР. Также был применен вестерн-блоттинг для изучения уровня белка каспазы 3, ключевого исполнителя апоптоза клеток.
Наши данные показали, что старые НЧ ZnO вызывают гораздо меньшую цитотоксичность, чем свежие НЧ ZnO в A L клетки. LC 100 обоих свежих НЧ ZnO (90-200 нм и 20 нм) в нашем настоящем исследовании было ниже 15 мкг / мл (рис. 3), что согласуется с предыдущими выводами о том, что LC 100 НЧ ZnO с длиной волны 19–36 нм до клеток NIH-3T3 или MSTO составляет около 15 мкг / мл [29]. Мы подтвердили, что экологические преобразования физико-химических свойств НЧ могут резко изменить их токсичность. Сообщалось, что процесс сульфидирования НЧ ZnO изменяет их заряд, гидрофобность и состояние агрегации, что приводит к адсорбции НЧ в сульфидном состоянии в человеческой слюне, потом и жидкости бронхоальвеолярного лаважа. При этом белок, адсорбированный НЧ ZnO, образовывал особую белковую корону, что сказывалось на его биологическом действии [19]. Фосфаты, широко присутствующие в физиологических растворах (таких как слюна), могут превращать НЧ ZnO в метастабильный ZnHPO 4 и Zn 3 (ЗП 4 ) 2 в течение 5–10 ч и проявил цитотоксичность по отношению к эпителиальным клеткам пищеварительного тракта [20]. Иваск и др. доказали наличие полной трансформации НЧ ZnO (≤ 3 мкг / мл) в системе воздействия in vitro на Т-лимфоциты человека (37 ° C, среда для культивирования клеток RPMI1640, содержащая 10% FBS, в течение 24 ч) с использованием синхротронного излучения, поглощающего рентгеновские лучи. спектроскопия ближней структуры (XANES). Спектр и цитотоксичность продуктов трансформации соответствовали таковым ZnSO 4 [21]. Наши результаты показали дозозависимую и зависящую от времени токсичность ZnCl 2 в A L клетки, цитотоксичность которых намного ниже, чем у свежих и старых НЧ ZnO (рис. 3). Это наблюдение дополнительно объясняет открытие, что цитотоксичность свежего ZnO NP не полностью объясняется высвобожденным Zn 2+ [30].
Наше предыдущее исследование также показало, что старые НЧ ZnO обладают более высокой эффективностью в отношении выработки АФК (активных форм кислорода), а также меньшей способностью убивать клетки по сравнению со свежими НЧ ZnO [23]. Мы полагаем, что более низкая цитотоксичность, вызванная старыми НЧ ZnO, может быть более переносимой в клетках млекопитающих. Настоящее исследование данных секвенирования РНК показало, что апоптотические гены были активированы в свежих клетках, обработанных ZnO NP, тогда как в старых группах, обработанных NP, на них было гораздо меньше воздействовать. IL1α и IL1β являются членами семейства цитокинов интерлейкина 1. Высвобождение IL1α и IL1β активирует частично зависимый апоптоз от каспазы 8 [31]. CD69 кодирует член суперсемейства кальций-зависимых лектинов трансмембранных рецепторов типа II. Повышенная экспрессия CD69 была связана с повышенной экспрессией апоптоза аннексина V и маркера CD95 (Fas) [32]. JUN представляет собой субъединицу фактора транскрипции AP-1. Повышенная активность JUN протеолитически расщепляет альфа-фодрин, субстрат интерлейкин-1-бета-превращающего фермента (ICE), и семейство цистеиновых протеаз CED-3, что в дальнейшем вызывает запрограммированную гибель клеток [33]. Повышенная экспрессия этих апоптотических генов показала, что свежие НЧ запускают апоптоз несколькими различными способами. After the elevation of these apoptotic gene expressions, apoptosis processes are eventually executed by apoptotic proteins (Fig. 7). Caspase 3 is the core protease for various apoptotic scenarios; cleavage of this protein is necessary to activate both extrinsic and intrinsic apoptotic pathways [34, 35]. Therefore, detection of cleaved caspase 3 is a common method for identifying apoptosis induced by a wide variety of apoptotic signals [36]. Our Western blotting data revealed that, for both 20 nm and 90–200 nm ZnO NPs, sublethal exposure did not alter the level of Pro caspase 3 in all treatment groups. In contrast, cleaved Caspase 3 was significantly elevated by fresh NPs treatment, where aged NPs showed few (if any) effects on the level of cleaved caspase 3 (Fig. 6). Combined with RNA expression analysis, our results clearly elucidated the higher potency of fresh ZnO NPs in inducing cell apoptosis.

Model for Fresh ZnO NPs but not aged ZnO NPs induces Caspase 8- and Caspase 3-dependent apoptosis. The increased expression of apoptotic gene CD69 activates Fas and apoptosis annexin V expression in fresh ZnO NP-exposed mammalian cells. The increased expression of apoptotic gene IL1α and IL1β partially activates Caspase 8-dependent apoptosis. It further causes activation of Caspase 3 and induces apoptosis. All these changes in mRNA and protein level were not detectable in aged ZnO NPs-exposed mammalian cells
Conclusions
In the present study, the natural physicochemical transformation of ZnO NPs in ultrapure water was confirmed, and variations in cytotoxicity induced by fresh &aged NPs were investigated. We focused on RNA sequencing data from our aged ZnO NP-treated A L cells and that of fresh NPs from database. We compared those signaling pathway specifically enriched in aged NP-treated group, which are different from that of fresh NP- or ZnCl2 -treated groups. Our data indicated that the lower cytotoxicity of aged ZnO NPs is closely related to its attenuated ability in inducing apoptosis, while the transcriptional regulation of the multiple pathways activated by NPs promotes the establishment of cellular homeostasis in mammalian cells.
Доступность данных и материалов
Не применимо.
Сокращения
- НП:
-
Nanopowders
- ZnO:
-
Оксид цинка
- Zn5 (CO3 ) 2 (ОН) 6 :
-
Hydrozincite
- Zn (OH)2 :
-
Zinc hydroxide
- ZnCl2 :
-
Zinc chloride
- ZnSO4 :
-
Сульфид цинка
- ZnHPO4 :
-
Zinc hydrogen phosphate
- Zn3 (ЗП 4 ) 2 :
-
Zinc phosphate
- A L cells:
-
Human–hamster hybrid cells
- CHO cells:
-
Chinese hamster ovary cells
- Jurket cells:
-
Peripheral blood T lymphocyte cells
- HMDM cells:
-
Human monocyte-derived macrophages
- NIH-3T3cells:
-
Mouse embryonic cells
- MSTO cells:
-
Human lung cancer cells
- RPMI1640:
-
Roswell Park Memorial Institute 1640
- ICE:
-
Interleukin 1beta-converting enzyme
- CED-3:
-
Caenorhabditis elegans death gene
- IL1α:
-
Interleukin 1alpha
- IL1β:
-
Interleukin 1beta
- mRNA:
-
Messenger ribonucleic acid
- cDNA:
-
Complementary deoxyribonucleic acid
- FBS:
-
Фетальная бычья сыворотка
- ТЕМ:
-
Просвечивающая электронная микроскопия
- XRD:
-
Рентгеновская дифракция
- RT-PCR:
-
Real-time polymerase chain reaction
- CPI:
-
The Nanotechnology Consumer Product Inventory
- XANES:
-
Synchrotron radiation X-ray absorption near-edge structure spectroscopy
- RIPA:
-
Radio immunoprecipitation assay
- SDS-PAGE:
-
Polyacrylamide gel electrophoresis
- PVDF:
-
Поливинилиденфторид
- ECL:
-
Enhanced chemiluminescence
- CCK-8:
-
Набор для подсчета клеток-8
Наноматериалы
- Биосовместимые наночастицы FePO4:доставка лекарств, стабилизация РНК и функциональная активность
- Влияние толщины бислоя на морфологические, оптические и электрические свойства наноламинатов Al2O3 / ZnO
- Настройка морфологии поверхности и свойств пленок ZnO путем создания межфазного слоя
- Зеленая способность к синтезу и стабилизации наночастиц меди:каталитическая, антибактериальная, цитотоксич…
- Изготовление и характеристика ZnO Nano-Clips с помощью процесса, опосредованного полиолом
- Сравнительное исследование биосинтезированных наночастиц оксида меди и цинка in vivo путем внутрибрюшинного и…
- Зеленый синтез металлов и наночастиц оксидов металлов и их влияние на одноклеточную водоросль Chlamydomonas reinhardtii
- Свойства наночастиц оксида цинка и их активность против микробов
- Простой подход к получению наноразмерного оксида цинка в воде / глицерине с чрезвычайно концентрированными и…
- Роль робототехники и автоматизации в Индустрии 4.0