


F-actin регулирует остеобластную дифференцировку мезенхимальных стволовых клеток на нанотрубках TiO2 посредством MKL1 и YAP / TAZ
Аннотация
Титан и титановые сплавы широко используются в ортопедических имплантатах. Изменение нанотопографии обеспечивает новую стратегию улучшения остеоинтеграции титановых субстратов. Полимеризация нитчатого актина (F-актина), как структура механической нагрузки, обычно считается вовлеченной в миграцию клеток, эндоцитоз, деление клеток и поддержание формы клеток. Вовлечен ли F-actin и как он действует в индуцированной нанотрубками остеогенной дифференцировке мезенхимальных стволовых клеток (МСК), еще предстоит выяснить. В этом исследовании мы изготовили TiO 2 нанотрубки на поверхности титановой подложки путем анодного окисления и охарактеризовали их особенности с помощью сканирующей электронной микроскопии (SEM), рентгеновского энергодисперсионного анализа (EDS) и атомно-силовой микроскопии (AFM). Окрашивание щелочной фосфатазой (ALP), вестерн-блоттинг, qRT-PCR и иммунофлуоресцентное окрашивание проводили для изучения остеогенного потенциала, уровня F-актина и экспрессии MKL1 и YAP / TAZ. Наши результаты показали, что внутренний диаметр и шероховатость TiO 2 нанотрубки увеличивались с увеличением напряжения анодного окисления от 30 до 70 В, в то время как их высота постоянно составляла 2 мкм. Кроме того, чем больше диаметр трубки, тем выше способность TiO 2 нанотрубки, способствующие остеогенной дифференцировке МСК. Ингибирование полимеризации F-актина с помощью Cyto D подавляло остеогенную дифференцировку МСК, а также экспрессию белков, содержащихся в комплексах фокальной адгезии, таких как винкулин (VCL) и киназа фокальной адгезии (FAK). Напротив, после обработки Jasp полимеризация F-актина усиливала экспрессию RhoA и факторов транскрипции YAP / TAZ. На основании этих данных мы пришли к выводу, что TiO 2 нанотрубки способствовали остеогенной дифференцировке МСК, и эта способность усиливалась с увеличением диаметра нанотрубок в определенном диапазоне (30–70 В). F-актин опосредовал этот процесс через MKL1 и YAP / TAZ.
Введение
Титан и титановые сплавы из-за их превосходной биосовместимости, коррозионной стойкости и механических свойств широко используются в клинических применениях, таких как полная замена суставов и дентальные имплантаты [1,2,3]. Однако еще предстоит решить множество проблем, включая асептическое расшатывание и инфекцию [4, 5]. В последние годы был проведен ряд исследований, направленных на усиление остеоинтеграции и антибактериальных свойств. Например, MoS 2 / PDA-RGD покрытие на титановых имплантатах может не только способствовать интеграции титанового имплантата с костью-хозяином, но также с высокой эффективностью подавлять рост бактерий [6]. Кроме того, топография поверхности привлекает все больше и больше внимания, и топографическая модификация отличается от химических модификаций только изменением микро- и наноразмерной структуры. Стимуляция химических сигналов на клетках нестабильна и цитотоксична. Для сравнения, безопасные и контролируемые физические сигналы могут избежать некоторых побочных эффектов, вызванных химическими молекулами. Следовательно, топографическая модификация поверхности имплантатов и регулирование остеоинтеграции с помощью топографической структуры могут предоставить новый способ решения клинической проблемы плохой остеоинтеграции после имплантации протезов.
В области инженерии костной ткани и регенерации костей взаимодействие морфологии клеток считается многообещающей стратегией управления для точного контроля функции и дифференцировки семенных клеток. В то же время сама кость имеет элегантную иерархию в нанометровом и микронном диапазоне [7]. Следовательно, морфология поверхности может обеспечить подобную нишу, которая может имитировать естественную структуру кости и способствовать остеогенной дифференцировке мезенхимальных стволовых клеток на поверхности кости-хозяина и имплантата. Морфология поверхности может состоять из множества различных структур, включая нанотрубки, нанопроволоки, нанопоры и так далее. В частности, в последние годы массивы нанотрубок вызвали большой интерес во многих областях из-за их уникальных характеристик поверхности, таких как высокое отношение поверхности к объему, биологическая пластичность и высокая адсорбционная способность. Например, новое исследование показывает, что нанотрубки нитрида бора (BNNT) представляют собой газочувствительный материал, который можно использовать в качестве газового датчика для контроля работы трансформаторов путем определения состава и содержания растворенных газов в масле [8]. В биомедицине топография поверхности также может управлять поведением клеток, включая миграцию, адгезию, пролиферацию и дифференциацию клеток. Последние исследования показывают, что наноразмерная топография может направлять мезенхимальные стволовые клетки (МСК) на дифференцировку в остеобласты, чтобы усилить раннюю остеоинтеграцию [9,10,11,12]. Сообщается даже, что комбинированная модификация поверхности на микро- и нанометрах может вызывать дифференциацию МСК в сократительные гладкомышечные клетки [13]. Однако молекулярные механизмы того, как топография поверхности определяет судьбу клеток, еще предстоит выяснить, что важно для оценки безопасности материалов и дизайна материалов.
Нитчатый (F) -актин, также называемый микрофиламентом, является одним из трех основных компонентов цитоскелета эукариотических клеток. Он состоит из полимеров глобулярного (G) -актина, модифицированного множеством других белков. F-актин имеет структурную полярность из-за того, что все субъединицы микрофиламентов указывают на один и тот же конец. Зубчатый конец направлен на другой соседний мономер, в то время как заостренный конец содержит субъединицу актина с открытым участком связывания АТФ. То есть АТФ участвует в процессе преобразования между G-актином и F-актином. Этот процесс находится в динамическом равновесии с одновременным протеканием полимеризации и деполимеризации, также известного как тредмилинг, который часто наблюдается у ламеллиподий и филоподий [14]. Следовательно, очевидно, что динамика актина играет важную роль в таких клеточных функциях, как миграция клеток, деление клеток и поддержание формы клеток. Однако F-актин не только действует как физическая структура, поддерживающая механическую нагрузку, но также участвует в других биологических процессах, таких как передача сигнала и экспрессия генов. Накапливающиеся данные демонстрируют, что F-актин может преобразовывать физические сигналы в химические сигналы, взаимодействуя с другими белками [15,16,17,18]. Например, биомеханическая и геометрическая реконструкция способствует апоптозу опухолевых клеток, предотвращая наложение полимеризации мономера актина на F-актин [15]. Ультразвуковые импульсы усиливают остеогенез мезенхимальных стволовых клеток человека, ингибируя деполимеризацию F-актина [16]. Наше предыдущее исследование также показало, что механическое напряжение увеличивает стабильность F-актина [17]. Поскольку полая структура нанотрубок обеспечивает меньшее количество участков адгезии для клеток, перестройка цитоскелета неизбежна для поддержания биомеханического баланса. Следовательно, у нас определенно есть основания полагать, что F-актин может опосредовать дифференцировку клеток, индуцированную нанотопографией.
В этом исследовании мы изготовили TiO 2 нанотрубки, изменили свою топографию за счет анодного окисления и исследовали их способность способствовать остеогенной дифференцировке МСК. Затем мы исследовали, играет ли F-актин критическую роль в механотрансдукции. Цитохалазин D (Cyto D), который конкурентно связывается с зазубренным концом F-актина для предотвращения включения G-актина в филамент, был использован для ингибирования полимеризации F-актина, а джасплакинолид (Jasp) был использован для усиления стабилизации сборка актина. Кроме того, мы также хотели выяснить, как F-актин функционирует в превращении физических сигналов в биохимические. Основываясь на результатах нашего предыдущего исследования, мы предположили, что путь MAPK может быть вовлечен в этот процесс [17]. Факторы транскрипции, такие как Yes-связанный белок (YAP) / коактиватор транскрипции с PDZ-связывающим мотивом (TAZ) и MKL1, которые считаются механосенсорами и механотрансдукторами, также были объектами нашего исследования для проверки того, как F-актин влияет на стволовые клетки. судьба, потому что некоторые исследования в других областях предполагают, что они связаны с F-actin [19,20,21]. В целом, мы надеемся прояснить роль F-актина в процессе дифференцировки стволовых клеток, индуцированной нанотрубками, чтобы помочь при разработке материалов и оценке биобезопасности имплантатов, модифицированных нанотрубками.
Материалы и методы
Изготовление TiO 2 Нанотрубки
Срезы чистого титана (чистота 99,9%, толщина 2 мм; Шэншида, Хэбэй, Китай), использованные в качестве подложки, полировали наждачной бумагой из карбида кремния зернистостью № 400 и 1500. Затем образцы промывали последовательно ацетоном, безводным спиртом и деионизированной водой в аппарате для ультразвуковой очистки и, наконец, сушили при комнатной температуре в течение 3 часов. Для создания нанотопографии предварительно обработанные образцы фиксировали в качестве анода, а платиновую деталь в качестве противокатода использовали в водном растворе электролита 0,15 М NH 4 . F и 90% гликоля в течение 1 часа. Напряжение анодирования было постоянным напряжением 30, 40, 50, 60 или 70 В. После анодного окисления каждый образец промывали деионизированной водой в течение 30 минут и промывали безводным спиртом в аппарате для ультразвуковой очистки в течение 15 минут. Наконец, все образцы стерилизовали в автоклаве при 120 ° C в течение 1 часа, а затем увлажняли культуральной средой перед использованием.
Механизм реакции изготовления нанотрубок не ясен, и текущая основная теория - это теория растворения с усилением поля. Формирование массивов нанотрубок является результатом динамического равновесия под действием полевого окисления, полевого растворения и химического растворения (рис. 1б). Процесс анодирования можно описать следующим образом:на первом этапе на границе раздела электролит – металл формируется оксидный барьерный слой:
$$ {\ mathrm {Ti}} ^ {4 +} + 2 {\ mathrm {H}} _ 2 \ mathrm {O} \ to {\ mathrm {Ti} \ mathrm {O}} _ 2 + 4 {\ mathrm { H}} ^ {+} $$ (1)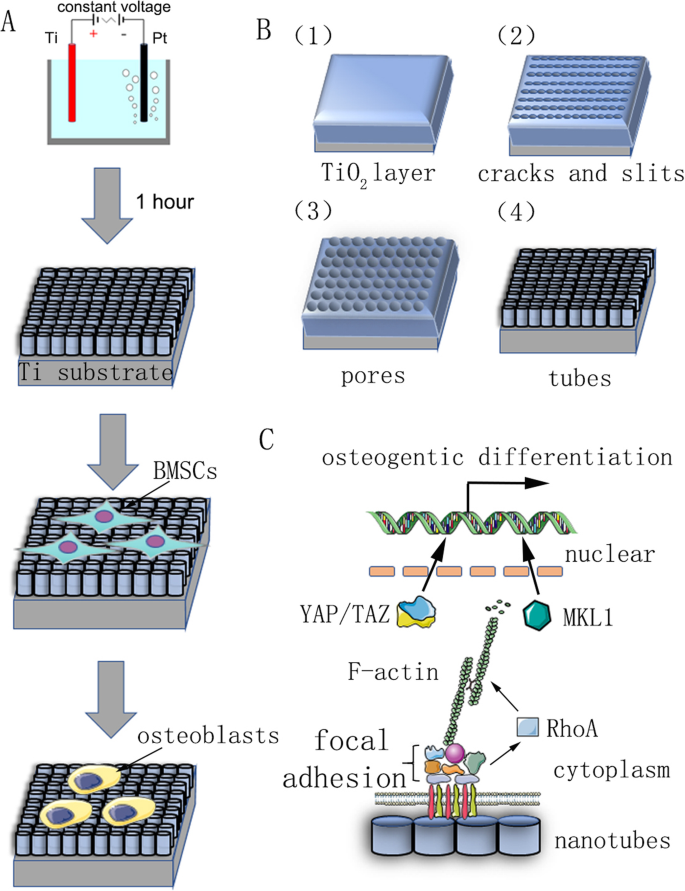
Сводная диаграмма нашего исследования. а Блок-схема, показывающая анодное окисление и индукцию дифференцировки клеток. б Предварительно обработанную титановую подложку фиксировали как анод в водном растворе электролита с 0,15 М NH 4 . F и 90% гликоля при постоянном напряжении в течение 1 часа. Было предсказано, что самоорганизующиеся нанотрубки будут формироваться однородно. Механизм его реакции описан в материалах и методах. c Принципиальная схема механизма остеогенной дифференцировки стволовых клеток, индуцированной нанотрубками
Затем на поверхности появляются трещины и узкие щели из-за растворения оксидного слоя под действием поля. Распространение F - попадание ионов в эти трещины и щели увеличивает скорость растворения. Трещины увеличиваются и соединяются с соседними трещинами. Наконец, скорость образования и скорость растворения слоя оксида титана достигают динамического баланса, и нанотрубки больше не растут:
$$ {\ mathrm {TiO}} _ 2 + 6 {\ mathrm {F}} ^ {-} + 4 {\ mathrm {H}} ^ {+} \ to {\ left [{\ mathrm {TiF}} _ 6 \ right]} ^ {2 -} + 2 {\ mathrm {H}} _ 2 \ mathrm {O} $$ (2)Характеристика поверхности
Образцы, изготовленные с различным напряжением (30, 40, 50, 60 или 70 В), промывали этанолом и деионизированной водой в течение 15 мин, затем сушили при комнатной температуре. Сканирующая электронная микроскопия (SEM450, FEI Nova Nano SEM; Thermo Fisher Scientific, Waltham, MA, USA) использовалась для характеристики структуры поверхности и для измерения внутреннего диаметра и высоты нанотрубок после покрытия образцов тонким слоем золота. Между тем, для анализа элементного состава нанотрубок был проведен рентгеновский энергодисперсионный анализ (EDS). Атомно-силовая микроскопия (AFM, NanoManVS, Bruker Nano Surfaces, Bruker MicroCT, Kontich, Бельгия) использовалась для исследования морфологии и шероховатости поверхности образцов. Из каждого образца были выбраны три разные области, и измерения были повторены трижды.
Культура клеток
Четырехнедельные самцы крыс Sprague-Dawley (SD) были приобретены в центре экспериментальных животных Шанхайской девятой народной больницы (Шанхай, Китай). Мезенхимальные стволовые клетки костного мозга крысы (BMSC) выделяли в асептических условиях из бедренных и большеберцовых костей. BMSC очищали и далее размножали в α-минимальной эссенциальной среде (α-MEM; Hyclone, Logan, UT, USA), содержащей 10% (об. / Об.) Фетальной бычьей сыворотки (FBS) (Gibco / Life Technologies, Карлсбад, Калифорния, США). ), 100 мг / мл стрептомицина (Gibco) и 100 Ед / мл пенициллина (Gibco), и инкубировали при 37 ° C в увлажненной атмосфере, состоящей из 95% воздуха и 5% CO 2 . Культуральную среду обновляли каждые 2 дня, клетки обрабатывали трипсином и субкультивировали при 80% конфлюэнтности. Все клетки, использованные в этом исследовании, находились между пассажами 3 и 5. Среда для остеогенной индукции состояла из ростовой среды с добавлением 100 нМ дексаметазона, 10 мМ β-глицерофосфата и 50 мМ аскорбиновой кислоты (Sigma-Aldrich, Сент-Луис, Миссури, США). .
Анализ пролиферации клеток
TiO 2 Срезы титана, модифицированные нанотрубками, нарезали круглой формы и помещали в лунки 24-луночного планшета для культивирования клеток. BMSC между пассажами 3-5 культивировали на TiO 2 . нанотрубки с плотностью 3 × 10 4 клетки / диск в среде для выращивания или в остеогенной среде. Через 2 дня культивирования клеток в среду добавляли цитохалазин D (Cyto D, Sigma – Aldrich) и джасплакинолид (Jasp, Sigma – Aldrich), используемые для нарушения полимеризации F-актина, каждый день в течение 3 дней. Конечная концентрация и рабочее время Cyto D и Jasp составляли 5 мкМ, 1 час и 2 мкМ, 3 часа соответственно. Культуральную среду обновляли после инкубации с реагентами. Жизнеспособность и пролиферацию клеток оценивали с помощью теста Cell Counting Kit-8 (CCK8) (Dojindo, Kumamoto, Japan) через 12 часов после обработки Cyto D или Jasp. Клетки инкубировали с 10% (об. / Об.) Раствором CCK8 в течение 2 часов в инкубаторе для клеток при 37 ° C и 5% CO 2 . . Затем мы перенесли 100 мкл реакционной смеси в лунки 96-луночного планшета и оптическую плотность (OD) формазанового красителя в культуре измеряли при 450 нм с помощью спектрофотометра Multiscan UV-visible (Safire2; TECAN, Mannedorf, Швейцария). ). Кроме того, мы также подсчитали приблизительное количество ячеек с помощью автоматического счетчика ячеек (AMQAX1000, Life Technologies). Перед подсчетом клеток BMSC были ферментативно отделены от TiO 2 нанотрубок и окрашены трипановым синим (Sigma – Aldrich).
Окрашивание щелочной фосфатазой и анализ активности ЩФ
BMSC были засеяны на пять разных TiO 2 срезы титана, модифицированные нанотрубками (30, 40, 50, 60 или 70 В) с плотностью 3 × 10 4 на лунку и культивировали в остеогенной среде. Реагенты добавляли, как описано выше. После 7 дней инкубации BMSC культивировали на TiO 2 нанотрубки трижды промывали PBS, фиксировали 4% параформальдегидом и инкубировали в рабочем растворе щелочной фосфатазы (ЩФ) набора ЩФ в соответствии с инструкциями производителя (Хунцяо, Шанхай, Китай). Результаты наблюдались под стереомикроскопом после промывки PBS.
Для анализа активности ЩФ клетки сначала лизировали буфером RIPA без ингибиторов протеаз и фосфатаз, а затем центрифугированные лизаты анализировали с помощью набора для анализа ЩФ (Beyotime Institute of Biotechnology, Цзянсу, Китай) в соответствии с предоставленным протоколом. В конечном итоге активность была нормализована до концентрации белка в соответствующем лизате.
Иммуноцитохимия
После 3 дней обработки Cyto D и Jasp BMSC фиксировали 4% параформальдегидом в течение 20 минут при комнатной температуре, а затем трижды промывали PBS. Клетки были проницаемы 0,3% Triton-X 100 в течение 30 мин, трижды промыты PBS и окрашены фаллоидином, конъюгированным с родамином, в течение 1 ч при комнатной температуре в темноте. Затем клетки промывали PBS и контрастировали DAPI (Beyotime Institute of Biotechnology) в течение 10 мин при комнатной температуре. После трех дополнительных промывок PBS образцы фиксировали на предметном стекле и наблюдали с помощью конфокальной микроскопии.
Вестерн-блоттинг
Чтобы оценить экспрессию белка, BMSC культивировали на TiO 2 нанотрубки собирали с трипсином (Gibco). Клетки трижды промывали PBS и лизировали буфером RIPA с добавлением смеси ингибиторов протеаз и фосфатаз в течение 30 мин на льду. Лизат собирали центрифугированием при 12000 × g в течение 15 мин при 4 ° C. Концентрацию общих белков в супернатанте измеряли с использованием набора для анализа белков бицинхониновой кислоты (BCA) (Beyotime) в соответствии с инструкциями производителя. Загрузочный буфер добавляли к образцам белка, как указано выше, и затем их кипятили при 95 ° C в течение 15 минут. Для вестерн-блоттинга 10 мкл белкового препарата загружали в 12,5% гель SDS-PAGE (EpiZyme Inc., Кембридж, Массачусетс, США) и подвергали электрофорезу при 120 В в течение 1 ч, затем переносили электропереносом на поливинилидендифторид. (PVDF) мембрана при 250 мА в течение 2 часов. Затем мембраны блокировали 5–10% обезжиренным сухим молоком в TBST в течение 1 ч на шейкере при комнатной температуре и инкубировали с первичным антителом, разведенным в буфере для разведения (Beyotime), при 4 ° C в течение ночи. Затем флуоресцентно-конъюгированные вторичные антитела, разведенные в буфере для разведения, добавляли к мембранам после трехкратной промывки TBST в течение 5 минут, а затем мембраны инкубировали при комнатной температуре в течение 1 часа в темноте. Полосы белка детектировали с помощью системы двухцветной инфракрасной флуоресцентной визуализации (Odyssey, LiCor Biosciences, Lincoln, NE, USA). В частности, если полосы внутреннего эталонного белка были объединены, мембрану отделяли и повторно зондировали другим первичным антителом с последующим тем же процессом. Мы использовали антитело GAPDH в качестве внутреннего эталонного белка для нормализации экспрессии белка, а другие первичные антитела, использованные в этом исследовании, были анти-винкулин (разведение 1:1000, Abcam, Кембридж, Массачусетс, США), анти-FAK (разведение 1:1000). , Cell Signaling Technology, Дэнверс, Массачусетс, США), anti-Runx2 (разведение 1:1000, Cell Signaling Technology), anti-RhoA (разведение 1:1000, Cell Signaling Technology), анти-F-актин (разведение 1:500). , Abcam), anti-Osx (разведение 1:500, Abcam) и anti-pYAP (разведение 1:1000, Cell Signaling Technology). Вторичными антителами были козьи антимышиные IgG H&L (IRDye® 680RD, разведение 1:5000, Abcam) и козьи антикроличьи IgG H&L (IRDye® 680RD, разведение 1:5000, Abcam).
Количественная ПЦР в реальном времени
Количественная ПЦР в реальном времени была проведена на 7 день для оценки экспрессии генов фактора транскрипции 2, связанного с runt (Runx2), Osterix (Osx), Alp, остеокальцина (OCN), RhoA, YAP, TAZ, винкулина (VCL), киназа фокальной адгезии (FAK) и мегакариобластный лейкоз 1 (MKL1) в клетках, выращенных в остеогенной среде на TiO 2 нанотрубки. Тотальную РНК экстрагировали из клеток с использованием набора Total RNA Kit (R6812-01HP, Omega Bio-Tek Inc., Norcross, GA, USA). Концентрацию и чистоту образцов РНК определяли по оптической плотности на длине волны 260, и анализировали только образцы, имеющие как отношения A260 / 280, так и отношения A260 / 230 выше 1,8. Образцы РНК подвергали обратной транскрипции в кДНК с использованием набора для синтеза кДНК qScript (Takara, Shiga, Japan) в соответствии с инструкциями производителя. Количественная ПЦР в реальном времени была проведена с SYBR® Premix Ex Taq ™ (Takara) с использованием системы ПЦР в реальном времени QuantStudio 6 Flex (Life Technologies). GAPDH, ген домашнего хозяйства, использовался в качестве внутреннего эталона. Данные были проанализированы с использованием сравнения Ct (2 -ΔΔCt ) и выражается в кратных изменениях по сравнению с контролем. Последовательности использованных праймеров перечислены в таблице 1.
Статистический анализ
Все данные являются репрезентативными по крайней мере для трех независимых экспериментов с использованием трех образцов, если не указано иное. Данные выражены в виде среднего значения ± стандартное отклонение (SD). Различия между группами оценивали с помощью одностороннего дисперсионного анализа с последующим апостериорным тестом Стьюдента-Ньюмана-Кеулса или t Стьюдента. контрольная работа. P значения <0,05 считались статистически значимыми.
Результаты
Характеристика поверхности
Чтобы изготовить нанотопографию, TiO 2 Нанотрубки были сформированы на чистой титановой подложке с использованием оборудования для анодного окисления при различных постоянных напряжениях (30, 40, 50, 60 и 70 В) в течение 1 ч (рис. 1). Равномерно распределенный массив самособирающихся нанотрубок наблюдался с помощью сканирующей электронной микроскопии (SEM). Виды на нанотрубки сбоку и сверху показаны на рис. 2а, б. Высота нанотрубок во всех образцах в этом исследовании составляла приблизительно 2 мкм, в то время как внутренний диаметр нанотрубок составлял приблизительно 74 нм (30 В), 92 нм (40 В), 112 нм (50 В), 128 нм (60 В). V) и 148 нм (70 В) (рис. 2в). Это указывало на то, что высота нанотрубок была связана со временем анодного окисления, а внутренний диаметр был связан с напряжением анодного окисления. Затем был проведен рентгеновский энергодисперсионный анализ (EDS) для анализа элементного состава нанотрубок. Это показало, что нанотрубки состоят только из двух элементов, O и Ti (рис. 2d, e). Атомно-силовая микроскопия (АСМ) использовалась для обнаружения нанотрубчатых структур и измерения среднего арифметического отклонения профиля нанотрубок как шероховатости поверхности (Ra) (рис. 2f). Данные показали, что шероховатость поверхности нанотрубок увеличивается с увеличением диаметра (т. Е. Напряжения анодного окисления) (рис. 2g).
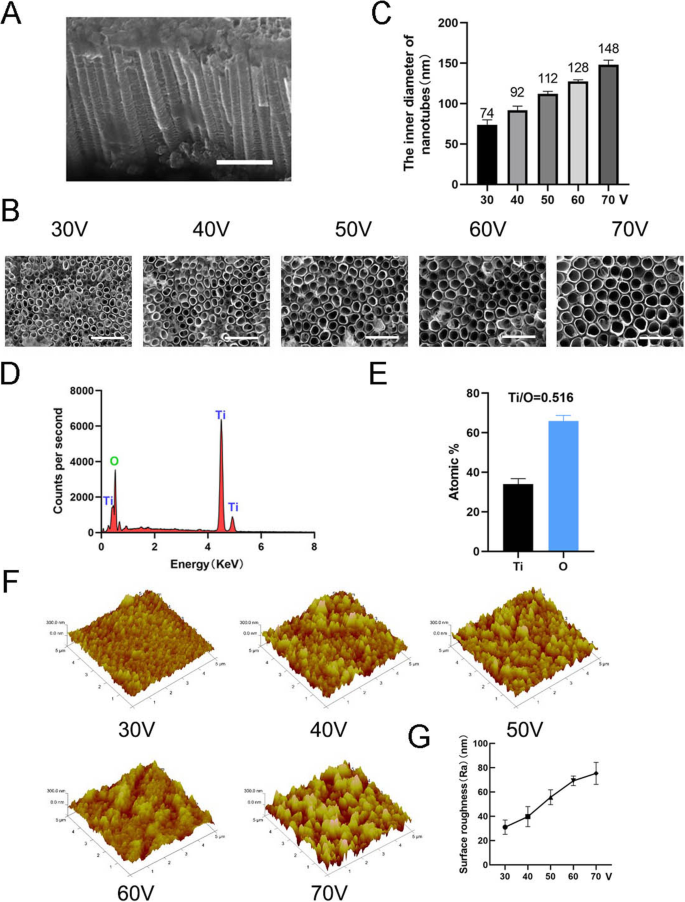
Характеристика поверхности нанотрубок. а Вид сбоку на нанотрубки. Масштабные линейки:1 мкм. б Вид сверху нанотрубок. Масштабные линейки:500 нм. c Внутренний диаметр нанотрубок при пяти различных постоянных напряжениях (30, 40, 50, 60, 70 В). г Химический элементный состав EDS структур нанотрубок. е Соотношение элементов Ti и O. f Изображения структур нанотрубок с помощью атомно-силовой микроскопии (АСМ). г Средняя шероховатость поверхности (Ra) нанотрубок
TiO 2 Нанотрубки индуцировали остеогенную дифференцировку МСК
После 7 дней остеоиндукции сначала проводили окрашивание ALP для оценки остеогенной дифференцировки МСК. Результаты окрашивания показали, что МСК, культивированные на TiO 2 нанотрубки обладали более высокой активностью ЩФ, чем клетки, культивируемые на гладкой титановой подложке (контрольная группа) (рис. 3а). Статистический анализ области окрашивания продемонстрировал, что способность нанотрубок вызывать остеогенную дифференцировку была значительно увеличена по сравнению с контрольной группой. Между тем, мы наблюдали тенденцию, согласно которой в пределах диапазона диаметров этого эксперимента, чем больше диаметр TiO 2 нанотрубки, тем сильнее способность вызывать остеогенную дифференцировку (рис. 3b). Таким образом, группа 70 В использовалась в последующих экспериментах для лучшего отображения результатов. Затем мы проанализировали экспрессию остеогенных генов на 3-й и 7-й дни. МСК, культивированные на TiO 2 нанотрубки в течение 3 и 7 дней показали значительное усиление экспрессии остеогенных генов (RUNX2, ALP, OCN и OSX) по сравнению с контрольной группой (рис. 3d – g). Результаты вестерн-блоттинга подтвердили, что экспрессия белков RUNX2 и OSX также увеличивалась после 7 дней остеоиндукции (фиг. 3c). Интересно, что мы обнаружили, что F-актин активируется в TiO 2 группа нанотрубок. Следовательно, было очевидно, что TiO 2 нанотрубки направляли МСК к дифференцировке остеобластов, что было связано с диаметром нанотрубок. Наши результаты также предполагают участие F-актина в этом процессе.
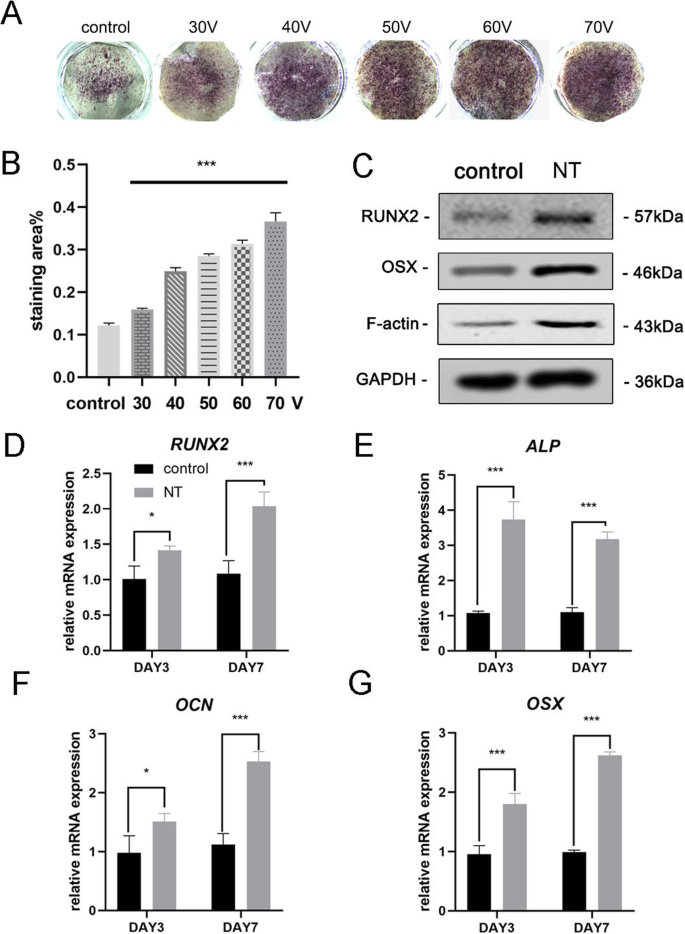
TiO 2 нанотрубки усиливали экспрессию остеогенных генов BMSC. а Окрашивание ALP гладкой титановой подложки и пяти различных подложек нанотрубок. Клетки индуцировали остеогенной средой в течение 7 дней. б Статистический анализ области окрашивания проводили с помощью ImageJ. c Связанные с остеогенезом белки (RUNX2 и OSX) и F-актин в МСК анализировали с помощью вестерн-блоттинга на 7 день. Экспрессия мРНК RUNX2 ( d ), ALP ( e ), OCN ( f ) и OSX ( g ) на 3-й и 7-й дни, анализировали с помощью qRT-PCR. NT группа нанотрубок. Данные представляют собой среднее значение ± стандартное отклонение трех образцов. * P <0,05, ** P <0,01 и *** P <0,001
Дифференциация остеобластов, опосредованная F-актином, на TiO 2 Нанотрубки
Чтобы дополнительно изучить, участвует ли F-актин в дифференцировке клеток, индуцированной нанотопографией, мы использовали два реагента, ясплакинолид (Jasp) и цитохалазин D (Cyto D), чтобы регулировать полимеризацию F-актина положительным и отрицательным образом соответственно. Конфокальные микрофотографии окрашивания родамином-фаллоидином показали, что F-актин в группе лечения Cyto D был почти деполимеризован и фиброзные структуры наблюдались редко, в то время как Jasp стабилизировал и полимеризовался F-актин, что подтверждается более отчетливыми и яркими пучковидными структурами, чем наблюдаемые. в контрольной группе (рис. 5а). Кроме того, вестерн-блоттинг также подтвердил влияние на экспрессию белка F-актина, который доказал, что Cyto D и Jasp действовали, как ожидалось (фиг. 5b). Анализ пролиферации клеток показал, что Cyto D значительно ингибирует пролиферацию клеток, в то время как Jasp способствует росту клеток (рис. 4a). Результаты подсчета клеток согласуются с этим выводом (рис. 4b).
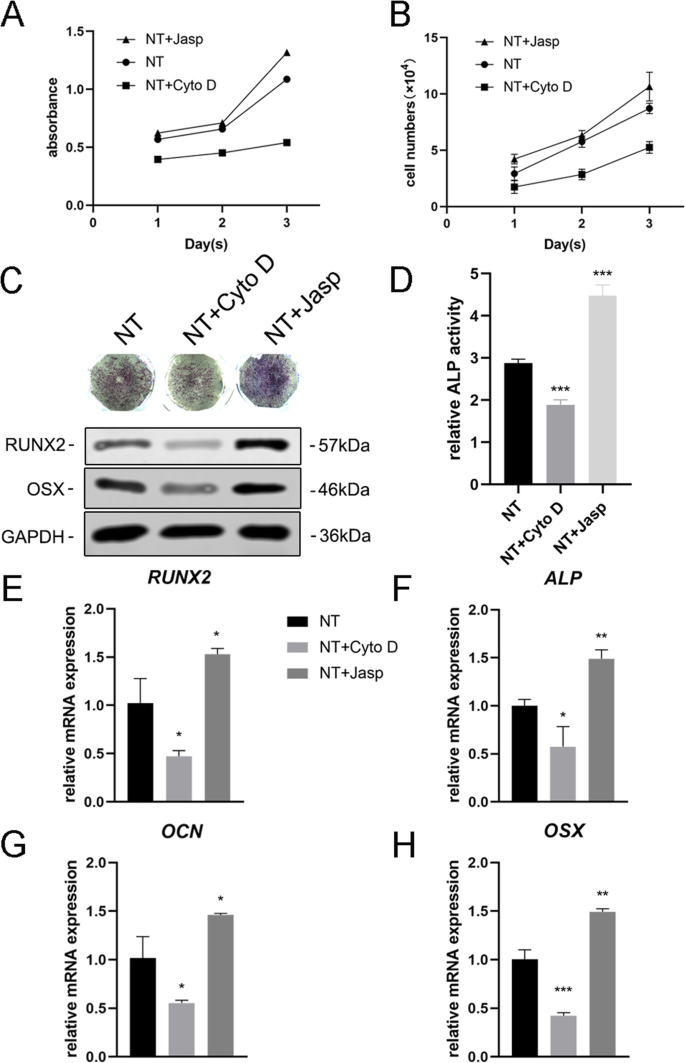
Сборка F-актина регулирует экспрессию остеогенных генов в BMSC. а , b Пролиферацию клеток после обработки Cyto D и Jasp определяли с помощью анализа CCK-8 или автоматического счетчика клеток в дни 1, 2 и 3. c , d Окрашивание ALP и анализ активности ALP выполняли для оценки экспрессии ALP в МСК после лечения препаратом в течение 3 дней. Область окрашивания анализировали с помощью ImageJ. c , e - ч Вестерн-блоттинг и qRT-PCR использовали для сравнения изменений маркеров, связанных с остеогенезом, в группе NT + Cyto D и группе NT + Jasp с таковыми в контрольной группе (без лекарственной обработки). NT группа нанотрубок. Данные представляют собой среднее значение ± стандартное отклонение трех образцов. * P <0,05, ** P <0,01 и *** P <0,001
Затем мы оценили способность МСК дифференцироваться в остеобласты, чтобы выяснить, опосредует ли F-актин этот процесс. Мы впервые обнаружили ALP как ранний маркер остеогенеза. По сравнению с контрольной группой обработка Cyto D снижала экспрессию ALP и ее активность, в то время как в группе обработки Jasp была повышена регуляция (фиг. 4c, d). В соответствии с этим результатом обработка Jasp приводила к увеличению уровней белка RUNX2 и OSX, тогда как Cyto D оказывал противоположный эффект (рис. 4c). В соответствии с этим, уровни экспрессии мРНК остео-специфических генов, включая RUNX2, ALP, OCN и OSX, показали такую же тенденцию после лечения лекарствами (рис. 4e-h). Прежде всего, эти данные указывают на то, что F-актин играет важную роль в процессе остеогенной дифференцировки МСК, индуцированной TiO 2 нанотрубки. Содействие деполимеризации F-актина ингибировало индуцированную нанотопографией дифференцировку остеобластов, в то время как стабилизация и полимеризация F-актина усиливали дифференцировку остеобластов.
Дифференциация остеобластов, регулируемых F-актином, на TiO 2 Нанотрубки через MKL1 и YAP / TAZ
Чтобы проанализировать лежащий в основе механизм с участием F-актина в регуляции судьбы МСК, мы исследовали белки / молекулы, которые напрямую взаимодействуют с F-актином или влияют на полимеризацию F-актина. Во-первых, мы попытались определить, как нанотопография влияет на баланс между F-актином и G-актином. TiO 2 нанотрубки как физический сигнал отличаются от проницаемых через мембрану химических сигналов и, следовательно, должны использовать некоторые компоненты клеточной мембраны для передачи стимулов в клетки. Накапливающиеся данные показывают, что комплекс фокальной адгезии, включая интегрин, талин, киназу фокальной адгезии (FAK), винкулин (VCL), тензин и другие белки, функционирует как носитель сигнала, который информирует клетки о состоянии внеклеточного матрикса (ECM). ) и тем самым влияет на их биологическое поведение [22, 23]. Что еще более важно, F-actin связывается с интегринами посредством таких комплексов фокальной адгезии и тем самым формирует механические связи между внутриклеточными пучками актина и ECM [24]. Следовательно, мы затем проанализировали экспрессию компонентов фокального адгезионного комплекса. Результаты показали, что экспрессия белка и мРНК VCL и FAK соответствовала изменению F-актина, что указывает на то, что комплекс фокальной адгезии участвует в процессе остеогенной дифференцировки МСК, индуцированной TiO 2 нанотрубки (рис. 5б, 6а, б). Кроме того, мы также обнаружили, что RhoA, небольшой белок GTPase в семействе Rho GTPases, активируется в группе лечения Jasp и ингибируется Cyto D (фиг. 5b и 6a, b). RhoA является важной вышестоящей молекулой для передачи сигнала в пути MAPK и может регулироваться с помощью FAK [25, 26]. Основная функция RhoA - способствовать полимеризации и стабильности стрессовых волокон (F-актин) и сборке фокального адгезионного комплекса [27]. Взятые вместе, эти данные позволяют предположить, что TiO 2 нанотрубки могут влиять на полимеризацию F-актина через комплекс фокальной адгезии и RhoA.
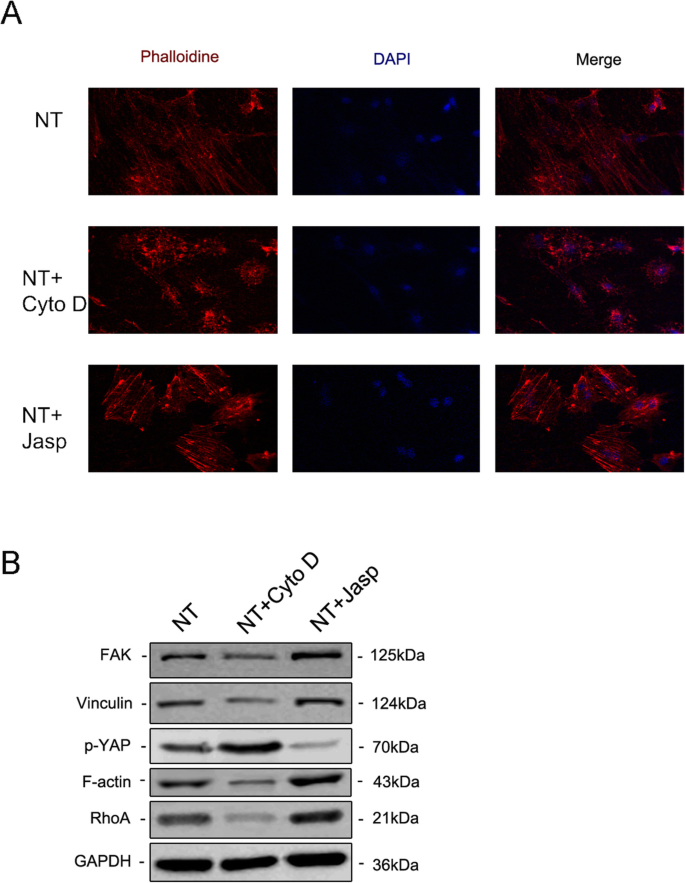
Иммунофлуоресцентное окрашивание выявило уровень F-актина путем окрашивания фаллоидином, конъюгированным с родамином ( a ). The protein expression of FAK and VCL contained in the focal adhesion complex, RhoA and phosphorylated YAP were investigated by Western blotting (b ). NT the nanotubes group

Effect of F-actin assembly on FAK (a ), vinculin (VCL) (b ), RhoA (c ), MKL1 (d ), YAP (e ), and TAZ (f ) gene expression in MSCs. NT the nanotubes group. Data represent the mean ± SD of three samples. * P <0.05, **P <0.01, and ***P <0.001
But how does F-actin regulate cell fate? Most studies have demonstrated that F-actin is involved in cell migration, cell division, endocytosis, and especially tumor cell invasion [28,29,30]. Few studies have suggested that F-actin could also regulate cell differentiation, let alone its specific molecular mechanism [31, 32]. Consequently, we searched for articles that mentioned the F-actin changes and found that YAP/TAZ, two closely related transcriptional co-activators in the Hippo signaling pathway, which shuttle between the cytoplasm and the nucleus, may serve as mechanotransducers in regulating MSC differentiation [33,34,35]. In addition, we also found that MKL1, a key regulator of smooth muscle cell differentiation, which interacts with the transcription factor serum response factor, could bind to G-actin and also circulate between the cytoplasm and the nucleus [21, 36]. Our results ultimately identified the involvement of YAP/TAZ and MKL1 in nanotube-induced osteoblast differentiation mediated by F-actin (Figs. 5b and 6d–f). Interestingly, the protein expression of phosphorylated YAP showed the opposite trend, indicating that not only was the expression of YAP changed, but the phosphorylation of YAP was also changed by Cyto D and Jasp (Fig. 5b). This result was consistent with the report that the phosphorylation of YAP/TAZ could be sequestrated in the cytoplasm [35].
In summary, our results preliminarily demonstrated that F-actin regulated osteoblast differentiation of MSCs on TiO2 nanotubes through MKL1 and YAP/TAZ (Fig. 7).

Schematic representation of F-actin assembly induced by nanotubes, and the putative role of MKL1 and YAP/TAZ in acting as the downstream mediators of F-actin signaling to regulate gene expression
Discussion
Titanium and titanium alloys are the most widely used metal materials in orthopedic clinical implants due to the good properties of titanium [3]. However, aseptic loosening is still an urgent problem to be solved and improved, and the key is likely to lie in improving the integration of the implant and the host bone. Previous studies have shown that surface coating and modification or immobilization of biofunctional molecules will be beneficial to osseointegration [37]. Recently, the surface topography of implants has attracted the attention of many researchers thanks to studies into the cell response to physical cues [9,10,11, 13, 38]. In this study, we demonstrated the ability of nanotubes to promote osteogenic differentiation of MSCs, and this ability was enhanced with increasing inner diameter of the nanotubes (30–70 V). This will help guide the diameter of the nanotubes on the surface of the implants.
As a topographical structure, nanotubes first change the physical properties of the material, such as adsorption capacity and electrical and thermal conductivity. These physical properties determine their application in the industrial field. For example, most high-voltage power transformers need to be filled with insulating material, which is usually transformer oil or insulating gas. When the insulation of a transformer fails due to overheating and partial discharge, a serious discharge accident will occur. Therefore, finding an effective method that accurately detects the concentration and types of dissolved gases or insulating gas decomposition components in a transformer is necessary to monitor the operating state of the transformer [8, 39,40,41]. The traditional approach is to look for materials with good gas adsorption in transition elements, which are rich in d electrons, such as Pd(1 1 1) [39]. Nowadays, nanotubes are widely studied for their good gas adsorption properties. He et al. found that CuO-BNNT was suitable for the adsorption of C2 H 2 , because of its stronger adsorption on C2 H 2 [8]. Meanwhile, TiO2 itself can be a gas-sensing material. Gui et al. found that Co-doped TiO2 further enhanced gas adsorption capacity and exhibited a superior adsorption ability and conductivity change toward C2 H 4 molecules [40]. Consistent with this study, Mn-doped graphene also exhibited enhanced conductivity and superior capability of C2 H 2 and CO detection than pristine graphene [41]. The above research indicates that the TiO2 nanotubes prepared in our experiment have a potential application in the field of monitoring the operative state of a transformer. However, the adsorption capacity and electrical conductivity of the nanotubes to gases need to be further studied, especially whether these properties are enhanced after doping with transition elements (e.g., Mn).
In addition to changing the physical properties of a surface, nanoscale morphology also affects the biological behavior of the cells attached to it. Cells first adhere to the surface of the material and then migrate, proliferate, and differentiate. Compared to a flat surface, the hollow structure of the nanotubes provides fewer adhesion sites for cells. Therefore, in order for the cells to adhere to the nanotube surface steadily and maintain the biomechanical balance within the cell, the focal adhesion complex begins to assemble and mature, and F-actin becomes strong and stable.
F-actin, a linear polymer microfilament consisting of G-actin monomers, is one of the three major components of the cytoskeleton. As a mechanical-loading structure, F-actin is generally believed to be involved in cell division, cell migration, endocytosis, and tumor cell invasion [28,29,30], but some recent studies showed that it can also affect cell differentiation [31,32,33, 36]. For example, actin cytoskeletal depolymerization by simvastatin induces chondrocyte differentiation [31], and actin depolymerization enhances adipogenic differentiation in human stromal stem cells [32]. Our results also revealed that, compared with the control group, MSCs cultured on nanotubes had higher F-actin levels and a more obvious fibrous structure. Meanwhile, promotion of F-actin polymerization by Jasp enhanced osteogenic differentiation, while the depolymerization of F-actin inhibited osteogenic differentiation, suggesting that F-actin mediates TiO2 nanotube-induced osteoblastic differentiation of MSCs.
F-actin can be regulated by Rho GTPases, members of the Ras superfamily [23, 42], and Rho can induce actin reorganization through at least two effectors, ROCK and Dia. ROCK is activated by binding to Rho-GTP and then myosin light chain (MLC), the substrate of ROCK, plays an important role in F-actin assembly. ROCK inhibits the activity of MLC phosphatase, leading to an increase in MLC phosphorylation, which stimulates the ATPase activity of myosin II and promotes the assembly of F-actin. In addition, ROCK also targets LIM kinase (LIMK). Phosphorylated LIMK inactivates cofilin by phosphorylation, which can disassemble F-actin in its active state. Another effector is Dia, a member of the formin-homology (FH) family of proteins which contains two FH domains. These domains contain multiple proline-rich motifs which bind to the G-actin-binding protein, profilin. This interaction contributes to actin polymerization and F-actin organization [42]. We detected one of the Rho GTPases, RhoA, and found that the expression of RhoA was consistent with the level of F-actin. However, we were unable to clearly describe how the nanotubes regulate the expression of RhoA, because there are many other regulators, including integrin signaling, other adhesion receptors, G protein-coupled receptors (GPCRs), soluble factors such as LPA, receptor tyrosine kinase signaling, and so on [43].
Knowing that F-actin can be regulated by RhoA, we next asked what role focal adhesion played in this process, because focal adhesion complexes, containing integrins, talin, vinculin, paxillin, and focal adhesion kinase (FAK), are formed and mature when cells attach to the surface of nanotubes. Integrins are transmembrane heterodimers that couple the ECM to the other focal adhesion proteins so as to facilitate cell attachment. They not only act simply as hooks but also transmit to the cell critical signals about the nature of its surroundings, which along with other signals such as EGFR, prompt the cell to make decisions about its biological behaviors. These signals are further transmitted to F-actin, which is directly connected to the focal adhesion complexes. On the one hand, the nanoscale morphology causes focal adhesion complex assembly and maturation. On the other hand, kinases such as FAK and Src kinase family members will recruit molecules such as CRK to self-regulate the assembly and maturation of focal adhesion complexes [44,45,46]. Our results demonstrated that the formation and maturation of focal adhesion complexes were impaired by F-actin depolymerization, suggesting that there was a feedback from focal adhesion complexes to actin assembly in line with published reports.
However, it should not be ignored that these proteins contained in focal adhesion complexes have the function of signal transduction [47]. That is to say, nanotubes may directly regulate gene expression through signal cascades, and F-actin may just participate in or be affected by this process. For instance, the dual kinase complex of FAK and Src can regulate Rho GTPases such as RhoA. This shows that nanotubes can regulate RhoA through integrins and the FAK/Src complex. In addition Src, a non-receptor tyrosine kinase protein, can activate Ras (small GTPase) by phosphorylating FAK at tyrosine residue 925 [47, 48]. Then, Ras activates numerous biochemical pathways, including the well-studied MAPK pathway and the PI3K/AKT/mTOR pathway. In the MAPK pathway, Ras activates c-Raf, followed by mitogen-activated protein kinase kinase (MAP2K) and then MAPK1/2, also known as extracellular signal-regulated kinase (ERK). ERK in turn activates transcription factors such as serum response factor (SRF) and c-Myc that are involved in regulating growth and differentiation [49]. What is more, Runx2, a key transcription factor in osteogenic differentiation, can also be regulated by ERK [50], and our previous study confirmed that mechanical strain promoted osteogenic differentiation of BMSCs through the FAK-Erk1/2-Runx2 pathway [17]. Therefore, we cannot rule out that ERK plays a role in nanotube-induced osteogenic differentiation and further study is still needed.
So what exactly is the role of F-actin in inducing differentiation of nanotubes, because its change can affect cell differentiation? One possibility is that the change of F-actin assembly can inversely regulate the level of FAK so as to induce osteogenic differentiation through the FAK-Erk1/2-Runx2 pathway as described above, because in our results, focal adhesion complexes and actin polymerization showed the same trends of change, indicating that they act as a whole in response to the extracellular environment. However, some other possibilities also exist, and a number of articles have shown that MKL1 and YAP/TAZ act downstream of the actin dynamic balance [20, 51,52,53,54]. Both of them shuttle between the cytoplasm and the nucleus, and may help to transduce signals from the cytoskeleton to the nucleus.
MKL1, also termed myocardin-related transcription factor A, is sensitive to changes in G-actin levels. When cytoplasmic G-actin levels increase, monomeric G-actin binds to MKL1 and prevents it from binding to SRF and activating transcription. SRF target genes include actins such as smooth muscle actin (SMA) as well as other actin-binding proteins, including immediate early genes like c-fos and egr1. Recent studies have demonstrated that changing SRF activity could regulate adipogenesis by activating the adipogenesis transcription factor peroxisome proliferator-activated receptor γ (PPARγ), and also regulate bone formation via IGF-1 and Runx2 signaling [55, 56].
YAP and TAZ are two transcriptional coactivators in the Hippo signaling pathway, identified as an important regulatory pathway that restricts cell proliferation, thereby controlling organ size and morphogenesis [20]. Large tumor suppressor genes 1 and 2 (LATS1/2) phosphorylate them, thereby creating a binding site for 14-3-3 proteins, the binding of which prevents their nuclear import [53, 54]. As a consequence, phosphorylated forms of YAP/TAZ are sequestered in the cytoplasm, preventing the expression of genes like Ctgf and Areg. In addition, some studies have shown that YAP/TAZ can interact with T-box 5 (TBX5), RUNX2, and p73 to regulate gene expression [57,58,59]. Further, cell adhesion to cell matrix proteins has been shown to trigger YAP nuclear localization through an integrin/FAK/Src axis. In our study, the results suggested that this pathway was possibly involved in nanotube-induced differentiation. Further study into the downstream mediators of the integrin/FAK/Src axis should be carried out to clarify the specific mechanism.
On the other hand, more and more studies illustrate that F-actin interacts with Hippo signaling, and somehow inhibits the phosphorylation of YAP [54, 60], which is consistent with our experimental results that promoting F-actin polymerization reduces the expression of phosphorylated YAP. We hypothesize that ATP involved in the process of the transformation between G-actin and F-actin may also play an important role in the phosphorylation of YAP, which is yet to be studied.
After understanding the above possible molecular mechanisms, we can try to explain some of the experimental phenomena found in this study. Our results revealed that the larger the diameter of the nanotubes, the stronger the ability of the nanotubes to promote osteogenic differentiation. This is consistent with previous research [61, 62]. The reason for this phenomenon is that the larger the diameter of the nanotubes, the less adhesion sites they can provide to the cells, and the greater the assembly and maturity of focal adhesion complexes. Along with these, stress fibers made of F-actin will have greater strength and stability. These structures enhance the signaling that promotes osteogenic differentiation. Predictably, however, this effect is significantly reduced when the nanotubes become too large in diameter, making it difficult for the cells to adhere to the surface [12]. Similarly, when the height of the nanotubes is inconsistent, the differences in height can result in a change of adhesion site and rearrangement of the cytoskeleton, which will further affect cell differentiation. Intriguingly, even flat surface materials without nanotube modification can induce changes in cell differentiation. A number of studies have demonstrated that focal adhesion formation and stress fiber organization are regulated by substrate stiffness [63,64,65], and YAP/TAZ also plays an important role in this process. Therefore, it is obvious that the integrins–FAs (focal adhesions)–F-actin axis plays a role in the transduction of physical signals into intracellular chemical signals.
In summary, our results demonstrated that F-actin regulates osteoblastic differentiation of mesenchymal stem cells on TiO2 nanotubes through MKL1 and YAP/TAZ, whose target genes partly explained the proliferation and differentiation of MSCs. We know that there is no single change in the signal network and any change is regulated by numerous molecules and proteins. One type of biological behavior must be the result of the regulation of a series of signaling pathways. Nanotubes induce cell differentiation by triggering a complex network of signals, including integrins, proteins contained in focal adhesion complexes, FAK, Src, Rho GTPase, the MAPK pathway, the Hippo pathway, and other reported signaling pathways. At least as important, there are many signal cycles in the signal network and a downstream signal can regulate the upstream signal via feedback. In this study, we found that vinculin and FAK can be regulated backwards by F-actin assembly, increasing the uncertainty of molecular function. Therefore, more details of the molecular mechanism await further study.
Conclusions
Our results showed that TiO2 nanotubes promoted the osteogenic differentiation of MSCs, and this ability was enhanced with the increasing diameter of nanotubes within a certain range (30–70 V). F-actin mediated nanotube-induced cell differentiation through MKL1 and YAP/TAZ, providing a novel insight into the study of cell differentiation.
Доступность данных и материалов
Наборы данных, использованные и проанализированные в ходе текущего исследования, доступны у соответствующего автора по разумному запросу.
Сокращения
- MSCs:
-
Mesenchymal stem cells
- SEM:
-
Сканирующая электронная микроскопия
- EDS:
-
X-ray energy dispersive analysis
- AFM:
-
Атомно-силовая микроскопия
- ALP:
-
Alkaline phosphatase
- Cyto D:
-
Cytochalasin D
- Jasp:
-
Jasplakinolide
- VCL:
-
Vinculin
- FAK:
-
Focal adhesion kinase
- BCA:
-
Bicinchoninic acid
- PVDF:
-
Polyvinylidenedifluoride
- Runx2:
-
Runt-related transcription factor 2
- Osx:
-
Osterix
- OCN:
-
Osteocalcin
- YAP:
-
Yes-associated protein
- MKL1:
-
Megakaryoblastic leukemia 1
- FBS:
-
Фетальная бычья сыворотка
- ECM:
-
Extracellular matrix
- MLC:
-
Myosin light chain
- LIMK:
-
LIM kinase
- FH:
-
Formin-homology
- GPCR:
-
G protein-coupled receptors
- MAP2K:
-
Mitogen-activated protein kinase kinase
- ERK:
-
Extracellular signal-regulated kinase
- SRF:
-
Serum response factor
- SMA:
-
Smooth muscle actin
- PPARγ:
-
Peroxisome proliferator-activated receptor γ
- LATS1/2:
-
Large tumor suppressor gene 1 and 2
- TBX5:
-
T-box 5
Наноматериалы
- ДИОКСИД ТИТАНА - TiO2 - Цена, рынок и анализ
- Наножидкости TiO2 - Часть 1:Приготовление и свойства
- Нанотехнологии инактивируют раковые стволовые клетки
- На пути к наножидкости TiO2 - Часть 2:Приложения и проблемы
- Влияние распределения наночастиц золота в TiO2 на оптические и электрические характеристики сенсибилизирован…
- Синтез нанокристаллов ZnO и применение в инвертированных полимерных солнечных элементах
- Оптимальный предшественник титана для изготовления компактного слоя TiO2 для перовскитных солнечных элементо…
- Влияние упругой жесткости и поверхностной адгезии на отскок наночастиц
- Влияние наночастиц Ag разного размера и концентрации, внедренных в компактный слой TiO2, на эффективность преоб…
- Новый материал для преобразования с повышением частоты из трехдегированного TiO2, легированного Ho3 + -Yb3 + -Mg2 +, и …