


Полые наносферы кремнезема, инкапсулированные пероксидазой хрена для внутриклеточного определения активных форм кислорода
Аннотация
Активные формы кислорода (АФК) играют решающую роль в передаче сигналов и гомеостазе клеток. Избыточное производство АФК может вызывать окислительное повреждение различных биомолекул и клеточных структур. Таким образом, разработка подхода к мониторингу и количественной оценке АФК в живых клетках имеет важное значение для физиологии и клинической диагностики. Некоторые разработанные проницаемые для клеток флуорогенные зонды полезны для обнаружения ROS в сочетании с пероксидазой хрена (HRP). Однако их внутриклеточный сценарий затруднен из-за непроницаемости для мембран ферментов. В данном документе с помощью метода микроэмульсии был разработан новый подход к внутриклеточному восприятию АФК с использованием инкапсулированных пероксидазой хрена полых наносфер кремния (обозначенных как HRP @ HSN) с удовлетворительной каталитической активностью, проницаемостью клеточной мембраны и биосовместимостью.
Эти HRP @ HSN в сочетании с селективными зондами или нацеливающими лигандами могут быть предусмотрены в качестве инструментов для обнаружения АФК в определенных органеллах или типах клеток. Таким образом, HRP @ HSN, связанные с дигидрородамином 123, использовали для качественного и полуколичественного анализа физиологического H 2 О 2 уровни в активированных макрофагах RAW 264.7. Мы предполагаем, что эти HSN, инкапсулирующие активные ферменты, можно конъюгировать с селективными зондами и нацеливающими лигандами для обнаружения ROS в конкретных органеллах или интересующих типах клеток.
Фон
Активные формы кислорода (АФК), состоящие из радикальных и нерадикальных молекул, таких как супероксид-анионы, перекись водорода, гидроксильный радикал, синглетный кислород и пероксинитрит, непрерывно образуются во время аэробного метаболизма. Клеточные АФК в основном образуются из митохондриальной цепи переноса электронов (mETC) и обычно уравновешиваются ферментативными (такими как супероксиддисмутазы, каталазы и пероксидазы) и неферментативными (например, витаминами A, C и E; уратами и билирубином) ) антиоксидантная защита [1]. Однако дисбаланс в производстве АФК может привести к окислительному стрессу и последующему повреждению ДНК, жирных кислот, белков и других клеточных компонентов, потенциально способствуя развитию диабета [2], рака [3], сердечно-сосудистых заболеваний [4] и нейродегенеративных расстройств. [5], такие как болезнь Альцгеймера и болезнь Паркинсона. Прямая визуализация и количественное определение АФК в живых клетках очень желательно, но очень сложно.
Достижения в флуоресцентной микроскопии [6, 7] позволили разработать неинвазивные методы измерения и визуализации эволюции АФК на уровне отдельных клеток. Для обнаружения АФК большинство зондов предназначены для измерения изменений интенсивности флуоресценции или сдвигов в длине волны излучения (т.е. ратиометрические методы) после окисления профлуоресцентных ароматических молекул или снятия защиты с замаскированных соединений до флуоресцентных продуктов [8]. Специфичность к определенному типу ROS важна при разработке успешных зондов; например, окисление бороната используется как подход биоортогональной реакции для изучения химии перекиси водорода в живых системах [9]. Чтобы исследовать пространственно-временную динамику ROS, были созданы несколько зондов на основе бороната, конъюгированных с положительно заряженным фосфониевым фрагментом, для нацеливания на митохондрии [10, 11]. Однако их потенциал для визуализации in vivo ограничен их нестабильностью в биологической среде, низким проникновением через тканевые барьеры и быстрым выведением из организма через мочевыделительную систему [12,13,14]. Чтобы преодолеть такие проблемы, были разработаны некоторые стратегии путем химической прививки дополнительной стабилизирующей структуры к зонду [15] (например, цепи триэтиленгликоля), разработки генетически кодируемых индикаторов на основе флуоресцентных белков [16] или применения основанных на реакциях биолюминесцентные репортеры [17] или зонды для позитронно-эмиссионной томографии (ПЭТ) для молекулярной визуализации АФК [18]. Более того, несколько всеобъемлющих исследований подчеркнули, что наночастицы являются важным аспектом дизайна, и продемонстрировали, что зонды на основе наночастиц могут обеспечить понимание механизмов и новаторские стратегии для визуализации ROS в живых организмах с высокой специфичностью и чувствительностью [19,20,21,22]. Ферменты с высокой каталитической активностью и особой селективностью к субстрату также использовались в качестве инструментов клинической диагностики для идентификации целевых аналитов. Однако отсутствие длительной стабильности и трудности с проникновением свободных ферментов через биологические мембраны часто ограничивают их применение в сложной биологической среде. Несмотря на то, что применение электрода не подходит для внутриклеточных анализов или визуализации in vivo, значительные усилия были направлены на разработку биосенсоров, содержащих пероксидазу хрена (HRP), для определения H 2 О 2 на основе электрохимических методов [23, 24].
В этой работе ферментные нанореакторы, состоящие из HRP, инкапсулированного в полые наносферы из диоксида кремния размером 45 нм, были синтезированы методом микроэмульсии вода-в-масле (w / o) с последующим мягким процессом травления [25]. Ранее мы продемонстрировали, что такие полые наноматериалы могут поддерживать стабильную активность инкапсулированных ферментов и нанокатализаторов, защищая от протеолиза и спекания соответственно [26, 27]. В этой работе мы оценили их потенциальное использование в качестве внутриклеточных биосенсоров, изучая эффективность захвата ферментов, нагрузочную способность, реактивность и селективность пероксида, клеточное поглощение, токсичность и эффекты пролиферации HRP @ HSN. Используя дигидрородамин 123 (DHR123) в качестве субстрата, который обычно связывают с HRP для обнаружения внутриклеточного образования пероксида водорода, взаимодействия между HRP @ HSN и различными типами ROS в водных растворах исследовали с помощью проточной цитометрии и флуоресцентной микроскопии. Кроме того, было продемонстрировано, что использование HRP @ HSN с DHR123 может одновременно отображать и количественно определять физиологический H 2 О 2 уровни в макрофагах RAW264.7, стимулированных форбол-12-миристат-13-ацетатом (PMA). Взятые вместе, ферментные нанореакторы HRP @ HSN обладают потенциалом для визуализации воспалительных клеток, ассоциированных с АФК, in vivo, а инкапсулированные компоненты могут быть расширены до нескольких различных ферментов [28], наночастиц [26] и узнающих молекул для синергетических применений.
Методы / экспериментальные
Химические вещества и реагенты
Декан, n -гексанол (98%), гидроксид аммония (NH 4 OH, 35 мас.%), Тетраэтилортосиликат (TEOS, 98%), 3-аминопропилтриметоксисилан (APTMS, 95%) и изомер флуоресцеинизотиоцианата (FITC) были приобретены у ACROS. Полиоксиэтилен (5) изооктилфениловый эфир (Igepal CA-520), HRP типа VI-A (HRP), 3,3'5,5'-тетраметилбензидин (TMB), лимонная кислота, диметилсульфоксид (DMSO) и изотиоцианат родамина B ( RITC) были приобретены у Sigma-Aldrich. 2- (4-Йодофенил) -3- (4-нитрофенил) -5- (2,4-дисульфофенил) -2H-тетразолий был приобретен у Clontech. DHR123 и PMA были приобретены у Cayman Chemical. Перекись водорода (H 2 О 2 , 35%) была приобретена у SHOWA Chemical Industry. Раствор трет-бутилгидропероксида (70% в H 2 O) был приобретен у Aldrich. Перхлорат железа (II) (Fe (ClO 4 ) 2 ) был куплен у Альфа Азар. Сверхчистая деионизированная (D.I.) вода была получена с помощью системы Millipore Milli-Q Plus. Все реагенты использовали без дополнительной очистки.
Синтез полых кремнеземных наносфер (HSN)
HSN были синтезированы с помощью системы обратной микроэмульсии, сопровождаемой методом селективного травления, как описано в наших предыдущих исследованиях [25, 29]. Обычно 20 мл декана в качестве масляной фазы, 1,63 мл CA-520 в качестве поверхностно-активного вещества и 550 мкл n -гексанол в качестве вспомогательного поверхностно-активного вещества смешивали, а также перемешивали магнитной мешалкой с 2-сантиметровой мешалкой с покрытием из ПТФЭ при 650 об / мин. После этого 350 мкл D.I. к смеси добавляли воду при комнатной температуре, получая систему микроэмульсии вода-в-масле (w / o). Затем при перемешивании добавляли 25 мкл этанольного раствора APTMS (200 мкл APTMS в 1,4 мл абсолютного этанола) и 100 мкл TEOS. После перемешивания в течение 10 мин в систему вводили 250 мкл водного раствора аммиака (35 мас.%) При перемешивании при 20 ° C. Через 10 ч добавляли 95% этанол для дестабилизации системы микроэмульсии и твердые наночастицы диоксида кремния (SSN) собирали центрифугированием при 11000 об / мин в течение 20 минут. Для получения HSN, SSN были приостановлены в D.I. воду при перемешивании при 40 ° С в течение 40 мин. Затем HSN собирали центрифугированием при 11000 об / мин в течение 20 минут и несколько раз промывали 95% этанолом. Наконец, HSN суспендировали и хранили в 99,5% этаноле.
Синтез полых наносфер кремния, инкапсулированных пероксидазой хрена (HRP @ HSN)
HRP @ HSN были синтезированы методом, основанным на наших предыдущих исследованиях [27, 28]. Обычно синтез аналогичен описанной выше процедуре, за исключением того, что 350 мкл D.I. воду заменяли 350 мкл водной HRP (90 мкл 10 мг / мл раствора HRP в 350 мкл D.I. воды). После синтеза HRP @ HSN хранили в D.I. вода при 4 ° С.
Синтез FITC-HSN и HRP @ FITC-HSN
HSN и HRP @ HSN со встроенным флуоресцентным красителем, излучающим зеленый свет (обозначенные как FITC-HSN и HRP @ FITC-HSN), были синтезированы аналогично описанной выше процедуре, за исключением того, что раствор APTMS в этаноле был заменен на раствор FITC-APTMS. Этанольный раствор FITC-APTMS был приготовлен путем смешивания 10 мг FITC и 200 мкл APTMS с 1,4 мл абсолютного этанола в темноте в течение 18 часов при комнатной температуре.
Эффективность захвата HRP и грузоподъемность HRP @ HSN
Сначала смесь, содержащую HRP (6 мг в 500 мкл воды D.I.) и RITC (3 мг в 350 мкл ДМСО), перемешивали в темноте в течение 24 часов при 4 ° C. После этого смесь переносили на диализную мембрану, состоящую из регенерированной целлюлозы с отсечкой по молекулярной массе 12 ~ 14 кДа. Затем, чтобы удалить непрореагировавший RITC, диализный мешок подвергали диализу против 1 л D.I. воды и осторожно перемешивают 3 дня. Наконец, для синтеза RITC-HRP @ HSN использовали меченную RITC HRP (обозначенную как RITC-HRP).
Чтобы определить нагрузочную способность HRP, RITC-HRP @ HSN растворяли в 1 мл NaOH (1 M) в течение 1 часа, и количество захваченного RITC-HRP рассчитывали по калибровочной кривой, построенной путем построения графика зависимости интенсивности флуоресценции от концентрация RITC-HRP. Флуоресценцию измеряли с помощью прибора Hitachi F-4500 при длине волны возбуждения 543 нм и длине волны излучения 550 ~ 650 нм. Эффективность улавливания HRP и нагрузочная способность HRP @ HSN определялись следующим образом:эффективность улавливания (%) =масса RITC-HRP в RITC-HRP @ HSN / начальная масса RITC-HRP; и грузоподъемность =масса RITC-HRP в HRP-RITC @ HSN / масса RITC-HRP @ HSN.
Анализ активности HRP
Для определения активности фермента пероксидазы использовали хромогенный субстрат TMB. TMB может превращаться в окрашенный продукт при окислении HRP с использованием перекиси водорода в качестве окислителя. Сначала готовили различные концентрации нативной HRP и HRP @ HSN в фосфатном и цитратном буфере (pH 5,2). Затем в каждый раствор добавляли 50 мкл раствора TMB (20 мкМ в ДМСО) и 50 мкл H 2 . О 2 (20 мкМ в D.I. воде). За реакцией следили, измеряя оптическую плотность при 655 нм с использованием считывающего устройства для микропланшетов (BioTek Synergy Hybrid Reader). Активность HRP, инкапсулированного в HSN, рассчитывалась по калибровочной кривой нативной HRP.
Анализ реактивности HRP @ HSN с различными ROS
DHR123 (20 мкМ) отдельно или в смеси с HRP @ HSN (50 мкг / мл) инкубировали с различными типами ROS (100 мкМ) в 100 мкл раствора DMEM (pH 7,4). Эмиссию флуоресценции при 530 нм (λex =488 нм) контролировали каждые 5 минут в течение первых 120 минут. Исследованные АФК были получены следующим образом:пероксид водорода (H 2 О 2 ) и трет-бутилгидропероксид (TBHP) получали из коммерчески доступных 32 и 70% водных растворов соответственно. Супероксид (O 2 • - ) был получен из 10 мМ исходного раствора супероксида калия (KO 2 ) в DMEM. Гидроксильные радикалы (• OH) и трет-бутокси-радикалы (• OtBu) были получены реакцией 1 мМ Fe (ClO 4 ) 2 с 100 мкМ H 2 О 2 или 100 мкМ TBHP соответственно.
Анализ клеточной культуры и жизнеспособности
Клеточная линия макрофагов мыши RAW264.7 была получена от ATCC. Клетки RAW264.7 поддерживали в DMEM с 10% FBS, 100 Ед / мл пенициллина и 100 мкг / мл стрептомицина (Gibco) при 37 ° C в 5% CO 2 Атмосфера. Обычно 2 × 10 5 Клетки RAW264.7 на лунку высевали в 24-луночные планшеты для анализа жизнеспособности. Через 24 ч клетки дважды промывали PBS и инкубировали с различными количествами (0, 50, 100 и 200 мкг / мл) суспензии наночастиц в бессывороточной среде DMEM в течение 2 часов. Для анализа цитотоксичности клетки, обработанные наночастицами, дважды промывали культуральной средой с последующей инкубацией с реагентом WST-1 (Clontech) при 37 ° C в течение 2 часов. Для анализа пролиферации клеткам после обработки наночастицами в течение 2 часов позволяли расти в обычной питательной среде в течение 24 часов с последующей инкубацией с реагентом WST-1. Жизнеспособность клеток определяли с помощью красителя формазана, генерируемого живыми клетками, и измеряли оптическую плотность при 450 нм с эталонной длиной волны 650 нм с использованием считывающего устройства для микропланшетов (Bio-Rad, модель 680).
Анализ поглощения ячеек
RAW264,7 ячеек при 1 × 10 6 на лунку высевали в шестилуночные планшеты в течение ночи. Затем макрофаги RAW264.7 обрабатывали различными количествами (0, 50, 100 и 200 мкг / мл) суспензии наночастиц в бессывороточной среде DMEM в течение 2 часов. После этого клетки трижды промывали PBS и отделяли раствором трипсин-ЭДТА. Поглощение наночастиц макрофагами RAW264.7 исследовали методом проточной цитометрии. Трипановый синий использовался для гашения флуоресценции наночастиц, адсорбированных на внешней мембране клеток.
Анализ проточной цитометрии продукции ROS в макрофагах RAW264.7, стимулированных PMA
Обычно после 2 часов обработки макрофагов RAW264.7 наночастицами клетки промывали трижды PBS с последующей инкубацией с 20 мкМ DHR123 в бессывороточной среде DMEM в течение 30 минут. Затем клетки RAW264.7 промывали PBS и инкубировали с культуральной средой, содержащей PMA в различных концентрациях, в течение 1 часа. После промывки макрофаги RAW264.7 собирали и анализировали с помощью проточного цитометра FACS Canto II.
Количественный анализ
RAW264,7 ячеек при 3 × 10 4 на лунку высевали в 96-луночные планшеты для полуколичественных анализов. После инкубации с 50 мкл 100 мкг / мл суспензии наночастиц в бессывороточной среде DMEM в течение 2 часов обработанные наночастицами клетки обрабатывали 50 мкл бессывороточной среды DMEM, содержащей различные концентрации PMA, и 20 мкМ DHR123 для дополнительной обработки. 1 ч при 37 ° C. В то же время внешние стандарты H 2 О 2 смешанные с 50 мкг / мл HRP @ HSN, использовали для построения калибровочной кривой путем построения графика зависимости интенсивности флуоресценции от концентрации H 2 О 2 . Интенсивность флуоресценции измеряли с помощью считывающего устройства для микропланшетов (BioTek Synergy Hybrid Reader) при возбуждении при 488 нм и испускании при 530 нм. Используя установленную калибровочную кривую, количество H 2 О 2 в клетках RAW264.7, стимулированных различными количествами ФМА.
Характеристика
Изображения на просвечивающем электронном микроскопе (ПЭМ) были получены на JEOL JEM-1200 EX II, работающем при 100 кВ. Изображения были записаны с помощью камеры GatanOrius CCD. Образцы диспергировали в 95% этаноле и капали на покрытую углеродом медную сетку, а затем сушили на воздухе и исследовали. Для проверки HRP в полых сферах образец с отрицательным окрашиванием перемешивали в 1% водном растворе уранилацетата (UA) в течение 1 ч, а затем центрифугировали для удаления оставшегося UA. Наконец, образец был диспергирован в этаноле и помещен на медную сетку для визуализации. Измерения динамического рассеяния света (DLS) и дзета-потенциала проводили на Zetasizer Nano ZS (Malvern, UK). Оптические изображения клеток RAW264.7 были получены с помощью инвертированного микроскопа Zeiss Axio Observer Z1.
Результаты и обсуждение
Разработка и синтез HSN и HRP @ HSN
Обычно HSN и HRP @ HSN синтезировали с помощью золь-гель процесса, катализируемого аммиаком, в сочетании с системой микроэмульсии вода-в-масле (w / o) в соответствии с нашим предыдущим методом [27, 28]. Схема 1 иллюстрирует синтез HRP @ HSN. Согласно изображениям ПЭМ (рис. 1), HSN с инкапсулированной HRP и без нее показали средний диаметр 45 нм (дополнительный файл 1:Рисунок S1). Окрашивание UA ясно показало повышенную электронную плотность внутри HRP @ HSN, но не наблюдалось окрашивания вне HRP @ HSN (рис. 1b), что указывает на то, что ферменты HRP были успешно захвачены во внутренней полости HRP @ HSN.
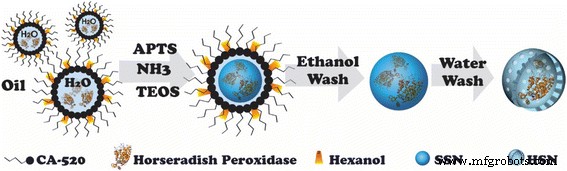
Блок-схема синтеза инкапсулированных пероксидазой хрена полых наносфер кремния (HRP @ HSNs). APTMS, 3-аминопропилтриметоксисилан; ТЭОС, тетраэтилортосиликат; SSN, твердые наночастицы диоксида кремния
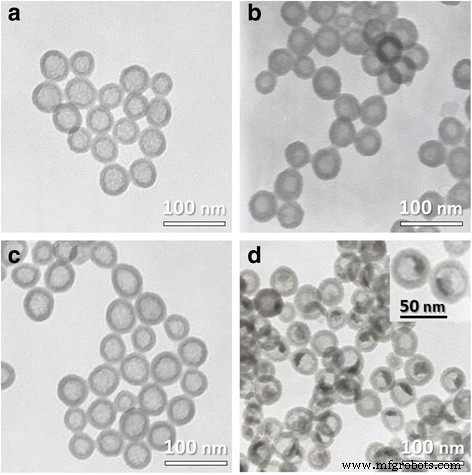
TEM-изображения a полые наносферы из диоксида кремния (HSN), b HSN, окрашенные уранилацетатом, c HSN, инкапсулированные пероксидазой хрена (HRP @ HSN), и d HRP @ HSN, окрашенные уранилацетатом. Врезка:увеличенный вид
Измерения DLS и анализы дзета-потенциала, выполненные при комнатной температуре, показаны в таблице 1. Данные DLS показали, что как HSN, так и HRP @ HSN давали положительные дзета-потенциалы в воде (pH ~ 6,5) с гидродинамическими диаметрами 188 ± 4 и 184 ± 6 нм. в воде соответственно. Однако, когда наночастицы были диспергированы в бессывороточной среде DMEM, гидродинамические диаметры увеличились до 1767 ± 94 нм для HSN и 1598 ± 127 нм для HRP @ HSN. Это указывает на небольшую степень агрегации HSN, но они все еще хорошо взвешены в среде. Между тем, отрицательные дзета-потенциалы обеих наночастиц, измеренные в среде, предполагали, что некоторые из ионов и биомолекул из биологической среды могли адсорбироваться на поверхности наночастиц [30, 31]. В этом случае положительно заряженные поверхности наночастиц были покрыты отрицательно заряженными веществами, которые быстро вызывали агрегацию наночастиц за счет электростатических взаимодействий. Для уменьшения неспецифической агрегации и повышения коллоидной стабильности наночастиц в биологические среды был введен бычий сывороточный альбумин (БСА) [28]. Впоследствии гидродинамические диаметры HSN и HRP @ HSN показали значительное уменьшение гидродинамических диаметров до 197 ± 43 и 195 ± 19 нм соответственно.
Эффективность захвата HRP и грузоподъемность HRP @ HSN
Чтобы исследовать эффективность и нагрузочную способность захвата HRP, была приготовлена HRP, меченная флуоресцентным красителем (RITC) (обозначенная как RITC-HRP). Интенсивность флуоресценции RITC-HRP @ HSN измеряли путем суспендирования наночастиц в 1 М NaOH, и количество инкапсулированного RITC-HRP определяли в соответствии с калибровочной кривой, построенной путем построения графика зависимости интенсивности флуоресценции от концентрации нативного RITC-HRP. при тех же условиях (дополнительный файл 1:рисунок S2). Чтобы изучить влияние концентрации фермента на эффективность захвата и загрузочную способность, в синтез вводили три различных количества HRP (11,1, 22,2 и 33,3 нмоль). Стоит отметить, что в этом диапазоне концентраций, независимо от того, сколько фермента было введено, эффективность улавливания ферментов для каждого из трех случаев составляла около 6%. Такая низкая эффективность может быть связана с тем, что только часть капель микроэмульсии образовала зародыши и выросла до HSN; большая часть капель микроэмульсии не зародилась и сохранила свой небольшой размер ~ 8 нм [25]. В будущем может потребоваться работа для повышения эффективности погрузки. Однако емкость загрузки HRP @ HSN постепенно увеличивалась до 12,5 ± 1,2 мкг HRP / мг HSN при использовании 33,3 нмоль HRP (дополнительный файл 1:таблица S1). Это указывает на то, что загрузочная способность HRP может контролироваться количеством фермента, присутствующего в реакции.
Цитотоксичность и клеточное поглощение HSN и HRP @ HSN
Чтобы оценить цитотоксичность HSN и HRP @ HSN in vitro, жизнеспособность клеток исследовали с помощью анализов WST-1. Как показано в Дополнительном файле 1:Рисунок S3, не наблюдалось значительного изменения пролиферации клеток RAW264.7 после обработки наночастицами в течение 2 часов или 2 часов с последующими дополнительными 24 часами культивирования. Не было обнаружено очевидного влияния наночастиц диоксида кремния на клеточную митохондриальную функцию в указанные моменты времени, независимо от присутствия или отсутствия HRP внутри HSN.
Затем были приготовлены FITC-конъюгированные HSN и HRP @ HSN, соответственно, для исследования влияния концентрации наночастиц на маркировку RAW264.7. Результаты проточной цитометрии (дополнительный файл 1:рисунок S4) показали, что клетки RAW264.7 были успешно помечены FITC-HSN и HRP @ FITC-HSN в различных концентрациях в течение 2 часов в бессывороточной среде. В обоих случаях было обнаружено дозозависимое повышение эффективности мечения, и более 80% клеток RAW264.7 были помечены воздействием наночастиц в концентрации> 50 мкг / мл в течение 2 часов. Такие свойства, как высокоэффективное внутриклеточное мечение с коротким временем инкубации, относительно низкая доза наночастиц и отсутствие цитотоксичности, делают HRP @ HSN подходящими для внутриклеточного обнаружения ROS.
Реакция HRP @ HSN на различные ROS
Согласно анализу активности фермента HRP с использованием TMB в качестве субстрата, около 40% исходной активности фермента оставалось после последующей инкапсуляции HRP в HSN. Это снижение наблюдаемой специфической активности инкапсулированного фермента (количество молей субстрата, преобразованных на единицу фермента за единицу времени) могло быть результатом ограничений массопереноса, которые возникают, когда субстраты пересекают кремнеземную оболочку в направлении HRP [32]. Тем не менее, стратегия инкапсуляции обеспечивает дополнительные функции, например, пористая оболочка из диоксида кремния может защищать HRP от протеолиза, позволяя транспортировать небольшие молекулы реагентов и продуктов [26, 27]. Взятые вместе, наблюдаемая реакционная способность HRP @ HSN к ROS, оцененная путем включения флуоресцентного зонда (DHR123), может быть результатом сочетания сродства наночастиц, а также внутреннего свойства HRP для ROS.
Бесклеточные системы использовали для генерации различных биологически релевантных АФК, включая перекись водорода (H 2 О 2 ), TBHP, гидроксильные радикалы (• OH), трет-бутокси-радикалы (• OtBu) и супероксид (O 2 • - ). Сначала DHR123 инкубировали с панелью ROS в отсутствие и в присутствии HRP или HRP @ HSN с последующим измерением интенсивности флуоресценции продукта родамина 123 (R123). Как показано на рис. 2, независимо от того, какой тип ROS использовался, интенсивность флуоресценции измерялась в зависимости от времени (30, 60, 90 и 120 минут). Однако очевидные различия в интенсивности между различными АФК зависят от внутренних свойств DHR123. С одной стороны, в соответствии с предыдущим исследованием [33], рис. 2а показывает, что ни H 2 О 2 ни O 2 • - может окислить DHR123 до R123. Кроме того, DHR123 проявил более высокую реакционную способность в отношении радикалов • OtBu и • OH по сравнению с другими АФК. При каталитической активности HRP заметное увеличение интенсивности флуоресценции наблюдалось в присутствии нативных HRP и HRP @ HSN, как показано на фиг. 2b, c. Было отмечено, что более высокая интенсивность флуоресценции, обнаруженная в случае нативной HRP по сравнению с HRP @ HSN при том же времени реакции, положительно коррелировала с наблюдаемой ими ферментативной активностью.
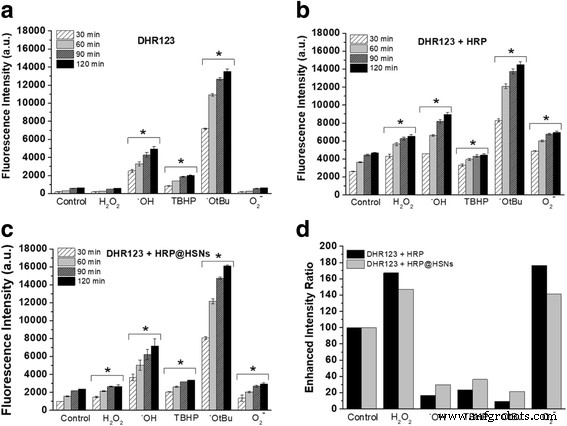
а - c Зависящая от времени интенсивность флуоресценции реакции выбранных активных форм кислорода (АФК) с a дигидрородамин 123 (DHR123), b DHR123 + пероксидаза хрена (HRP) и c DHR123 + HSN, инкапсулированные пероксидазой хрена (HRP @ HSN). г Повышенное соотношение интенсивностей реакции выбранных АФК с DHR123 + HRP и DHR123 + HRP @ HSN через 1 час. Показанные данные относятся к 20 мкМ DHR123, 400 нг / мл HRP, 50 мкг / мл HRP @ HSN и 100 мкМ ROS. (* p <0,05 по сравнению с контрольной группой в соответствующие моменты времени)
Чтобы позволить прямое сравнение между различными ROS, данные с интервалом времени 60 минут были выбраны и представлены как относительная интенсивность флуоресценции, нормализованная к контролю (дополнительный файл 1:рисунок S5). Последующий анализ отношения повышенной интенсивности был продемонстрирован путем деления относительной интенсивности флуоресценции DHR123 + HRP или DHR123 + HRP @ HSN на DHR123 (фиг. 2d). В обоих случаях, содержащих HRP, наблюдается аналогичная тенденция увеличения отношения интенсивностей для различных ROS, а также значительное увеличение реакционной способности DHR123 по отношению к H 2 О 2 и O 2 • - наблюдались, демонстрируя, что инкапсулированный HRP дает высокую степень внутренней ферментативной активности, а кремнеземные оболочки HRP @ HSN позволяют транспортировать небольшие молекулы для проведения селективного биокатализа.
Обнаружение внутриклеточных ROS с помощью HRP @ HSN
Чтобы оценить функциональность обнаружения АФК HRP @ HSN внутри клеток, макрофаги RAW264.7 инкубировали с HRP @ HSN в течение 2 часов с последующей промывкой и затем инкубированием с DHR123 (20 мкМ) в течение 30 минут. Затем клетки промывали и обрабатывали PMA (1 мкг / мл) в течение еще 1 часа. Известно, что стимуляция макрофагов ФМА приводит к образованию супероксида, который превращается в перекись водорода с помощью супероксиддисмутазы или спонтанной дисмутации [34,35,36]. Таким образом, PMA может действовать как стимулятор для выработки H 2 О 2 в макрофагах RAW264.7 для оценки внутриклеточного H 2 О 2 -сенсорная способность HRP @ HSN. Как показано на рис. 3a, оба случая макрофагов RAW264.7, культивированных отдельно и культивированных с HSN, показали слабую флуоресценцию в анализе проточной цитометрии, что указывает на то, что нестимулированные клетки продуцировали слабый базальный уровень ROS, при этом значительная ROS не индуцировалась в наличие HSN. Кроме того, клетки, обработанные HRP @ HSN, показали значительное увеличение интенсивности (рис. 3a), предполагая, что доставленные HRP @ HSN обеспечивали дополнительную каталитическую активность внутри клеток.
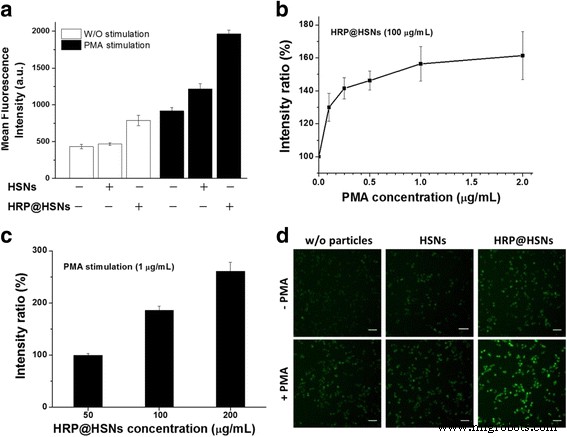
а Анализ проточной цитометрии макрофагов RAW264.7, стимулированных форбол-12-миристат-13-ацетатом (PMA) и без него, в присутствии и в отсутствие наночастиц. б PMA и c Концентрация HSN, инкапсулированных пероксидазой хрена (HRP @ HSN), зависимо изменяла флуоресценцию макрофагов RAW264.7. г Репрезентативные флуоресцентные изображения макрофагов RAW264.7 в указанных условиях. Шкала 50 мкм
Для экспериментов по стимуляции клетки, обработанные PMA, обычно генерировали более чем в два раза более высокие уровни флуоресценции R123 по сравнению с нестимулированными клетками. Кроме того, клетки, обработанные HRP @ HSN, имели наивысший уровень флуоресценции, за ними следовали HSN, а затем только клетки. Было отмечено, что обработка стимулированных макрофагов RAW264.7 HSN приводила к небольшому увеличению интенсивности флуоресценции по сравнению с контролем. Этот результат свидетельствует о том, что клеточные реакции на стресс запускаются чрезвычайно быстро и чувствительны к внешним раздражителям, включая воздействие наночастиц [37]. Кроме того, как PMA (0,1, 0,25, 0,5, 1 и 2 мкг / мл), так и HRP @ HSN (50, 100 и 200 мкг / мл) индуцировали экспрессию R123 в зависимости от дозы, как видно на рис. . 3б, в.
В соответствии с данными проточного цитометрического анализа на рис. 3d представлены типичные флуоресцентные изображения макрофагов RAW264.7, стимулированных с помощью и без PMA в присутствии и в отсутствие наночастиц. Система была способна визуализировать эндогенный H 2 О 2 генерации в клетках RAW264.7, а самая слабая интенсивность флуоресценции наблюдалась в клетках, обработанных HRP @ HSN с последующей стимуляцией PMA. Как показано на фиг. 4a, жизнеспособность клеток макрофагов RAW264.7 в присутствии стимулятора PMA или экзогенного H 2 О 2 был исследован тестами WST-1. В то время как АФК участвуют в апоптозе [38], в указанный момент времени было обнаружено лишь небольшое влияние на жизнеспособность клеток, что делает следующий полуколичественный анализ практическим и значимым.
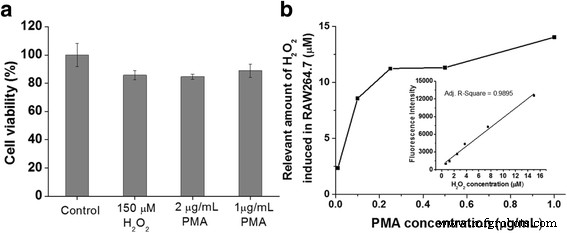
а Анализ WST-1 макрофагов RAW 264.7 после обработки экзогенным H 2 О 2 или стимуляция форбол 12-миристат 13-ацетат (PMA) в течение 1 часа. б Определение концентрации H 2 О 2 эндогенно продуцируется макрофагами RAW264.7 при различных концентрациях стимулятора PMA в присутствии инкапсулированных пероксидазой хрена полых наносфер кремния (HRP @ HSN) и дигидрородамина 123 (DHR123). Inset:a calibration curve obtained from the external standards of H2 О 2 mixed with HRP@HSNs and DHR123
Application of HRP@HSNs In Vitro for Quantitative Analysis of H2 О 2
To evaluate the capacity of HRP@HSNs for quantifying endogenous hydrogen peroxide produced in PMA-stimulated RAW264.7 cells, a calibration curve from the exogenous H2 О 2 experiment, with a detection range of 0.625~15 μM, was established by microplate measurements (Fig. 4b, inset). The standard calibration curve appears to be linear as expected. Then, RAW264.7 cells were treated with 100 μg/mL of HRP@HSNs for 2 h, followed by co-incubation with various concentrations of PMA and 20 μM of DHR123 at 37 °C for 1 h. After that, the concentration of H2 О 2 endogenously produced by PMA-stimulated RAW264.7 cells was determined by measuring the fluorescence intensity, followed by conversion using the established calibration curve. Notably, because most of the HRP@HSNs were uptaken within the cells, the H2 О 2 -triggered fluorescence of R123 could be attributed to intracellular enzyme-catalyzed reactions rather than the extracellular contribution. Although H2 О 2 is able to diffuse across biomembranes, due to its limited diffusion and rapid enzymatic consumption inside cells, concentration gradients of H2 О 2 are formed across membranes [39, 40]. Typically, under normal physiological conditions, H2 О 2 has an extracellular concentration estimated at 10 − 7 ~10 − 6 M, which is about 10-fold higher than that observed in intracellular fluid [1, 41, 42]. In pathological conditions, extracellular concentrations of H2 О 2 are in the range of 10~50 μM and are additionally elevated to as high as 10 − 4 M in apoptosis [1]. As shown in Fig. 4b and Additional file 1:Table S2, endogenous hydrogen peroxide caused by PMA-stimulated RAW264.7 cells was created in a dose-dependent manner and produced at levels of about 10 μM when the concentration of PMA used exceeded 0.25 μg/mL. Taken together, these results indicate that HRP@HSNs were capable of detecting semi-quantitatively endogenous the concentration of hydrogen peroxide of RAW264.7 macrophages under oxidative stress conditions.
Conclusions
In summary, we have demonstrated that hollow silica nanospheres encapsulating HRP can be synthesized via a microemulsion-templating system and act as intracellular fluorescent ROS sensors. The shells of HRP@HSNs are permeable to small molecules, such as the enzyme substrates, which allows them to react with large enzyme payloads in the hollow cavity. Both the effective intracellular delivery and satisfactory catalytic activity of HRP@HSNs significantly enhance reduction-triggered fluorescence and constitute the ability of semi-quantitative measurements of endogenous H2 О 2 in RAW264.7 macrophages under oxidative stress conditions.
Because the concentration and location of H2 О 2 in eukaryotic cells strongly rely on the types of cells, and cellular compartments [1], specific targeting of tumor cells or organelles could further be achieved by surface modification of HRP@HSNs with monoclonal antibodies or peptides. Also, non-enzymatic H2 О 2 detection could be realized by replacing the interior nanoreactors of HRP with nanoparticles [43, 44] or boronate-based fluorescent probes [42, 45]. Future efforts should be devoted to maximizing the sensitivity and specificity for H2 О 2 as well as enabling more informative designs of next-generation nanomaterials. Such hollow capsules could be a promising platform for modern nanomedicines that aims to simultaneously image, sensing, and deliver therapeutic molecules specifically to defective cells.
Сокращения
- APTMS:
-
3-Aminopropyltrimethoxysilane
- BSA:
-
Bovine serum albumin
- DHR123:
-
Dihydrorhodamine 123
- DLS:
-
Динамическое рассеяние света
- FITC:
-
Fluorescein isothiocyanate
- HRP:
-
Horseradish peroxidase
- HSNs:
-
Hollow silica nanospheres
- Igepal CA-520:
-
Polyoxyethylene (5) isooctylphenyl ether
- mETC:
-
Mitochondrial electron transport chain
- PET:
-
Positron emission tomography
- PMA:
-
Phorbol 12-myristate 13-acetate
- R123:
-
Rhodamine 123
- RITC:
-
Rhodamine B isothiocyanate
- ROS:
-
Reactive oxygen species
- SSN:
-
Solid silica nanoparticles
- TBHP:
-
Tert-butyl hydroperoxide
- ТЕМ:
-
Transmission electron microscopic
- TEOS:
-
Tetraethyl orthosilicate
- TMB:
-
3,3′5,5′-Tetramethylbenzidine
Наноматериалы
- Образец высоко реактивной треугольной молекулы IBM и Warwick впервые
- Настройка токсичности активных форм кислорода для расширенной терапии опухолей
- Электроспряденные полимерные нановолокна, украшенные наночастицами благородных металлов для определения х…
- Монодисперсные углеродные наносферы с иерархической пористой структурой в качестве материала электродов дл…
- Наночастицы кремнезема для доставки внутриклеточного белка:новый подход к синтезу с использованием зеленог…
- Иерархическая гетероструктура полых сфер ZnO @ TiO2 для высокоэффективного фотокаталитического выделения водор…
- Нанопетали из мезопористого оксида никеля (NiO) для сверхчувствительного определения уровня глюкозы
- Генерация активных форм кислорода в водных растворах, содержащих наночастицы GdVO4:Eu3 + и их комплексы с метилен…
- Оксид кремния, покрытый титаном, модифицированный альгинатом натрия в качестве сорбентов для ионов тяжелых м…
- Усиленный эффект объединения хлорогеновой кислоты на наночастицы селена в ингибировании агрегации β-амилои…