


Идентификация характерных макромолекул генотипов Escherichia coli с помощью наномасштабного механического картирования атомно-силового микроскопа
Аннотация
Классификация микробных штаммов обычно основана на молекулярном методе и редко является морфологическими характеристиками изучаемых бактериальных штаммов. В этом исследовании мы выявили макромолекулярные структуры бактериальной поверхности с помощью механического картирования АСМ, разрешение которого определялось не только размером наконечника в наномасштабе, но и механическими свойствами образца. Этот метод позволил провести наноразмерное исследование мембранных структур микробных штаммов с простой подготовкой образцов и гибкой рабочей средой, что позволило преодолеть многочисленные ограничения электронной микроскопии и биохимических аналитических методов, позволяющих использовать метки. Характерные макромолекулы, расположенные на клеточной поверхности, считались белками поверхностного слоя и оказались специфичными для Escherichia coli генотипы, у которых средний размер молекул характеризовался диаметрами в диапазоне от 38 до 66 нм, а форма молекул была похожа на почки или круглую. В заключение, поверхностные макромолекулярные структуры обладают уникальными характеристиками, которые связаны с E. coli генотип, который предполагает, что влияние генома на морфологию клеток можно быстро идентифицировать с помощью механического картирования АСМ.
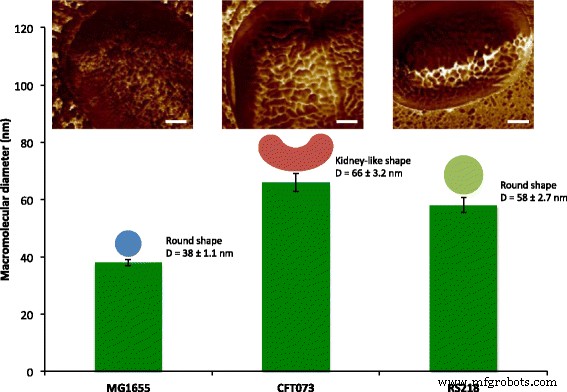
Количественное определение поверхностных макромолекул E. coli клетки с использованием механического картирования АСМ. Поверхностные макромолекулы клеточной поверхности трех E. coli генотипы MG1655, CFT073 и RS218 были охарактеризованы с размерами в диапазоне от 38 до 66 нм и с круглой или похожей на почки формой. Изображения топографии были окрашены с помощью карты адгезии с масштабными полосами =200 нм.
Фон
Из-за низкого разрешения оптической микроскопии и ограниченной рабочей среды электронной микроскопии исследователи в области микробов редко рассматривают возможность использования внешнего вида бактериальных клеток, а вместо этого применяют молекулярные или химические аналитические методы для идентификации геномных фрагментов, экспрессии белков и т. Д. Неудивительно, что эти методы имеют ряд недостатков, в том числе трудоемкость и длительность, поэтому необходимы более простые, эффективные и гибкие подходы. Изобретенный в 1982 году Биннигом и др., Атомно-силовой микроскоп (АСМ) предназначен для использования наноразмерного зонда, контролируемого лазерным лучом, для наблюдения за поверхностью образца с наноскопическим или даже атомным разрешением [1]. Получая изображение морфологии с помощью наконечника физического зонда, этот метод преодолевает предел разрешения и ограничения окружающей среды как оптической, так и электронной микроскопии и имеет ряд преимуществ, таких как простая подготовка образца и гибкие рабочие условия в условиях окружающего воздуха или жидкости [2, 3]. Тем не менее, что касается микробных исследований, текущие применения АСМ в основном включают качественное отображение статической или динамической морфологии бактериальных клеток или экспрессии жгутиков и пилей [4,5,6], в то время как мало исследований было сосредоточено на ультраструктуре поверхности микробные клетки и количественный анализ клеточных свойств.
В этом исследовании AFM был выбран для изучения поверхности Escherichia coli . ( E. coli ) клеток, а формы и размеры отдельных бактерий наблюдались с помощью топографии и фазовых изображений АСМ. Кроме того, было обнаружено, что одновременное механическое картирование выявило дополнительную биомеханическую информацию о компонентах поверхности, где небольшие различия в адгезионных характеристиках между макромолекулами и окружающей матрицей можно было обнаружить во время каждого физического контакта между зондом и образцом. Применяя такие передовые методы к микробным полям, мы исследовали три E. coli генотипы, содержащие один лабораторный штамм и два патогенных штамма человека для идентификации поверхностных макромолекул. Результаты показали, что этот метод может обеспечить разрешение изображения, превышающее масштаб наконечника АСМ, за счет определения механического распределения образца. В заключение мы предполагаем, что такое развитие науки о поверхности не только предоставит исследователям, работающим в области микробов, подробную информацию о внешнем виде клеток, но также внесет вклад в наши знания о характеристиках поверхности других систем био- или наноматериалов.
Методы
Образцы микробов
Три Э. coli штаммы, протестированные в этом исследовании, были клинически изолированы и предоставлены лабораторией профессора Чинг-Хао Тэна в Институте молекулярной медицины Национального университета Ченг Кунг. MG1655 представляет собой кишечный и лабораторный штамм дикого типа E. coli K-12 и два других штамма являются патогенами человека:CFT073, основная причина инфекций мочевыводящих путей, и RS218, ассоциированный с неонатальным менингитом у младенцев [7].
Функционализированный субстрат
Были применены две стадии модификации поверхности для ковалентного связывания между твердой поверхностью и микробными клетками. Сначала раствор 3-аминопропилтриэтоксисилана (APTES, Sigma-Aldrich Co. LLC, США) использовали для образования исходного NH 2 -функционализированный слой на поверхности, где стабильное покрытие APTES было внесено в стабильные связи Si-O - Si, обеспечиваемый APTES и O с окисленной поверхности. Обладая двумя функциями COOH, глутаральдегид соединяется с NH 2 функция APTES с одним COOH и работает с NH 2 на бактериальной поверхности с другим COOH.
Чистые подложки погружали в раствор APTES со смесью 5% APTES в этаноле на час и промывали этанолом и ddH 2 . О. Предметные стекла сушили потоком азота, а затем помещали в 2% раствор глутаральдегида в PBS на ночь и промывали PBS.
Подготовка образца
Единичные колонии E. coli Штаммы отбирали из агаровых чашек с бульоном для лизогени (LB) и инкубировали в бульоне LB. После 12 часов культивирования бактериальный раствор разбавляли 1:100 свежим предварительно уравновешенным бульоном LB. Еще через 12 ч культивирования микробов бактериальный раствор подвергали центрифугированию при 1500 × g (4000 об / мин) в течение 3 мин и ресуспендировали в бульоне LB, причем этот процесс повторяли дважды. Двести микролитров бактериального раствора добавляли по каплям на функционализированный субстрат и оставляли на 30 мин. Затем образец дважды погружали в дистиллированную воду, чтобы удалить неприкрепленные клетки, и сразу же получали изображение с помощью АСМ в окружающем воздухе.
Характеристика AFM
Для исследования поверхности микробных образцов были выбраны АСМ-прибор (Bruker Nano, Санта-Барбара, Калифорния, США) и зонд из нитрида кремния с калиброванной жесткостью пружины 0,7 Н / м и радиусом кончика 10 нм. Частота сканирования АСМ и линейные пиксели составляли 0,5 Гц и 256 строк, соответственно, для размера сканирования 10 мкм для первого обнаружения топографии, а затем параметры были установлены как 0,3 Гц и 512 строк для размера сканирования 2 мкм для подробное наблюдение. Количественный наномеханический (QNM) режим PeakForce использовался для наномеханического картирования, где адгезионные свойства поверхности рассчитывались по максимальной силе притяжения среди кривых «сила отрыва - расстояние».
Наше предыдущее исследование Streptococcus mutans показали механическую эволюцию на бактериальной поверхности в течение 2 часов, контролируемую непрерывным механическим картированием АСМ, и было подтверждено, что образцы микробов оставались живыми в течение этого времени [8,9,10]. Для обеспечения жизнеспособности E. coli образцы, использованные в этой работе, были проведены предварительные исследования бактериальных образцов, и непрерывное изменение поверхностной адгезии в течение 4 часов предполагало, что клетки остаются живыми в течение как минимум 4 часов после подготовки образца (показано в дополнительном файле 1). Следовательно, микробные образцы, протестированные в этой работе, были измерены с помощью АСМ в течение 2 часов после подготовки образца. Все E. coli генотипы культивировались индивидуально и в разное время, а измерения АСМ проводились сразу после подготовки образца. Другими словами, образцы бактерий не были выстроены в очередь в ожидании исследования, поэтому влияние времени выдержки на различия между E. coli напряжения были сведены к минимуму. Количественные данные для каждого E. coli штамм были собраны из измерений, проведенных в течение 2 часов в этом исследовании.
Статистический анализ
Для статистического анализа в данной работе использовалась Prism (GraphPad Software, США). Длина клеток и размеры макромолекул были представлены как средние значения вместе со стандартной ошибкой среднего (SEM). Множественные сравнения между E. coli Генотипы обрабатывали с использованием обычного одностороннего дисперсионного анализа (ANOVA). Уровень достоверности 95% ( p <0,05), а звездочки указывают степень обнаруженной значимой разницы. Номер образца n каждого штамма был> 40.
Результаты и обсуждение
Ультраструктура поверхности с помощью множественного сопоставления
При сканировании с помощью АСМ в масштабе наблюдения 10 мкм на бактериальном образце несколько одиночных E. coli Клетки MG1655 можно было увидеть, а их форму в трех измерениях можно было наблюдать на топографическом изображении (рис. 1а); Четкие контуры клеток, показанные в двух измерениях, были получены с помощью изображения ошибки отклонения (рис. 1b), и кроме бактериальных клеток можно было обнаружить несколько канальцев. Волнообразные филаменты (рис. 1c) соответствовали появлению микробных жгутиков, описанных в другой работе, которая подтвердила такую находку, как жгутики, а также можно было увидеть более короткие и похожие на волосы пили [11]. При уменьшении области наблюдения зонда для детального изучения отдельных микробных клеток топография показала небольшую разницу в вертикальном направлении между клеточными поверхностями, как показано на рис. 1d, а изображение ошибки отклонения, по-видимому, предоставило больше морфологической информации, в то время как собранного шума окружающей среды было слишком много для исследования ультраструктуры клеточной поверхности (рис. 1e). Когда одновременные биомеханические свойства образца были измерены во время контакта между наконечником и тестируемым объектом, было обнаружено, что бактериальная поверхность на самом деле состоит из огромного количества макромолекул определенной формы и размера, что было выявлено при картировании силы адгезии ( Рис. 1е); таким образом, морфологическое разрешение в топографических изображениях и изображениях ошибок отклонения было дополнительно улучшено за счет биомеханической информации.
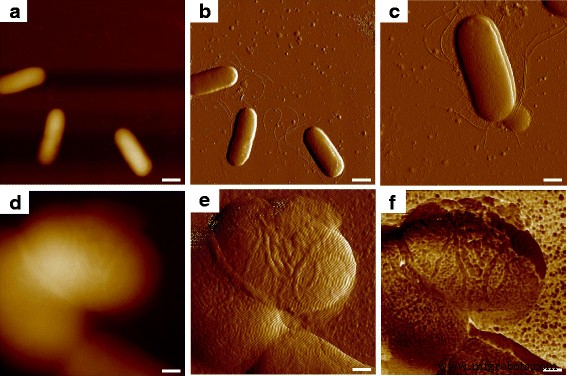
Ультраструктура поверхности E. coli MG1655 с использованием множественного отображения AFM. а и b были изображения топографической ошибки и ошибки прогиба, а c было детальное изображение ошибки отклонения бактериальной клетки с экспрессией жгутиков и пилей. Затем была сфокусирована одна ячейка, где d и е были изображения топографической ошибки и ошибки отклонения, а f было соответствующее отображение адгезии. Масштабные линейки =1 мкм в a и b , 500 нм дюйм c и 200 нм в d - е
На рис. 2а экспрессия жгутиков E. coli Видно было видно MG1655, где размер нитей был подобен размерам нитей на рис. 1b, с относительно более низкими адгезионными свойствами, чем у подложки. Вдобавок было обнаружено, что бактериальная поверхность состоит из круглых компонентов, которые характеризовались меньшей адгезией по сравнению с окружающей матрицей. Это наблюдение аналогично нашим предыдущим результатам о тканевых слоях кожи мышей, а повторяющиеся и сопоставимые зерна считались макромолекулами, чья структура более плотная и последовательная, чем структуры, наблюдаемые в межмолекулярной области, так что различия в характеристиках адгезии могут быть легко ощутимым [12]. Самый внешний слой клеточной оболочки у грамотрицательных бактерий - это слой белков самосборки, как показано на рис. 2b, который известен как белок поверхностного слоя (S-слой) [13]. Структуры S-слоя традиционно измерялись с помощью электронной микроскопии, в то время как требования к вакуумной среде и проводящему покрытию теряли естественную информацию о белках в реальном времени. Хотя некоторые исследователи извлекли белки S-слоя и повторно собрали их на слюдяной субстрат для сканирования АСМ, результатов не хватало для работы структуры S-слоя in situ и в реальном времени [14, 15]. Основываясь на клеточной архитектуре и некоторых предыдущих изображениях микробной поверхности с помощью электронной микроскопии, мы рассматривали наблюдаемые макромолекулы как белки S-слоя [16].
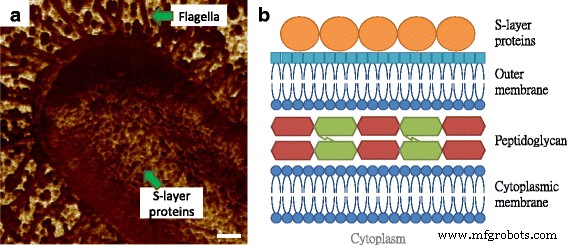
Иллюстрация вертикальных и поверхностных структур E. coli клетки. а Поверхность макромолекул на E. coli Клетка MG1655, полученная методом адгезионного картирования АСМ. б Молекулярная архитектура клеточной оболочки грамотрицательных микробов, которая состоит из цитоплазматической мембраны, пептидогликана, внешней мембраны и S-слоя. Масштабная линейка =200 нм
По сравнению с измерениями, проведенными с помощью АСМ и просвечивающей электронной микроскопии (ПЭМ), первый имеет несколько преимуществ по сравнению со вторыми, например, более простую подготовку образца, менее строгие экспериментальные требования и более безопасные для биологии приложения для получения изображений. Выбор ПЭМ в биологической области обычно выбирается из-за его способности видеть межклеточные органеллы и получения изображений сверхвысокого разрешения (обычно нанометрового или субнанометрового). Картирование адгезии АСМ в текущей работе улучшило разрешение элемента и представило расположение поверхностных макромолекул таким образом, который не зависел от размера иглы, а вместо этого от внутренней структуры самого образца. Кроме того, этот подход обеспечивает наноразмерное разрешение микробной поверхности в диапазоне квадратных десятков микрометров.
Геномно-манипуляционные различия в морфологических характеристиках
После наблюдения ультраструктуры поверхности E. coli Клетки MG1655, один интересный вопрос заключается в том, как устроены макромолекулы у других штаммов. Патогены человека E. coli Таким образом, CFT073 и RS218 были исследованы с помощью АСМ с одинаковыми экспериментальными параметрами, и не было значительных различий в клеточных формах и размерах между этими тремя генотипами при размере признаков 10 мкм, как показано на рис. 3a – c. Картирование силы адгезии использовалось для идентификации белков S-слоя, и среди разных E. coli деформации, как показано на рис. 3d – f. Для простоты сравнения детальные изображения карты адгезии E. coli деформации показаны на рис. 3g – i. Поверхностные макромолекулы были охарактеризованы как имеющие круглую форму в ячейках MG1655 и RS218, хотя их молекулярные диаметры различались, составляя 38 ± 1,1 нм ( n =80) для MG1655 и 58 ± 2,7 нм ( n =46) для RS218. С другой стороны, клетки CFT073 обладали уникальной формой белков S-слоя, которые были похожи на почки с разницей в длине между двумя конечными точками 66 ± 3,2 нм ( n =44). После анализа размеров белков S-слоя этих трех генотипов для множественных сравнений результаты продемонстрировали значительные различия между этими штаммами (рис. 4).
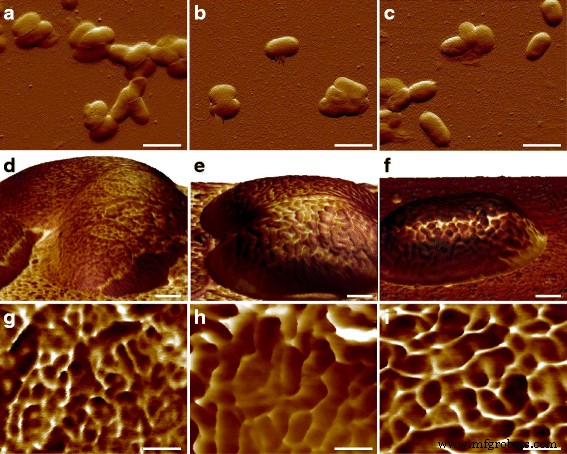
Морфологическая характеристика E. coli генотипы. В верхнем ряду отображены изображения ошибок отклонения a MG1655, b CFT073 и c RS218. В среднем ряду показаны трехмерные топографии, окрашенные картированием адгезии на d . MG1655, e CFT073 и f Ячейки RS218. Нижний ряд был подробным картированием адгезии на g . MG1655, h CFT073 и i RS218. При картировании адгезии более темный цвет означает меньшую адгезию и наоборот. Масштабные линейки составляли 2 мкм для a - c , 200 нм для d - е и 100 нм для g - я
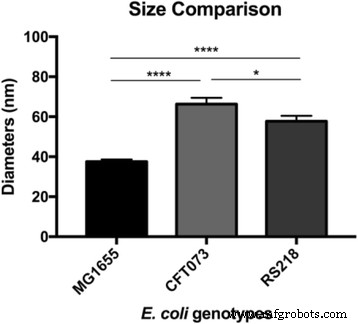
Размеры молекул E. coli генотипы. Диаметр поверхностных белков определяли с помощью АСМ и обрабатывали с помощью однофакторного дисперсионного анализа для множественных сравнений. **** p <0,001 и * p <0,05
Сообщается, что микробные S-слои играют важную роль во многих функциях, включая защиту клеток от неблагоприятных условий окружающей среды, атак фагоцитоза и хищных бактерий. Кроме того, S-слои также служат в качестве адгезина, который обеспечивает эффективную колонизацию [17]. Структуры S-слоя были хорошо изучены с помощью ПЭМ и были разделены на различные типы решеток с расстоянием между центрами в диапазоне 4–35 нм [16]. Разница между нашими результатами АСМ и отчетами ПЭМ из литературы рассматривалась как разные методологии визуализации, где ПЭМ дает двумерную морфологию структуры S-слоя, а АСМ фиксирует трехмерную топографию, которая включает в себя множественные влияния, вносимые радианом и шероховатостью клетки, а также геометрия зонда АСМ.
Изначально считалось, что различные типы структур S-слоя могут использовать их различные таксономические характеристики для различения видов бактерий, хотя затем было обнаружено, что даже для одного вида микробные красители могут иметь разные решетки белков [13, 16, 18, 19]. В то время как некоторые исследования изучали роль S-слоя в формировании филаментов, типы белков на клеточных мембранах и геномные различия по размеру среди E. coli генотипов различия в белках S-слоя отмечаются редко [20,21,22]. Результаты настоящего исследования выявили различия по морфологическим характеристикам между E. coli MG1655, CFT073 и RS218 и предполагают, что внешний вид поверхностных макромолекул, вероятно, был специфическим для отдельных E. coli генотип.
Выводы
В этой работе геномно-специфическая наноструктурная информация о бактериальной поверхности была обнаружена с помощью механического картирования АСМ, которое выявило адгезионные различия между макромолекулами и окружающей матрицей. В соответствии с молекулярной архитектурой грамотрицательных микробов поверхностные макромолекулы микробных клеток считались белками поверхностного слоя. Было обнаружено, что расположение и размеры этих макромолекул специфичны для тестируемого E. coli генотипы с различными формами и размерами, причем эти различия значимы статистическим анализом. В заключение мы считаем, что структура бактериального S-слоя является геном-зависимой и может быть потенциальным методом для быстрой диагностики заболеваний, связанных с микробами, или микробных штаммов. Для практического применения характеристики S-слоя требуется исследование большего количества бактериальных генотипов для каталога комплемента. В настоящее время мы создаем базу данных, объединяющую морфологические характеристики бактерий и физиологические / патологические характеристики, и полагаем, что это будет многообещающим прогрессом для практического применения АСМ исследования.
Сокращения
- AFM:
-
Атомно-силовой микроскоп
- ANOVA:
-
Односторонний дисперсионный анализ
- APTES:
-
3-аминопропилтриэтоксисилан
- LB:
-
Лизогенический бульон
- QNM:
-
Количественный наномеханический
- S-слой:
-
Поверхностный слой
- ТЕМ:
-
Просвечивающая электронная микроскопия
Наноматериалы
- Ученые IBM изобрели термометр для наномасштаба
- Пионеры AFM отмечены премией Кавли
- Следующий фонтан нефти находится в наномасштабе
- Непроводники проводят ток в наномасштабе
- Биогенный синтез, характеристика и оценка антибактериального потенциала наночастиц оксида меди против Escheric…
- Поверхностный эффект на транспортировку нефти в наноканале:исследование молекулярной динамики
- Малоугловое рассеяние от наноразмерных жирных фракталов
- Зависимость локализованного поверхностного плазмонного резонанса от смещенного усеченного димера наноприз…
- Высокопроизводительное производство качественных нановолокон с использованием модифицированного электрос…
- Изготовление наноразмерных ямок с высокой производительностью на полимерной тонкой пленке с использованием…