


Наночастицы, модифицированные hGC33 и содержащие сорафениб, обладают синергетическим эффектом против гепатомы, подавляя сигнальный путь Wnt
Аннотация
Доставка опухолеспецифических ингибиторов является сложной задачей при лечении рака. Модифицированные антителами наночастицы могут доставлять содержащиеся в них лекарства к опухолевым клеткам, которые сверхэкспрессируют специфические антигены, ассоциированные с опухолью. Здесь мы сконструировали нагруженные сорафенибом полимерные наночастицы полиэтиленгликоля-b-PLGA, модифицированные антителом hGC33 к глипикану-3 (GPC3 +), мембранному белку, сверхэкспрессируемому при гепатоцеллюлярной карциноме. Мы обнаружили, что модифицированные hGC33 НЧ (hGC33-SFB-NP) нацелены на GPC3 + Клетки гепатоцеллюлярной карциномы (ГЦК) путем специфического связывания с GPC3 на поверхности клеток ГЦК, ингибировали передачу сигнала, индуцированную Wnt, и ингибировали клетки ГЦК в G0 / 1 путем подавления экспрессии циклина D1, тем самым ослабляя миграцию клеток ГЦК за счет ингибирования эпителиальных клеток. мезенхимальный переход. hGC33-SFB-NP ингибировал миграцию, развитие цикла и пролиферацию клеток HCC путем ингибирования пути Ras / Raf / MAPK и пути Wnt в тандеме с молекулами GPC3, соответственно. hGC33-SFB-NP подавлял рост рака печени in vivo и улучшал выживаемость мышей с опухолями. Мы пришли к выводу, что hGC33 увеличивает нацеливание SFB-NP на клетки HCC. hGC33-SFB-NP синергетически подавляет прогрессирование HCC, блокируя путь Wnt и путь Ras / Raf / MAPK.
Введение
Глипикан3 (GPC3) представляет собой протеогликан гепарансульфата, экспрессируемый на поверхности клеток посредством механизма, включающего глицерофосфатидилинозитоловые якоря [1]. Хотя GPC3 экспрессируется во многих тканях во время развития, его экспрессия, по крайней мере, частично ингибируется в большинстве взрослых тканей метилированием ДНК в промоторной области [2]. Однако белок GPC3 сверхэкспрессируется примерно у 70% пациентов с гепатоцеллюлярной карциномой (ГЦК) [3, 4] и стимулирует классическую трансдукцию сигнала Wnt [5] посредством взаимодействия с лигандом Wnt, который способствует связыванию Wnt / frizzled, способствуя росту HCC [6]. . Активация классического сигнального пути Wnt - одно из наиболее частых событий, связанных со злокачественной трансформацией ГЦК [7, 8]. Основываясь на способности GPC3 усиливать передачу сигналов Wnt, мы предположили, что избыточная экспрессия GPC3 способствует росту HCC, стимулируя классический путь Wnt.
Терапевтическое моноклональное антитело, которое распознает эпитоп в C-концевой части GPC3 (524–563) (hGC33), которое распознает C-концевой эпитоп GPC3 (524–563), ингибирует рост опухоли в подкожных ксенотрансплантатах HepG2 и Huh- 7 у мышей [9, 10]. hGC33 был гуманизирован путем трансплантации дополнительной области принятия решения, и его противораковое действие столь же эффективно, как hGC33 для ксенотрансплантации HepG2, а hGC33 использовался в клинических испытаниях [11]. Эти результаты предполагают, что hGC33 обладает важной противоопухолевой активностью, и лечение анти-GPC3 будет напрямую ингибировать пролиферацию и / или выживание клеток HCC путем блокирования Wnt и / или других сигнальных путей.
Однако из-за сложного патогенеза ГЦК эффективность блокирования только GPC3 ограничена [12]. Следовательно, изучение детального механизма патогенеза ГЦК и определение многообещающих биомаркеров для диагностики и прогноза ГЦК может помочь обеспечить эффективные терапевтические цели и улучшить прогноз пациентов. Было высказано предположение, что антитела против GPC3 повышают чувствительность ГЦК к химиотерапевтическим агентам [13]. Противоопухолевая активность hGC33 в сочетании со стандартными химиотерапевтическими препаратами была недавно оценена [14]. Сорафениб - это новый пероральный препарат для лечения ГЦК, который может напрямую ингибировать пролиферацию опухолевых клеток путем блокирования клеточного сигнального пути, опосредованного RAF / MEK / ERK, для сдерживания роста опухоли [15, 16]. Однако нецелевое распределение сорафениба in vivo и аномально активированные сигналы Wnt при ГЦК ограничивают эффективность препарата и усиливают его побочные эффекты [14,15,16]. Wnt связывается с рецепторами семейства Frizzled, чтобы активировать внутриклеточную передачу сигнала, который регулирует пролиферацию клеток, апоптоз и миграцию клеток, и вызывает лекарственную устойчивость во многих опухолях, таких как HCC [17,18,19].
В моделях ксенотрансплантации HepG2 комбинация hGC33 и сорафениба более эффективна в подавлении роста опухоли, чем один сорафениб [15], а доставке лекарств с помощью полимерных наноносителей уделяется большое внимание при лечении рака. Наносители, содержащие противораковые лекарственные средства, могут предотвращать неспецифическое распределение и неспецифическую деградацию in vivo, улучшать биодоступность лекарств и направлять противоопухолевые препараты, а также упрощать оценку фармакокинетики и лечения [15, 16]. Во множестве наночастиц на основе полимеров составы на основе поли (молочной и гликолевой кислоты) (PLGA) считаются идеальными и безопасными носителями лекарств [17, 18]. В связи с этим полиэтиленгликоль - b -поли (d, l-лактид-со-гликолид) (ПЭГ- b -PLGA) основаны на сополимерах полиэтиленгликоля и PLGA, которые безопасны и нетоксичны после гидролиза и были одобрены Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов США [19,20,21,22]. Следовательно, внутривенная инъекция с наноносителем ПЭГ- b Сополимер -PLGA - многообещающая стратегия для достижения адресной доставки и повышения эффективности. Кроме того, нанесение специфического антитела hGC33 против молекул GPC3 на клеточную мембрану HCC может не только улучшить доставку нанопрепаратов, нацеленных на in vitro и in vivo [18, 19], но также заблокировать Wnt и / или другие сигнальные пути, связанные с GPC3, ингибировать пролиферации и / или выживаемости раковых клеток, и может достигать синергетической противоопухолевой активности.
В этом исследовании мы изучали, может ли модифицированный антителами hGC33 сополимер PEG- b Наночастицы -PLGA могут облегчить доставку сорафениба (hGC33-SFB-NP) к HCC in vivo и in vitro, а также повысить эффективность лечения HCC за счет HCC-активного нацеливания для изменения фармакокинетики лекарства. В соответствии с размером частиц, дзета-потенциалом, морфологией частиц, эффективностью захвата лекарственного средства, емкостью загрузки лекарственного средства и высвобождением лекарственного средства in vitro целевые NP были всесторонне охарактеризованы. Способность нацеливания in vitro характеризуется захватом клетками гепатомы HepG2. Биораспределение и синергетический терапевтический эффект hGC33-SFB-NP на HCC оценивали путем сравнения сорафениба и SFB-NP. Наши результаты показали, что hGC33-SFB-NP может нацеливаться на GPC3 + HCC. Он может подавлять развитие клеточного цикла, пролиферацию клеток и инвазию опухолей, подавляя сигнальные пути Wnt и Ras / Raf / MAPK, и синергетически подавлять прогрессирование рака печени.
Материалы и методы
Материалы
Антитело hGC33, 3- (4,5-диметилтиазол-2-ил) -2,5-дифенилтетразолий бромид (МТТ), 4 ', 6-диамидино-2-фенилиндол, дигидрохлорид (DAPI), 5,5', 6 , 6'-тетрахлор-1,1 ', 3,3'-тетраэтилбензимидазолилкарбоцианина иодид (JC-1) и диметилсульфоксид (ДМСО) были получены от Sigma-Aldrich, Inc. (Сент-Луис, Миссури). 1- (3-диметиламинопропил) -3-этилкарбодиимид гидрохлорид и N -гидроксисукцинимид были получены от Qiyun Biotech (Гуанчжоу, Китай). Набор для количественного определения белка бицинхониновой кислоты (BCA), кумарин-6 и набор для обнаружения апоптоза Annexin V-FITC / PI были приобретены у Beyotime Biotechnology (Шанхай, Китай). ПЭГ- b -ПЛГА-диблок-сополимер малеимида (мал-ПЭГ- b -PLGA; 25 000–30 000 Да, PLGA, LA:GA, w / w; ПЭГ, 13–15%) был приобретен в Polyscitech (West Lafayette, IN, USA). Набор антител PI3K (9655 #), набор антител p-Akt (9916 #), набор антител mTOR (9964 #), набор антител семейства Bcl-2 (9942 #), набор антител к апоптозу (9915 #) и вторичные козьи анти- кроличьи и антимышиные антитела были приобретены в Cell Signal Technology (Danvers, MA, США); циклин B1 и циклин-зависимая киназа были приобретены в Abcam Biological Technology (США). Антитела к фосфо-Rb, циклину D1, киназе контрольной точки 1 (CHK1), P53, фосфорилированному гену 1 восприимчивости к раку молочной железы (p-BRCA1), RAD51, цитохрому C и матриксной металлопротеиназе (MMP2 и MMP9) были приобретены у Abcam (США). . Все остальные химические вещества, реагенты и растворители аналитической чистоты были получены от стандартных поставщиков и использовались без дополнительной очистки.
Клетки и животные
Клеточная линия ГЦК HepG2 (полученная из американской коллекции типовых культур (Манассас, Вирджиния, США) культивировалась в среде DMEM (Invitrogen, Карлсбад, Калифорния, США) с добавлением 10% FBS (HyClone, Логан, Юта, США) и 80 Ед / мл пенициллина и 80 мкг / мл стрептомицина во влажной атмосфере с 5% CO 2 при 37 ° С. Голых мышей BALB / C массой 20–22 г (5–6 недель) предоставила компания Nanjing Junke Biotechnology Co., Ltd. (Китай). Бестимусных мышей BALB / C выращивали в помещении для SPF. Весь уход за животными и лечение проводились в соответствии с требованиями по уходу за животными Аньхойского университета науки и технологий. Все протоколы экспериментов были рассмотрены и одобрены комитетом по этике экспериментов на животных Аньхойского университета науки и технологий (номер разрешения:2019dw013).
Подготовка НП
Для получения НЧ мал-ПЭГ- b -PLGA и SFB или кумарин-6 взвешивали и растворяли в органической фазе (3:2 об. / Об. Дихлорметан / ацетон). Раствор добавляли к раствору поливинилового спирта (ПВС) (5% мас. / Об.) По каплям при непрерывном встряхивании. Смесь периодически обрабатывали ультразвуком с помощью зондового ультразвукового устройства (выходная мощность 550 Вт, 8 раз) на льду для создания водонефтяной эмульсии. Эмульсию добавляли к раствору ПВС (1% мас. / Об.) При перемешивании магнитной мешалкой. НЧ SFB и кумарин-6 собирали центрифугированием при 8000 об / мин в течение 30 минут и трижды промывали водой Milli-Q.
Для генерации hGC33-NP за счет тиоэфирных связей, образованных реакцией малеимида со свободными сульфгидрильными остатками в антителе hGC33, антитело hGC33 смешивали с функционализированными малеимидом НЧ в молярном соотношении 5:1 (hGC33:mal-PEG- b -PLGA) и инкубировали при 4 ° C в течение 16 ч при непрерывном перемешивании. hGC33 конъюгировали с NP посредством реакции сульфгидрильных групп антитела hGC33 с малеимидными группами цепей PEG. Неконъюгированные антитела удаляли пропусканием через колонки с сефарозой CL-4B. Эффективная конъюгация белков была подтверждена с помощью набора BCA (Thermo Fisher Scientific, Уолтем, Массачусетс, США).
Характеристика наночастиц (НЧ)
Морфология, размер частиц, эффективность инкапсуляции (EE) и стабильность NP
Морфологию НЧ оценивали с помощью просвечивающей электронной микроскопии (ТЕМ, H-600; Hitachi, Токио, Япония). НЧ-НЧ без лекарств и НЧ без лекарств регистрировали с помощью ИК-Фурье спектрофотометра (Thermo Nicolet, Мэдисон, Висконсин, США) с использованием бромида калия. Средний размер частиц и дзета-потенциал НЧ были охарактеризованы с помощью Malvern Zetasizer ZEN3600 Nano ZS (Malvern Instruments, Malvern, UK) при 20 ° C. Эффективность инкапсуляции лекарственного средства (EE) и эффективность загрузки лекарственного средства (LC) оценивали с помощью ультрафильтрации. Образцы загружали в устройство для ультрафильтрации (100 kMWCO; Sartorius, Göettingen, Germany) и центрифугировали при 8000 об / мин в течение 25 минут при 4 ° C для удаления свободного лекарственного средства. Одинаковый объем каждого образца растворяли в ацетонитриле для подтверждения общего количества лекарственного средства. Концентрация была измерена с помощью высокоэффективной жидкостной хроматографии. Длина волны поглощения 266 нм. Для расчета EE лекарственного средства (%) для NP использовали следующую формулу:(масса захваченного лекарственного средства / общая масса лекарственного средства) × 100%. LC (%) рассчитывали как (масса инкапсулированного лекарственного средства / масса NP) × 100%. Чтобы понять стабильность НЧ при комнатной температуре, изменение размера НЧ оценивали с помощью динамического рассеяния света (ДРС) в заранее определенные моменты времени (0,5, 1, 2, 4, 8, 12 ч, 16 ч, 20 ч и 24 часа). з) при 25 ° С.
Анализ высвобождения лекарства и клеточного поглощения in vitro
Препарат из НЧ исследовали с использованием диализных мешков с отсечкой по молекулярной массе 10 кДа. Вкратце, 1 мл наночастиц загружали в диализный мешок (MWCO 8 000–10 000 Да; Spectrum Labs Inc., Калифорния, США). Мешки для диализа погружали в PBS и перемешивали магнитной мешалкой при 25 ° C. Профили высвобождения лекарственного средства для НЧ измеряли в 100 мл 0,2 М фосфатно-солевого буфера (PBS; pH =7,4) в течение 7 дней. Концентрацию лекарственного средства в образцах измеряли с помощью высокоэффективной жидкостной хроматографии. В последующих исследованиях hGC33-кумарин 6-NP, имеющий тот же размер частиц, что и hGC33-SFB-NP, был использован для оценки нацеливания hGC33-SFB-NP. HepG2 (GPC3 + ) и Ли-7 (GPC3 - ) инкубировали с hGC33-кумарином 6-NP в течение 0,5, 2 или 4 ч при 37 ° C в 5% CO 2, соответственно. Совместно культивируемые клетки промывали и фиксировали 4% формальдегидом в течение 10 мин; ядра клеток окрашивали 5 мкг / мл Hoechst 33,342 в течение 15 мин для определения местоположения внутриклеточных НЧ. Конфокальная микроскопия для анализа изображений внутриклеточных наночастиц была проанализирована с помощью конфокальной микроскопии (Olympus FV1000; Olympus Corporation, Токио, Япония).
In Vitro Cellular Effect
Цитотоксичность свободного hGC33 (Ab), свободного SFB, hGC33-null-NP или hGC33-SFB-NP определяли с помощью анализа MTT. HepG2 (GPC3 + ) ячеек и Li-7 (GPC3 - ) клетки в логарифмической фазе высевали в 96-луночный планшет при плотности 4000 клеток на лунку с последующей инкубацией в течение 48 часов при 37 ° C в 5% CO 2 . Клетки обрабатывали hGC33, свободным SFB, hGC33-null-NP или hGC33-SFB-NP в течение 48 часов при 37 ° C в 5% CO 2 . После совместного культивирования в течение определенного времени активность против пролиферации клеток определяли с помощью анализа МТТ, как описано [20]. Оптическую плотность каждой лунки измеряли при 490 нм и вычисляли половину значения максимальной ингибирующей концентрации (IC 50) с помощью SPSS 17.0.
Измерение способности вторжения клеток
Клетки в фазе логарифмического роста высевали на 6-луночные планшеты с плотностью 5 × 10 4 . клеток / лунку, царапают всасывающей головкой и заменяют бессывороточной культуральной средой. Заживление ран регистрировали через 0, 24 и 48 ч в контрольной и экспериментальной группах. В то же время такое же количество клеток инокулировали в камеры Transwell и обрабатывали свободным hGC33, свободным SFB, hGC33-null-NP или hGC33-SFB-NP. В нижнюю камеру добавляли пятьсот микролитров 10% среды FBS. После инкубации в течение 24 ч камеру Transwell вынимали, клетки фиксировали 4% параформальдегидом и окрашивали 0,1% кристаллическим фиолетовым. Для оценки способности к миграции были рассчитаны размер заживающей раны и количество мигрирующих клеток.
Определение цикла ячейки
После инкубации в течение ночи в 6-луночных планшетах клетки обрабатывали свободным hGC33 (Ab), свободным SFB, hGC33-null-NP или hGC33-SFB-NP в течение 24 часов, затем собирали и фиксировали этанолом. После окрашивания йодидом пропидия проводили проточную цитометрию и анализировали клеточный цикл с помощью modifit 3.0 (Verity Software House, Топшам, штат Мэн).
Вестерн-блоттинг
Для оценки статуса активации сигнального пути и экспрессии молекул-мишеней клетки инкубировали в течение ночи в 6-луночных планшетах, и свободный hGC33, свободный SFB, hGC33-null-NP или hGC33-SFB-NP применяли в течение 24 часов. . Клетки каждой группы обработки собирали, белок экстрагировали и измеряли. Концентрацию белка измеряли и калибровали с помощью набора белков BCA (Biosharp, Хэфэй, Китай). Белок в образцах отделяли электрофорезом в 12-алкилсульфатном полиакриламидном геле, переносили на PVDF-мембрану и запечатывали обезжиренным молоком. Первое антитело (разведенное 1:1000) инкубировали в течение ночи при 4 ° C, а вторые антитела (1:2000) инкубировали в течение 1 ч при комнатной температуре. Полосы визуализировали с использованием субстратов ECL (Thermo Fisher Scientific Waltham, Массачусетс, США), изображения отображали с помощью системы анализа изображений геля, а β-актин использовали в качестве контроля.
Противоопухолевое действие in vivo
Определяли ингибирование свободного hGC33, свободного SFB, hGC33-null-NP, SFB-NP и hGC33-SFB-NP на рост HCC in vivo. В соответствии с правилами и руководящими принципами по здоровью животных комитета по этике Технологического университета Аньхой, все эксперименты проводились на мышах BALB / c в клетках в помещении с контролем температуры (23 ± 2 ° C) при 12 ч / 12 ч света / темный цикл. 50-мкл суспензии, содержащей 5 × 10 6 Живые клетки HepG2 вводили подкожно в правую часть живота 5-недельным самкам мышей BALB / c (20–22 г). Когда объем опухоли достигал примерно 50 мм 3 , мышей случайным образом разделили на 6 групп (по 10 мышей в каждой группе). Нормальный физиологический раствор контрольного NS (200 мг / кг нулевых NP в 200 мкл PBS), hGC33-null-NP (hGC33-null-NP в 200 мкл PBS, эквивалент hgc33 =100 мкг / кг / время), свободный hGC33 (hGC33 в 200 мкл PBS, 100 мкг / кг / раз), свободный SFB (доза SFB:8 мг / кг / раз), SFB-NP (доза SFB:8 мг / кг / раз) и hGC33-SFB-NP (эквивалент SFB =8 мг / кг / время, hGC33 =100 мкг / кг / время) вводили через хвостовую вену каждые 2 дня 10 раз. Вес и размер опухоли мышей измеряли каждые четыре дня. Формула расчета объема опухоли:объем =0,5 × л × Вт 2 , где L и W обозначают длину и ширину опухоли соответственно. Через четыре недели после введения животных анестезировали диэтиловым эфиром и измеряли размер и вес опухоли. Кроме того, были удалены опухоль, сердце, печень, почки, легкие и селезенка, зафиксированы 4% раствором параформальдегида, залиты парафином, сделаны срезы и окрашены гематоксилином и эозином для оценки гистологических изменений с помощью цифровой микроскопии.
Статистический анализ
Данные представлены в виде среднего значения ± стандартное отклонение (SD) и были оценены с помощью дисперсионного анализа с помощью SPSS 18.0. Парные статистические сравнения были выполнены с использованием двустороннего критерия t Стьюдента. Различия считались статистически значимыми для P <0,05.
Результаты
Характеристика НЧ и высвобождения лекарства In Vitro
Поскольку диаметр и поверхностные свойства NP влияют на поглощение клетками, высвобождение лекарства и распределение NP in vivo, мы охарактеризовали полученные NP с соответствующими параметрами. Морфология, размер частиц и распределение частиц по размерам полимеров SFB-NP, hGC33-null-NP и hGC33-SFB-NP суммированы на рис. 1. Морфология hGC33-SFB-NP, наблюдаемая с помощью просвечивающего электронного микроскопа (рис. 1а) показаны жесткие ядра с нечеткими краями, что указывает на присутствие hGC33 на поверхности НЧ. Размеры частиц SFB NP, hGC33-null-NP и hGC33-SFB-NP варьировались от 100 до 150 нм и имели типичное одномодальное распределение частиц по размерам. Средний диаметр hGC33-SFB-NP (120,2 ± 10,2 нм) был немного больше, чем у hGC33-null-NP, SFB-NP и нулевого NP (рис. 1a, таблица 1). Тонкие сферические нечеткие пленки с единственной поверхностью присутствовали на поверхности hGC33-SFB-NP и hGC33-null-NP, что указывало на присутствие антитела hGC33 на поверхности NP. Увеличенный размер hGC33-SFB-NP и hGC33-null-NP подтвердил существование пленки hGC33. Синтез hGC33-SFB-NP был подтвержден с помощью 1 H-ЯМР (рис. 1б) перед приготовлением НЧ. Пики при 5,2 м.д. и 1,58 м.д. были отнесены к протонам -CH3 молочной кислоты; пик при 4,8 м.д. был отнесен к -OCH2- гликолевой кислоты; а пик при 3,6–3,8 м.д. был отнесен к протонам -CH2CH2O- повторяющихся звеньев ПЭГ. Химия поверхности ПЭГ- b -ПЛГА и Ab-ПЭГ- b НЧ -ПЛГА также исследовали с помощью ИК-Фурье спектроскопии (рис. 2в). В спектре ПЭГ- b -Полимер PLGA, прочная полоса примерно при 1750 см -1 происходит от участка карбонильных групп (C =O) в цепи PLGA. Полоса на 2880 см −1 был обусловлен растяжением группы a –CH в цепи ПЭГ. При этом появился пик на 2520 см −1 который мы приписали пику растяжения -SH антитела hGC33. 1 Результаты H ЯМР и FTIR показали, что антитело было привито к остову PEG- b -ПЛГА полимеры.
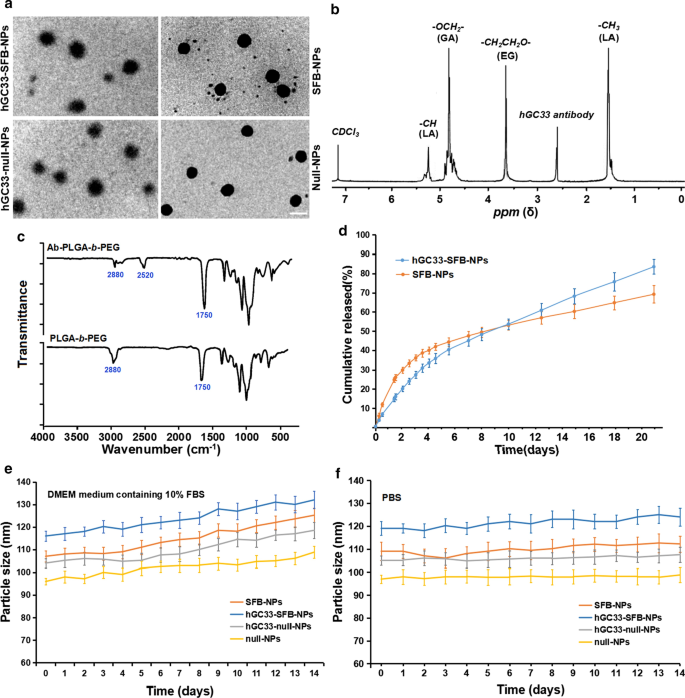
Характеристика НП. а ПЭМ-характеристика НЧ, шкала показывает 100 нМ; б 1 Спектры ЯМР 1Н синтетического hGC33-PLGA- b -ПЭГ в CDCl3; c ИК-Фурье спектры hGC33-PEG- b -PLGA и ПЭГ- b - PLGA; г кумулятивные профили высвобождения SFB-NP и hGC33-SFB-NP в PBS (pH =7,4) при 37 ° C; е изменения размера НЧ, инкубированных в среде DMEM, содержащей 10% FBS, в течение 14 дней; е изменения размера НЧ, инкубированных в PBS в течение 14 дней. SFB, сорафениб; ПЭМ, просвечивающая электронная микроскопия; НЧ, наночастицы; 1 H ЯМР, 1 Спектроскопия H-ядерного магнитного резонанса; FTIR, инфракрасная спектроскопия с преобразованием Фурье
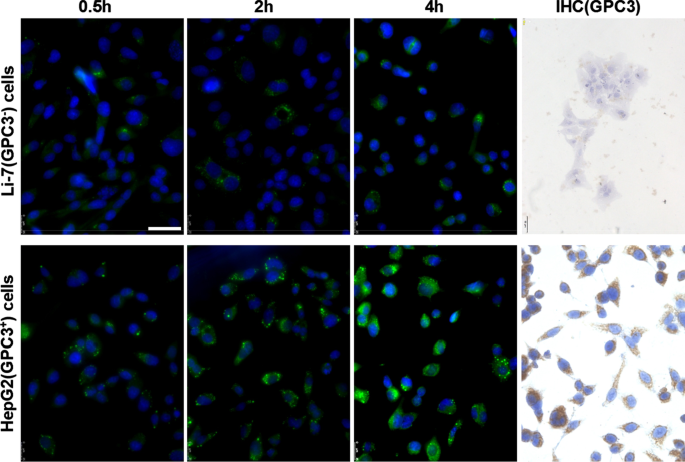
Экспрессия GPC3 и захват hGC33-кумарин6-NP в клетках Li-7 и HepG2. Клетки, инокулированные в культуральный планшет, промывали PBS и инкубировали со 100 мкг / мл hGC33-кумарин6-NP в среде DMEM в течение 2, 4 и 8 часов. Ядра окрашивали DAPI, клетки фиксировали и детектировали с помощью конфокальной лазерной сканирующей микроскопии. GPC3 не обнаруживался в клетках Li-7, но был высоко экспрессирован в клетках HepG2, как обнаружено иммуноцитохимическим методом. Масштабная линейка показывает 50 мкМ
Интересно, что наночастицы hGC33-SFB-NP и SFB-NP в среде DMEM, содержащей 10% FBS (pH =7,4), имели быстрое высвобождение лекарства примерно до 4 дней, а затем относительно медленное и стабильное высвобождение лекарства; кумулятивное высвобождение SFB hGC33-SFB-NP и SFB-NP за 20 дней составило около 77% и 65% соответственно (рис. 1d). Разница может быть связана с гидрофильной молекулой на поверхности ПЭГ- b Матрица -PLGA, которая может ускорить разложение наночастиц за счет увеличения гидратации и, тем самым, содействия гидролизу. Чтобы определить стабильность полученных НЧ, hGC33-SFB-NP, hGC33-нулевой NP, нуль-NP и SFB-NP помещали в среду DMEM, содержащую 10% FBS (pH =7,4) и в PBS (pH =7,4). ). Размеры различных НП оставались стабильными более 2 недель. SFB высвобождался из hGC33-SFB-NP длительным и стабильным образом в течение 14 дней, но было небольшое изменение размера частиц в среде DMEM по сравнению с таковым в 10% FBS (рис. 1e, f). Стабильность hGC33-SFB-NP подходит для SFB, играющего устойчивую биологическую роль.
Большой дзета-потенциал может вызвать сильное электростатическое отталкивающее взаимодействие между НЧ и поддерживать стабильность дисперсионной системы НЧ [23, 24]. Как показано в Таблице 1, дзета-потенциалы hGC33-SFB-NP, hGC33-null-NP и SFB-NP составляют -18,2 ± 2,2 мВ, -18,5 ± 1,8 мВ и -15,9 ± 2,1 мВ, соответственно, что может быть вызванным отрицательными зарядами, генерируемыми альдегидной группой, карбоксильной группой и фосфатной группой в гликопротеине антитела hGC33. Отрицательно заряженные НЧ могут привести к высокой стабильности суспензии НЧ. Кроме того, поскольку поверхность клеток отрицательно заряжена в нормальных физиологических условиях, полученные НЧ отталкивают клетки с низким зарядом и менее токсичны для тканей и клеток. Кроме того, распределение по размерам нулевых NP и SFB-NP (индекс полидисперсности [PDI] 0,18 и 0,19, соответственно) было немного, но не значительно меньше, чем у hGC33-null-NP (PDI =0,21) и hGC33-SFB- NP (PDI =0,23). Таким образом, размер наночастиц имеет хорошую однородность. Размер частиц, гранулометрический состав и дзета-потенциал NP показаны в таблице 1 в соответствии с результатами эффективности инкапсуляции SFB и содержания загрузки. Неконъюгированное антитело к GPC3 hGC33 удаляли ультрацентрифугированием и оценивали эффективность связывания антитела hGC33 с NP. Анализ белка BCA показал, что эффективность связывания антитела hGC33 с НЧ составила 79,5% ± 2,9%.
hGC33-Coumarin 6-NP эффективно нацелен на GPC3 + Клетки HepG2 линии клеток HCC
Чтобы выяснить, обладает ли еще антитело наночастиц hGC33 способность специфически нацеливаться на GPC3, мы использовали GPC3 + HepG2 и GPC3 - Клетки Li-7 в качестве клеток-мишеней и hGC33-кумарин 6-NP в качестве индикаторных наночастиц и инкубировали различные клетки в течение 2, 4 и 8 часов. Клетки промывали PBS 3 раза и реагировали с DAPI для окрашивания ядра. Клетки фиксировали и фотографировали с помощью флуоресцентного микроскопа Leica (DMi8, Германия). Было обнаружено, что зеленая флуоресценция в клетках HepG2 была значительно выше, чем в клетках Li-7 при том же времени инкубации (рис.2), что указывает на то, что количество hGC33-кумарин 6-NP, попадающее в клетки HepG2, было значительно выше, чем у клеток. в ячейках Li-7. Результаты документально подтвердили, что антитело hGC33 к hGC33-Coumarin6-NP по-прежнему обладает способностью нацеливаться на GPC3 и опосредовать интернализацию наночастиц. Экспрессию GPC3 в клетках HepG2 и Li-7 исследовали с помощью непрямой флуоресценции и цитохимического окрашивания. Результаты показали, что клетки HepG2 сверхэкспрессируют GPC3, тогда как клетки Li-7 не экспрессируют GPC3 (рис. 2).
hGC33-Null-NP подавляет пролиферацию клеток HepG2
Чтобы определить, может ли hGC33-модифицированный NP (hGC33-null-NP) ингибировать рост HCC, мы измерили ингибирование роста клеток GPC3-положительной линии клеток HCC HepG2 и GPC3-отрицательной клеточной линии Li-7. Мы обнаружили, что как hGC33-null-NP, так и hGC33 подавляли рост GPC3-положительной линии клеток HCC HepG2 после 24 часов обработки, но hGC33 оказывал более значительное ингибирующее действие на клетки HepG2, чем hGC33-null-NP (рис. 3a).; Напротив, hGC33-null-NP и hGC33 не влияли на рост GPC3-отрицательной клеточной линии Li-7 (фиг. 3b). Характерный временной ход hGC33-null-NP (эквивалент 0,1 мкм hgc33) и hGC33 (0,1 мкм) на пролиферацию клеток HepG2 проиллюстрирован на фиг. 3c. Поскольку hGC33 представляет собой C-концевой пептид, который распознает GPC3, hGC33 превосходит hGC33-null-NP в ингибировании HepG2. Результаты предполагают, что молекула hGC33 в hGC33-null-NP может обладать эффектом блокирования пространства, что может влиять на способность связывания hGC33 с эпитопами.
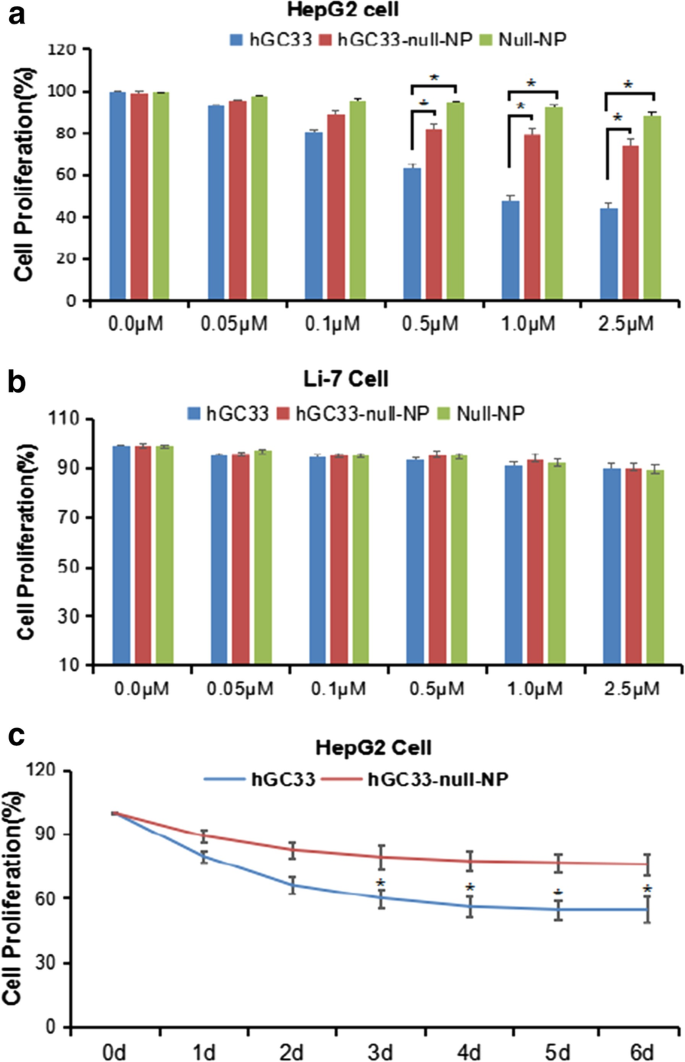
Разрастание клеток HepG2 и Li-7 ингибировалось hGC33, null-NP и hGC33-null-NP. а Тест на пролиферацию клеток проводили на GPC3-положительных клетках HepG2, обработанных hGC33, null-NP или hGC33-null-NP; б Тесты на пролиферацию клеток проводили на GPC3-отрицательных клетках Li-7, обработанных hGC33, null-NP и hGC33-null-NP. Клетки инкубировали с 0–2,5 мкМ hGC33, null-NP или hGC33-null-NP в течение 1 дня. Пролиферацию клеток измеряли методом МТТ и стандартизировали как необработанные клетки. Все значения представляют собой среднее значение ± стандартное отклонение. По сравнению с контрольной группой (0 мкМ) без обработки антителами * P <0,01; c репрезентативные результаты ответа HepG2 на hGC33, null-NP и hGC33-null-NP при лечении hGC33 (1,0 мкМ)
hGC33-Null-NP ингибирует клеточный цикл клеток HepG2
Чтобы понять механизм молекулярной активности антитела hGC33, модифицированного на наночастицах hGC33-null-NP, мы изучили развитие клеточного цикла HepG2, обработанного hGC33-null-NP. В клеточной линии HepG2 hGC33-null-NP и hGC33 значительно увеличивают долю клеток в G1 (рис. 4a, b), указывая на то, что как hGC33-null-NP, так и hGC33 могут вызывать остановку клеточного цикла в фазе G1. Более того, hGC33-null-NP и hGC33 могут значительно подавлять экспрессию cyclinD1 в клетках HepG2 (рис. 4c, d).
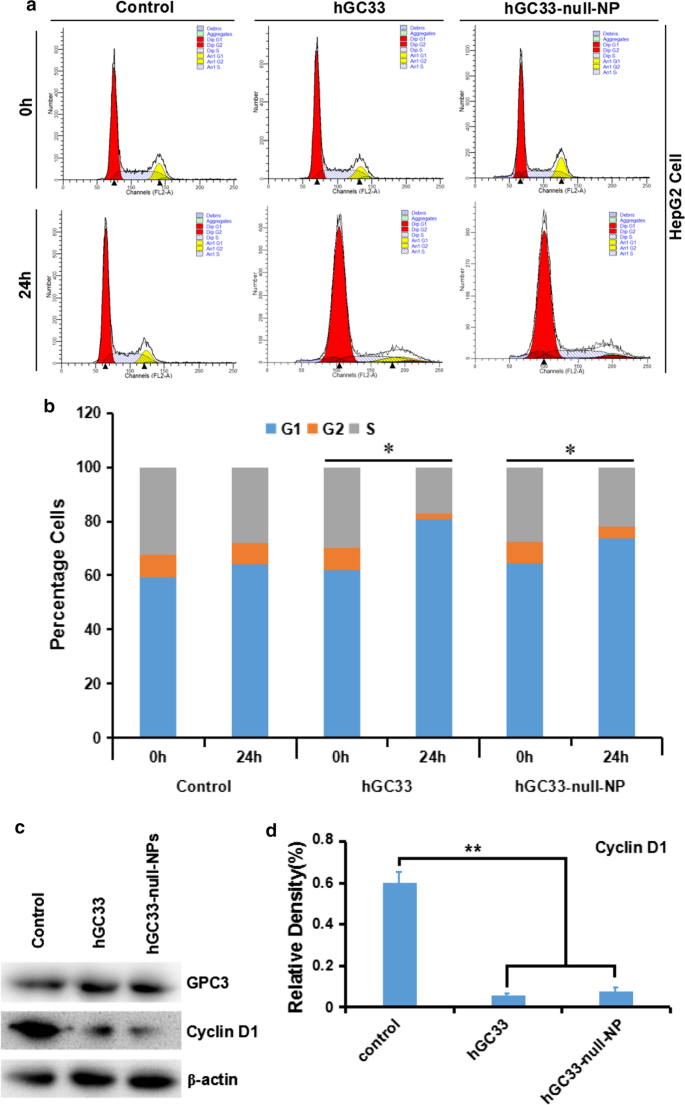
hGC33-null-NP и hGC33-индуцированная остановка клеточного цикла в фазе G1 и ингибирование экспрессии cyclinD1 в клетках HepG2. а Типичная диаграмма клеточного цикла различных групп, получавших hGC33-null-NP и hGC33. б Анализ клеточного цикла различных групп, получавших hGC33-null-NP и hGC33. Клетки HCC инкубировали с 0,5 мкМ hGC33 или hGC33-null-NP (с молярной концентрацией hGC33 в качестве сравнения). * P <0,05, фазу G1 hGC33-null-NP или hGC33 клеток сравнивали с фазой 0 h клеток. c , d После обработки hGC33-null-NP или hGC33, cyclinD1 значительно подавлялся в клетках HepG2 по сравнению с контрольной группой
Активация Wnt в клетках HepG2, Huh-7 и Li-7
Чтобы понять активацию классического сигнала Wnt / β-катенина в клетках HCC, мы сначала обнаружили экспрессию лиганда Wnt и рецепторного извитого белка (frizzled, FZD или Frz) в различных клеточных линиях HCC:HepG2 (GPC3 ++ ), Ха-7 (GPC + ) и Ли-7 (GPC3 - ). Результаты показали, что рецепторы Wnt3a и FZD, передающие путь β-катенина, экспрессируются во всех линиях клеток, особенно в клетках HepG2 и Huh-7 (рис. 5).
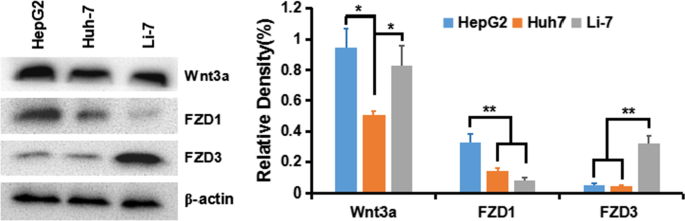
Экспрессия Wnt3a, FZD1 и FZD3 в линиях клеток HCC
hGC33-Null-NP ингибирует зависимую от передачи сигнала Wnt пролиферацию клеток, индуцированную Wnt3a
Некоторые исследования показали, что внеклеточная часть GPC3 может быть корецептором Wnt, который способствует активации и передаче сигнала Wnt / β-катенина. Следовательно, когда HepG2 (GPC3 + ), Ха-7 (GPC3 + ) и Ли-7 (GPC3 - ) cells were co-incubated with hGC33 and hGC33-null-NP, the activation of Wnt/β-catenin signal was blocked by hGC33 and hGC33-null-NP, and proliferation of HepG2 and Huh-7, but not Li-7, in Wnt3a-conditioned medium was inhibited. That proliferation of Li-7 cells most likely is due to the absence of GPC3 on the surface of Li-7 cells (Fig. 6). Our results indicate hGC33 and hGC33-null-NP nanoparticles specifically bind to GPC3 molecules on cell membrane. hGC33 and hGC33-null-NP partially neutralize the mitogenic activity of Wnt3a and inhibit the Wnt/β-catenin signaling pathway. However, the inhibition of proliferation by hGC33-null-NP nanoparticles was less than that of hGC33. Perhaps the spatial structure of the nanoparticles interferes with hGC33-null-NP and limits the function of the hGC33 molecule on the nanoparticles so they cannot completely block the interaction between GPC3 and Wnt3a.

GPC3-activated Wnt signal transduction in HCC. Fifty percent Wnt3a DMEM medium was added with anti-wnt3a antibody, hGC33, or hGC33-null-NP. HepG2 (GPC3 ++ ), Huh-7 (GPC3 + ) and Li-7 (GPC3 − ) cells were co-incubated for 48 h, and cell proliferation was measured by MTT assay. The data were expressed as mean ± SD (*P < 0.01)
hGC33-Null-NP Inhibits Wnt3a-Induced Signal Transduction in HepG2 and Huh-7 Cells
To understand the effect of hGC33-null-NP on Wnt/β-catenin signaling in HCC cells, we extracted the proteins of HepG2 and Huh-7 cells treated with hGC33 or hGC33-null-NP. The results of western blot showed that after hGC33-null-NP treatment, the levels of pYAP and pβ-catenin were increased, but the levels of YAP and β-catenin were decreased. Furthermore, the levels of cyclinD1, CD44, VEGF, and c-MYC in the hGC33-null-NP group were lower than those in the control group, but the level was less than with hGC33 treatment. Similar effects were observed in HepG2 and Huh-7 cells, as shown in Fig. 7.

Inhibition of Wnt3a-induced β-catenin signaling by hGC33-null-NP or hGC33. а Compared with the control group, Wnt/β-catenin signaling pathway in HepG2 and Huh-7 cells treated with hGC33-null-NP or hGC33 was inhibited, and the levels of β-catenin and YAP were decreased, while the increased phosphorylated β-catenin and phosphorylated YAP molecules were unstable, and degraded in cytoplasm. The decreased β-catenin was difficult to maintain in the nucleus and drive the expression of CyclinD1, CD44, VEGF, and c-MYC, which resulted in the decrease of cyclinD1, CD44, VEGF, and c-myc protein levels. б The mechanism pattern of Wnt/β-catenin signaling pathway inhibited by hGC33-null-NP or hGC33
hGC33-SFB-NP or hGC33 Attenuates HCC Cell Migration by Inhibiting Epithelial Mesenchymal Transition (EMT)
HCC is one of the deadliest cancers in the world, and its incidence is steadily increasing. Sorafenib is the only approved standard treatment for patients with advanced HCC, as it has been shown to improve the survival rate of these patients. However, clinical and preclinical observations suggest that sorafenib therapy has limited efficacy due to the rapid development of drug resistance. Therefore, elucidating the mechanism of escape resistance to sorafenib is a major emphasis in HCC research. In recent years, more and more attention has been paid to the role of epithelial mesenchymal transition (EMT) in the progress of HCC and the development of drug resistance. EMT refers to the transformation of epithelial to mesenchymal cells, which endows cells with the ability metastasize and invade, including acquisition of stem cell characteristics, reducing apoptosis and aging, promoting immunosuppression, and participating in the occurrence and development of cancer. The loss of E-cadherin expression is considered a key step in the carcinogenesis and EMT. EMT is a developmental multi-step molecular and cellular reprogramming process that cancer cells use to achieve aggression. This is mainly through down regulating the expression of E-cadherin, keratin, mucin, ZO-1 (tight junction protein); up regulating the expression of vimentin, alpha-smooth muscle actin (α-SMA), FN fibronectin, MMPs (degradation matrix), N-cadherin, snail, slug, twist, Rho, TGF-β, FGF, type I collagen, and type II collagen to achieve the invasion, metastasis, and anti-apoptosis of EMT characteristic tumor. The changes of these protein expressions mainly involve the activation of Wnt/β-catenin and Ras/Raf/MAPK signaling pathways.
Our experiments have shown that hGC33 antibody on the surface of NP vector can inhibit Wnt3a-induced β-catenin signal transduction, and then down regulate the expression of β—catenin, CD44, vascular endothelial growth factor (VEGF), cyclin D1, and c-MYC. Furthermore, hGC33-SFB-NP inhibits the activation of Ras/Raf/MAPK signal pathway and inhibits proliferation and apoptosis of HCC cells. hGC33 and SFB have a synergistic effect, inhibiting EMT and decreasing the migration of HCC cells (Fig. 8).

Effect of hGC33-SFB-NP on EMT inhibition. а Compared with the control group, the hGC33-SFB-NP treatment group had less cell migration. Photographs were taken under an optical microscope (magnification, × 200). The error value represents the standard deviation of three independent experiments. *Compared with the control group, p <0,01. б Compared with the control group, the EMT-related proteins snail, vimentin, and MMP-2 in HCCs treated with hGC33-SFB-NP decreased, whereas E-cadherin increased. c Molecular mechanism of EMT. EMT, epithelial–mesenchymal transition; MMP-2, matrix metalloproteinase-2; SFB, sorafenib; NP, nanoparticle
hGC33-SFB-NP Inhibits the Growth of Hepatocellular Carcinoma In Vivo and Improves the Survival Rate of Tumor-Bearing Mice
To evaluate the anti-tumor activity of hGC33-SFB-NP in vivo, HepG2 and Huh-7 cells were inoculated subcutaneously into the right abdomen and dorsal side of female BALB/c nude mice, respectively. When the tumor xenograft growth reached about 30 mm 3 , the mice were randomly divided into groups to further evaluate the inhibition of each group (hGC33-SFB-NP, hGC33-null-NP, SFB-NP, free hGC33, free SFB, and control group) HCC effect (n = 5 per group). It can be seen from Fig. 9a, b that hGC33-SFB-NP significantly slowed tumor growth in mice compared with the PBS control and other treatments. Compared with the PBS control, hGC33-null-NP, SFB-NP, free hGC33, and free SFB also had some inhibition of HCC, which is because free hGC33 and free SFB directly inhibit Wnt signal and Ras/Raf/MAPK, respectively. Such pathways can inhibit the proliferation of HCC cells to a certain extent. Although the nanoparticle-modified hGC33 (hGC33-null-NP) is connected to the nanosurface through chemical bonds, it did not affect hGC33′s targeting of GPC3 molecules and inhibition of Wnt activity. Nanoparticle-loaded SFB (SFB-NP), after being endocytosed by cells, was degraded to release SFB from the copolymer to inhibit the growth of HCC. In all, the inhibitory effect of hGC33-SFB-NP on HepG2 cell grafts was, as expected, more than on Huh-7 cell grafts, probably because HepG2 expresses GPC3 molecules.

The effect of hGC33-SFB-NP on xenotransplantation of HCC in nude mice and the changes of mice weight. Liver cancer cells were inoculated subcutaneously on the back of each nude mouse (n =10). After 10 days, the tumor bearing mice were treated with PBS (control), free hGC33, free SFB, hGC33-null-NP, SFB-NP, and hGC33-SFB-NP. Tumor size (a , b ) and body weight (c , d ) of mice were monitored at designated time points
The body weight of nude mice in each treatment group also was measured, as shown in Fig. 9c, d. The body weight of the control group decreased gradually. The weight of mice in free hGC33, free SFB, SFB-NP, and hGC33-null-NP treatment groups also decreased progressively and not significantly less than in the control group. However, the weight of nude mice bearing HepG2 and Huh-7 treated with hGC33-SFB-NP only slightly decreased, and the weight remained relatively stable during the treatment cycle. These results support that the novel hGC33-SFB-NP nanodrug has no significant toxicity in nude mice, and the SFB loaded on the nanocarrier and the surface modified hGC33 can produce additive or even synergistic anti-tumor effect.
Discussion
To examine the suitability of hGC33-SFB-NP for targeted HCC therapy, we tested the model conjugates for their ability to bind to human glypican-3 on HCC cells in vitro; to inhibit glypican-3-positive HCC cell proliferation, migration, and Wnt/β-catenin signal transduction; and inhibit HCC that overexpress glypican-3 in vivo.
To covalently bind GPC3-specific antibody hGC33 with mal-PEG-b -PLGA nanoparticles, we cross-linked the free sulfhydryl group in the Fc segment of hGC33 with maleimide functionalized PEG-b -PLGA (mal-PEG-b -PLGA) by forming stable thioether bonds. Conjugation is a prerequisite for targeting of GPC3-positive HCC. A series of experiments, including the changes of nanoparticle diameter and zeta potential detected by lens and the intracellular uptake of hGC33-SFB-NP, verified the targeting of hGC33-SFB-NP to HepG2 (GPC3 + ) клетки. These results indicated that the binding activity of antibody hGC33 was not altered by the conjugation.
We directly detected the phagocytic effect of GPC3 + HepG2 and GPC3 − Li-7 cells on PEG-b -PLGA NP surface-modified hGC33 by confocal microscopy. After HepG2 and Li-7 cells were co-cultured with hGC33-coumarin 6-NP, the green signal intensity in HepG2 cells was significantly higher than in Li-7 cells, indicating that there were more nanoparticles in the HepG2 cells. This finding is consistent with the hGC33 antibody modified on the surface of PEG-b -PLGA NP specifically binding to glypican-3 on the surface of HCC cells and being internalized. The efficiency of hGC33-modified NP internalization depends on the expression level of GPC3 antigen on the cell surface.
We used the standard MTT assay to measure the efficiency of inhibiting the proliferation of hepatoma cells. Both hGC33-null-NP and hGC33 inhibited the growth of the GPC3-positive HCC cell line HepG2, but hGC33-null-NP and hGC33 did not affect the proliferation of GPC3-negative Li-7 cells (Fig. 3b). At the animal level, hGC33-null-NP or hGC33 alone inhibited the growth of Huh-7 and HepG2 xenografts to a certain extent, while hGC33-SFB-NP caused growth arrest of Huh-7 and HepG2 hepatoma xenografts in mice. The finding that hGC33-null-NP significantly inhibited GPC3-positive hepatoma cells showed that the inhibitory effect of PEG-b -PLGA NP surface-modified hGC33 on HCC cell proliferation depends on the expression of GPC3 antigen on the cell surface.
GPC3 regulates many pathways in HCC pathogenesis, including Wnt and YAP signaling [25,26,27]. GPC3 interacts with Wnt ligand and may be a coreceptor for Wnt and facilitate Wnt/Frizzled binding for HCC growth [28, 29]. We further examined the effect of nanodrug surface-modified hGC33 on Wnt signaling pathway in hepatoma cells. Like free hGC33, nanodrug surface-modified hGC33 inhibited the proliferation of hepatoma cells not only by blocking Wnt-induced signal transduction in HepG2 and Huh-7 cells of expressing GPC3, but also by inhibiting Wnt3a-induced β-catenin and YAP signal transduction. Previous studies have shown that YAP expression is regulated by β-catenin at the transcriptional level of HCC [30, 31]. In this study, free hGC33 and nanodrug surface-modified hGC33 inhibited Wnt3a-induced YAP activity, indicating that Yap/TAZ released from β-catenin complex can also initiate classic Wnt signaling transduction [32, 2]. These results indicate that typical Wnt and YAP cross talk through a variety of mechanisms. Compared with hGC33-null-NP and hGC33, hGC33-SFB-NP had stronger anti-proliferation and anti-migration ability in vitro and in vivo. Thus, hGC33 not only determines the specificity of HCC cells, but also increases the inhibitory effect of SFB on the proliferation and migration of HCC cells by blocking the key signals related to tumor growth, such as Wnt/β-catenin and Wnt/YAP signaling pathway.
Conclusion
Antibody hGC33 to glypican-3, a membrane protein that is overexpressed on hepatocellular carcinoma cells, increased binding of sorafenib-loaded polyethylene glycol-b-PLGA polymer nanoparticles (hGC33-SFB-NP) to glypican-3 on the cancer cells. Administration of the antibody-modified nanoparticles synergistically inhibited Wnt-induced signal transduction and Ras/Raf/MAPK signaling pathway; hepatocellular carcinoma cells were arrested in G0/1 phase by down-regulation of cyclin D1 expression, thus attenuating cancer cell migration by inhibiting epithelial–mesenchymal transition. hGC33-SFB-NP inhibited the growth of liver cancer in vivo and improved the survival rate of tumor-bearing mice.
Доступность данных и материалов
Yes, all data have presented in the manuscript.
Сокращения
- AKT/PKB:
-
Protein kinase B
- c-MET:
-
HGFR:Hepatocyte growth factor receptor
- EE%:
-
Encapsulation efficiency %
- EMT:
-
Epithelial mesenchymal transition
- FTIR:
-
Инфракрасная спектроскопия с преобразованием Фурье
- 1 H NMR:
-
1 H-nuclear magnetic resonance spectroscopy
- HCC:
-
Hepatocellular carcinoma
- LC%:
-
Drug loading %
- MAPK:
-
Mitogen-activated protein kinases
- PDI:
-
Индекс полидисперсности
- PI3K:
-
Phosphoinositide 3-kinase
- pRAD51:
-
Phospho-RAD51
- SFB:
-
Sorafenib
- ТЕМ:
-
Просвечивающая электронная микроскопия
- YAP:
-
Yes-associated protein-1
Наноматериалы
- Наночастицы для терапии рака:текущий прогресс и проблемы
- Получение и магнитные свойства легированных кобальтом наночастиц шпинели FeMn2O4
- Наночастицы как насос оттока и ингибитор биопленки для омоложения бактерицидного действия обычных антибиот…
- Синергетические эффекты наночастиц Ag / BiV1-xMoxO4 с повышенной фотокаталитической активностью
- Влияние упругой жесткости и поверхностной адгезии на отскок наночастиц
- Последние достижения в синтетических методах и применении серебряных наноструктур
- Токсичность наночастиц CoFe2O4, покрытых ПЭГ, с лечебным эффектом куркумина
- Влияние кислотного пептизирующего агента на соотношение анатаз-рутил и фотокаталитические характеристики н…
- Золотые наночастицы круглой формы:влияние размера и концентрации частиц на рост корней Arabidopsis thaliana
- Зеленый синтез металлов и наночастиц оксидов металлов и их влияние на одноклеточную водоросль Chlamydomonas reinhardtii