


Роль ROS и остановки клеточного цикла в генотоксичности, вызванной наноструктурой ядра наностержня золота / серебряной оболочки
Аннотация
Чтобы понять генотоксичность, вызываемую в печени наночастицами серебра (AgNP) и ионами серебра, в этом исследовании использовались сконструированная наноструктура из золотых наностержней / серебряной оболочки (Au @ Ag NR) и гуманизированные гепатоцитные клетки HepaRG. Участие окислительного стресса и остановки клеточного цикла в повреждении ДНК и хромосом, вызванном 0,4–20 мкг / мл -1 Au @ Ag NR исследовали с помощью анализа комет, анализа γ-H2AX и теста на микроядер. Далее было проанализировано распределение NR Au @ Ag. Наши результаты показали, что как Ag + и Au @ Ag NR приводят к расщеплению ДНК и повреждению хромосом (кластогенности) в клетках HepaRG, и что Au @ Ag NR, сохраняемый в ядре, может дополнительно высвобождать Ag + , усугубляя повреждения, которые в основном вызваны остановкой клеточного цикла и образованием АФК. Результаты показывают корреляцию между внутриклеточным накоплением Ag + высвобождение ионов и потенциальная генотоксичность AgNP.
Введение
Наночастицы серебра (AgNP) размером от 1 до 100 нм могут проявлять широкий спектр антимикробных свойств, проникая в патогены и инактивируя внутреннюю сульфгидрильную группу их метаболических ферментов [1]. Они продемонстрировали мощный бактериостаз и бактерицидное действие на Escherichia Coli , Neisseria gonorrhoeae и Chlamydia trachomatis , и широко используются в качестве медицинских покрытий, предметов домашнего обихода [2], а также в качестве повязок для ран [3]. Неопровержимые доказательства показывают, что наночастицы способны проникать в ядро и вмешиваться в процесс синтеза и транскрипции ДНК [4]. В нашем предыдущем исследовании мы сообщили, что однократная внутривенная доза 5 мг / кг AgNPs может привести к значительному разрушению хромосом в клетках костного мозга крыс Sprague – Dawley [5]. Однократная внутрибрюшинная инъекция AgNP в дозе 10 мг / кг или выше индуцировала повреждение как ДНК, так и хромосом [6]. Flower et al. [7] предположили, что AgNP в дозах 50 и 100 мкг / мл -1 может вызвать повреждение ДНК в течение пяти минут после введения, что подчеркивает генотоксичность быстро высвобождающегося серебра (Ag). Учитывая риск чрезмерного воздействия, большое внимание уделяется исследованиям наногенотоксикологии или повреждения ДНК и канцерогенного потенциала созданных наноматериалов [8].
Основными механизмами генетических повреждений, вызванных AgNP, считаются избыточное производство реактивных окислительных форм, воспаление и нарушение клеточного цикла [9, 10]. Как предполагалось в предыдущих исследованиях, AgNPs могут либо напрямую взаимодействовать с ДНК посредством окислительного повреждения [11] и вмешиваться в интерфазу на уровне ДНК и митоз на хромосомном уровне, либо взаимодействовать с нуклеопротеином и аппаратом митотического веретена, нарушая контрольные точки клеточного цикла [ 12]. Однако независимо от того, связана ли генотоксичность, индуцированная AgNPs, частично с наночастицами [13, 14] или полностью с высвобожденным Ag + ионов до сих пор неясно [15, 16].
Изучение генотоксичности AgNP затруднено из-за нестабильного и непрерывного высвобождения серебра в тканях, что приводит к трудностям в локализации AgNP и дифференциации наноядра от Ag. Наша группа недавно разработала наноструктуру с ядром из золотых наностержней / серебряной оболочкой (Au @ Ag NR) для изучения токсичности, вызванной наночастицами [17]. Золотое ядро Au @ Ag NR физиологически врождено для ткани и может использоваться в качестве внутреннего стандарта для контроля высвобождения Ag + ионами из стержня путем мониторинга изменения отношения Ag / Au, измеренного с помощью масс-спектрометрии с индуктивно связанной плазмой (ICP-MS) [18]. С помощью этого метода можно определить различное происхождение токсичности. Предыдущие исследования показали, что выпущенный Ag + ионы из оболочки НР Au @ Ag вызывали окислительное повреждение почек и, в конечном итоге, приводили к морфологическим изменениям и нарушению фильтрационной функции клубочков [19]. Jiang et al. [20] предположили, что как специфическая активность частиц, так и высвобождение внутриклеточного иона серебра с помощью Au @ Ag NR вносят вклад в токсический ответ гранулезных клеток. Мы также приняли Au @ Ag NR в качестве модели для изучения потенциала генотоксичности AgNP in vivo и продемонстрировали, что кластогенность, а не мутагенность, является основной формой генотоксичности, вызванной как оболочкой Ag, так и высвобожденным Ag + ионы, в то время как их токсичность не различалась [21].
Печень является одним из основных органов, склонных к накоплению AgNP, и считается органом / тканью-мишенью для генотоксичности, вызванной AgNP. Наше предыдущее исследование показало, что некоторое количество серебра (8,26 ± 3,90 мкг / г) и золота (80,07 ± 64,72 мкг / г) оставалось в печени крыс SD через восемь недель после внутривенного введения одной дозы Au @ Ag NR [21 ]. В этом исследовании мы попытались определить роль остановки клеточного цикла и реактивного окислительного стресса на AgNP-индуцированные повреждения хромосом и ДНК с использованием Au @ Ag NR в клетках HepaRG, происходящих от гепатомы человека. Анализы генотоксичности, включая анализ комет, анализ γ-H2AX и тест микроядер, проводились параллельно с улавливателем окислительных радикалов, чтобы исследовать вклад активных форм кислорода (АФК) в повреждение ДНК / хромосом, в то время как апоптоз клеток, клеточный цикл и родственные белки были полны решимости изучить механизмы, с помощью которых AgNPs прерывают синтез и репликацию ДНК. Кроме того, внутриклеточное накопление и распределение Au @ Ag NR было исследовано путем сочетания масс-спектрометрии с индуктивно связанной плазмой (ICP-MS) и просвечивающей электронной микроскопии (TEM), чтобы дифференцировать роль наночастиц и высвобожденных ионов Ag.
Материалы и методы
Культивирование и лечение клеток
В этом исследовании использовали клеточную линию гепатомы человека HepaRG (Thermo Fisher Scientific). Клетки культивировали в среде RPMI 1640, содержащей 10% фетальной бычьей сыворотки (FBS, Australia Origin, Gibco) и 1% раствор пенициллин-стрептомицин-глутамин (Gibco), в увлажненной атмосфере с 5% CO 2 при 37 ° С. Клетки обрабатывали возрастающими концентрациями Au @ Ag NR в течение 24 или 72 часов, соответственно, и концентрации определяли в соответствии с IC 50 оценивается с помощью анализа жизнеспособности клеток. Чтобы исследовать роль АФК в генотоксичности, 1 мМ N -Ацетил-1-цистеин (NAC, Sigma-Aldrich) применяли за 1 час до обработки Au @ Ag NR.
Анализ роста / жизнеспособности клеток АТФ
Клетки высевали в 96-луночный планшет при плотности 5 × 10 3 . /Что ж. Через 24 часа инкубации среду аспирировали и клетки подвергали воздействию различных концентраций Au @ Ag NR в течение 24 или 48 часов соответственно. Был подготовлен широкий спектр концентраций, и четыре лунки на каждую обработку были выполнены за один период обработки. Цитотоксичность Au @ Ag NR изучали с помощью анализа аденозинтрифосфата (АТФ) (CellTiter-Glo® 2.0 Assay, Promega), который измеряет клеточную метаболическую активность путем количественного определения количества АТФ, важного параметра метаболизма в жизнеспособных клетках. Люминесцентные сигналы, отражающие количество жизнеспособных клеток, детектировали с помощью считывателя планшетов VICTOR Multilabel Plate Reader (2030-0050, PerkinElmer) и IC 50 значения были оценены как концентрация Au @ Ag NR для полумаксимальной жизнеспособности с помощью Prism 7 (GraphPad Prism 7, CA, USA). Коэффициент жизнеспособности рассчитывается по следующей формуле:
$$ {\ text {Жизнеспособность}} \, {\ text {Ratio}} \, \ left (\% \ right) ={\ text {RLU}} _ {{{\ text {sample}}}} / { \ text {RLU}} _ {{{\ text {vehicle}}}} \ times {1} 00 \% $$где RLU - относительная световая единица, представленная как среднее значение для четырех лунок, RLU vehicle представлены клетки, не обработанные наностержнями, и RLU sample представляли клетки, обработанные различными концентрациями Au @ Ag NR.
Определение концентрации серебра и золота в ячейках
Образцы клеток переваривали в азотной кислоте с использованием системы для микроволнового разложения. После разложения образцы были приготовлены из смеси, содержащей 1% азотную кислоту и соляную кислоту. Количество Ag и Au в растворах определяли методом ICP-MS (NexION300X, PerkinElmer). ПЭМ-анализ использовали для определения присутствия в клетке Au NR и Au @ Ag NR. Образцы клеток фиксировали в смеси 2,5% глутарового альдегида и 2% параформальдегида в течение 2 ч при 4 ° C. Осадки клеток фиксировали и трижды промывали фосфатным буфером (pH 7,4), а затем фиксировали в 1% тетроксиде осмия в течение 2 часов при 4 ° C. Затем образцы трижды промывали дистиллированной водой и обезвоживали в течение 15 мин в различных концентрациях этанола (50%, 70%, 90% и 100% этанола соответственно) один за другим. Затем на смолу наносили пропиленоксид в разведении 1:1 и 1:3 при 20–26 ° C в течение 2 часов. Полимеризацию проводили путем постепенного нагрева при 35 ° C в течение 16 часов, 45 ° C в течение 8 часов, 55 ° C в течение 14 часов и 65 ° C в течение 48 часов. Ультратонкие срезы окрашивали в течение 25 мин уранилацетатом и цитратом свинца и анализировали с помощью просвечивающего электронного микроскопа (H-7650, HITACHI, Япония).
Обычный и модифицированный анализ комет
Клетки высевали в 12-луночные планшеты плотностью 2 × 10 5 . / лунка или 3 × 10 5 / лунка для 24- или 72-часового лечения соответственно. Перекись водорода (H 2 О 2 ) в концентрации 200 мкмоль подвергали воздействию клеток в качестве положительного контроля в течение часа. Для каждого образца готовили две лунки как для стандартной обработки, так и для обработки формамидопиримидингликозилазой (Fpg). Обычный анализ комет проводился в щелочных условиях (pH> 13), как описано ранее [21]. Для лунок, обработанных Fpg, перед процедурой раскручивания ДНК применяли дополнительную обработку Fpg, и слайды погружали в ферментный буфер (0,1 М KCl, 0,5 мМ EDTA, 40 мМ HEPES, 0,2 мг / мл -1. BSA) трижды по 5 мин. Fpg (New England Biolabs, Inc., UK) разводили ферментным буфером в соотношении 1:50 000. Аликвоты по сто миллилитров разбавленного фермента добавляли к каждому гелю на предметных стеклах микроскопа и инкубировали во влажной камере при 37 ° C в течение 30 минут. Остальные шаги были такими же, как и при обычном лечении. Кометные анализы были выполнены в трех экземплярах. Не менее 50 клеток на образец независимо оценивали с помощью флуоресцентного микроскопа Nikon Eclipse 80i (Nikon, Токио, Япония), в то время как Komet 6.0 (Andor Technology, Белфаст, Великобритания) использовали для анализа среднего значения процентного содержания ДНК в хвосте и оливковом хвосте. момент (OTM) каждого образца.
Квалификация очагов γ-H2AX с помощью проточной цитометрии и скрининга с высоким содержанием
Для количественной оценки с использованием проточной цитометрии клетки высевали в 12-луночные планшеты при плотности 2 × 10 5 . / лунка или 3 × 10 5 на лунку для 24- или 72-часовой обработки, соответственно, тогда как для скринингового анализа с высоким содержанием клетки высевали в 96-луночные планшеты при плотности 6 × 10 3 / колодец или 1 × 10 4 / лунка для 24- или 72-часового лечения соответственно. В качестве положительного контроля 2 мкМ метилметансульфоната (MMS, Sigma-Aldrich) применяли параллельно с клетками в течение часа. Клетки промывали трис-буферным физиологическим раствором (TBS) и фиксировали 4% параформальдегидом в течение 15 мин при комнатной температуре. После промывания TBS клетки инкубировали с 50 мкл ледяного метанола в течение 30 мин при -20 ° C. Клетки дополнительно промывали в TBS три раза, и блокирующий реагент (TBS, содержащий 0,3% Triton X-100 и 10% козью сыворотку) применяли на 1 час. Первичное антитело (мышиное анти-фосфо-H2AX Ser139, Millipore) разбавляли до 1:200 блокирующим реагентом и инкубировали с клетками в течение ночи при 4 ° C. Затем планшет снова трижды промывали TBS, после чего добавляли вторичные антитела (козьи антимышиные антитела Alexa Fluor 488, Life Technologies), разбавленные блокирующим реагентом в соотношении 1:20. Образцы хранили в темноте при комнатной температуре в течение 1 ч и 2 мкг мл -1 (20 мкл / лунку) DAPI (Invitrogen) добавляли в каждую лунку. Флуоресценцию измеряли с помощью проточной цитометрии (FACSCalibur, BD Bioscience, Нью-Джерси, США) или системы анализа высокого содержания (Operetta CLS, PerkinElmer). Для анализа проточной цитометрии были проанализированы данные по меньшей мере от 10 000 клеток на группу, и эксперименты были выполнены в трех повторностях; для анализа с высоким содержанием были проанализированы 20 полей зрения в каждой лунке и не менее пяти лунок в каждой группе.
Анализ цитомного микроядра с блокировкой цитокинеза (CBMN-cyt)
CBMN-cyt выполняли в соответствии с процедурой, описанной Fenech et al. [22]. Клетки высевали в 12-луночные планшеты плотностью 2 × 10 5 . / лунка или 3 × 10 5 / лунка для 24- или 72-часового лечения соответственно. 0,2 мкг мл -1 Митомицин C (MMC, Tokyo Chemical Industry Co., Ltd. Япония) подвергали воздействию клеток в качестве положительного контроля в течение 24 часов. 3 мкг мл -1 цитохалазин B применяли после 24- или 72-часовой обработки, чтобы блокировать процесс цитокинеза, и клетки собирали через 40 часов. Образцы окрашивали 5% Гимза после гипотоничности предварительно нагретым 0,075 моль л -1 KCl и фиксация смесью метанола и уксусной кислоты 3:1. Были подготовлены три лунки для каждой группы, и было исследовано не менее 1000 двуядерных клеток на лунку.
Измерение MDA, общего содержания GSH и SOD
Клетки культивировали в 12-луночных планшетах при плотности 5 × 10 5 . / лунка или 3 × 10 5 / лунка для 24- или 72-часового лечения соответственно. Затем клетки собирали и трижды промывали фосфатным буферным солевым раствором (PBS). Количество малонового диальдегида (МДА) в гомогенатах клеток определяли с использованием метода на основе тиобарбитуровой кислоты (Nanjing Jiancheng Bio-Engineering Institute, Нанкин, Китай). Количества общего глутатиона (GSH) и супероксиддисмутазы (SOD) определяли с использованием наборов для количественного определения общего глутатиона и анализа SOD (Dojindo Molecular Technologies, Inc., Кумамото, Япония), соответственно. Оптическую плотность (ОП) каждой лунки измеряли с помощью считывателя планшетов VICTOR Multilabel (2030-0050, PerkinElmer).
Проточный цитометрический анализ клеточного цикла
Клетки культивировали в 6-луночных планшетах при плотности 1 × 10 6 . / колодец или 5 × 10 5 / лунку для 24- или 72-часовой обработки соответственно, а затем фиксировали 70% этанолом при 4 ° C в течение ночи. Образцы трижды промывали PBS и окрашивали буфером для окрашивания PI / Rnase (BD Biosciences) в течение 15 минут при комнатной температуре. Популяции клеток в фазе G0 / G1, S и G2 / M среди 20 000 клеток определяли с использованием областей с площадью FL2 по сравнению с шириной FL2. Анализ проводился с помощью проточной цитометрии (FACSCalibur, BD Bioscience, Нью-Джерси, США) и FlowJo (BD Bioscience), и эксперименты проводились в трех экземплярах.
Проточно-цитометрический анализ апоптоза клеток
Клетки культивировали в 6-луночных планшетах при плотности 1 × 10 6 . / колодец или 5 × 10 5 / лунка для 24- или 72-часового лечения соответственно. Затем их дважды промывали PBS и разбавляли 500 мкл 1 × связывающего буфера (FITC Annexin V Apoptosis Detection Kit I, BD Bioscience), чтобы довести суспензию до примерно 1 × 10 6 клеток / мл, а затем 100 мкл разведения смешивали с 5 мкл FITC Annexin V и 5 мкл PI. Образцы окрашивали при комнатной температуре в течение 15 минут, и по меньшей мере 10000 клеток анализировали для определения популяции клеток при раннем и позднем апоптозе с использованием областей с FL1H по сравнению с FL2H с использованием проточной цитометрии (FACSCalibur, BD Bioscience, NJ, USA) и FlowJo. (BD Bioscience). Эксперименты проводились в трех экземплярах.
Вестерн-блоттинг
Клетки культивировали в 75 см 2 колба плотностью 1 × 10 7 / хорошо и 6 × 10 6 / лунка для 24- и 72-часового лечения соответственно. Клетки лизировали лизисным буфером RIPA, содержащим ингибитор протеазы (PMSF), и концентрацию белков определяли с использованием набора для количественного определения белков BCA (Beyotime Biotechnology, Китай). Концентрации образцов регулировали с использованием буфера для лизиса RIPA перед денатурацией путем нагревания при 95 ° C в течение 3 минут. Пробы белков разделяли электрофорезом на 12% SDS-полиакриламидных гелях и переносили на нитроцеллюлозные мембраны (Millipore). Мембраны блокировали 5% обезжиренным молоком в течение 30 мин и инкубировали с первичным p53 (SC-137174, Санта-Крус), p21 (SC-6246, Санта-Крус) и β-актином (sc-47778, Санта-Крус) и вторичными антителами. козьи антимышиные IgG (H + L) -HRP (SE131, solabio) соответственно. Уровни экспрессии целевых белков в образцах визуализировали с помощью метода усиленной хемилюминесценции (ECL) и анализировали с помощью системы ImageJ (Национальные институты здравоохранения).
Статистический анализ
Данные представлены как среднее значение ± стандартная ошибка среднего. Односторонний дисперсионный анализ (ANOVA) был использован для проверки статистической значимости различий между группами отрицательного контроля и обработанными группами, за которым последовал тест множественного сравнения Даннета с использованием SPSS (версия 22, IBM, Армонк, Нью-Йорк, США), и данные были учтены. статистически значимо при P <0,05. Рисунки были подготовлены с помощью GraphPad Prism 7 для Windows (GraphPad Software, Ла-Хойя, Калифорния, США).
Результаты
Характеристика Au NR и Au @ Ag NR
Золотые наностержни (Au NR), ядра золотых наностержней и наноструктуры серебряной оболочки (Au @ Ag NR) были спроектированы, подготовлены и охарактеризованы, как описано ранее [21]. Вкратце, средние диаметр и длина составляют 15,0 ± 2,5 нм, 66,7 ± 2,5 нм для НП Au и 26,2 ± 3,0 нм, 72,7 ± 8,9 нм для НП Au @ Ag. Толщина оболочки Ag составляет около 5 нм. Дзета-потенциалы Au NR, покрытых PDDAC, и NR Au @ Ag, диспергированных в воде, составляли 37,7 ± 1,6 мВ и 52,5 ± 1,4 мВ, соответственно. Массовое соотношение Ag / Au полученного Au @ Ag NR составило 2.3. Результаты характеризации показаны на рис. 1.
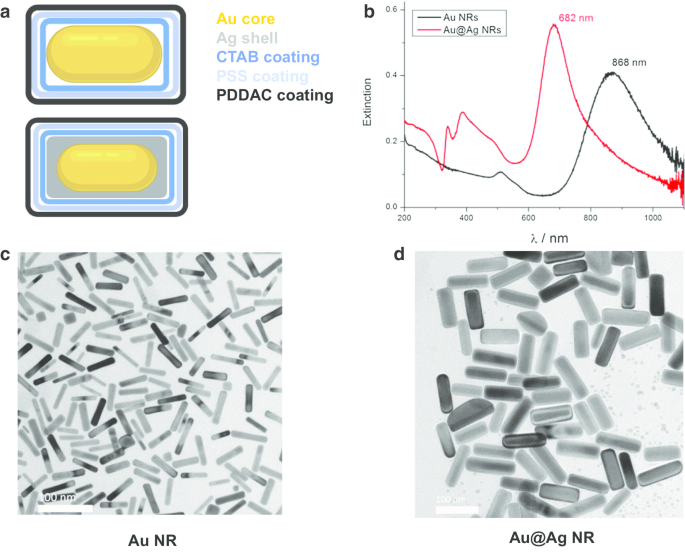
Характеристика Au NR и Au @ Ag NR. а Структурная схема Au NR и Au @ Ag NR; б UV – Vis – NIR спектры экстинкции Au NR и Au @ Ag NR, диспергированных в воде; c репрезентативные ПЭМ-изображения Au NR; г репрезентативные ПЭМ-изображения Au @Ag NR
Жизнеспособность клеток
Цитотоксичность Au @ Ag NR по отношению к клеткам HepaRG исследовали с помощью анализа жизнеспособности АТФ (таблица 1), и клетки подвергали действию Au @ Ag NR в течение 24 или 48 часов при концентрациях от 0,125 до 160 мкг · мл -1. . Au @ Ag NR индуцировал значительные цитотоксические эффекты как во времени, так и в зависимости от дозы после воздействия 24 и 48 часов, с% жизнеспособности IC 50 при 20 мкг мл -1 и 6 мкг мл -1 , оснащенный программой GraphPad Prism 7.0 соответственно. Принимая во внимание общую цитотоксичность, периоды лечения были скорректированы до 24 и 72 часов, а применяемые концентрации были определены как 0,8 мкг · мл -1 . , 4 мкг мл -1 и 20 мкг мл -1 . Кроме того, Au NR был включен в качестве инертного контроля, и содержание Au в группе AuNR было таким же, как 20 мкг мл -1 Au @ Ag NR, что составляет 16 мкг мл -1 . Напротив, предварительная обработка 1 мМ NAC была принята в группе Au @ Ag NR + NAC в качестве контроля реакции на окислительный стресс (концентрация Au @ Ag NR составляет 20 мкг / мл -1 ).
Распределение Au NR и Au @ Ag NR в клетках
Распределение содержания Au и Ag в клетках HepaRG анализировали с помощью ICP-MS. Как показано в таблицах 2 и 3, содержание Ag увеличивалось дозозависимым образом. Однако антиоксидант N -Ацетил-1-цистеин (NAC) в качестве акцептора свободных радикалов может ограничивать клеточное поглощение наночастиц, поскольку наблюдалось меньшее содержание Ag даже при той же концентрации Au @ Ag NR (20 мкг / мл -1 ) применялась в этой группе. Снижение соотношения Ag / Au с 24 до 72 часов указывает на непрерывное высвобождение Ag + из оболочки Au @ Ag NR. Кроме того, клеточное поглощение Ag намного больше, чем Au (Таблица 4). Кроме того, данные ПЭМ показали, что большая часть NR Au и Au @ Ag сохранялась в клетках в виде агломератов. Структуры наностержней были четко видны внутри клеток, подвергнутых воздействию Au NR или Au @ Ag NR без проникновения в ядро (рис. 2).
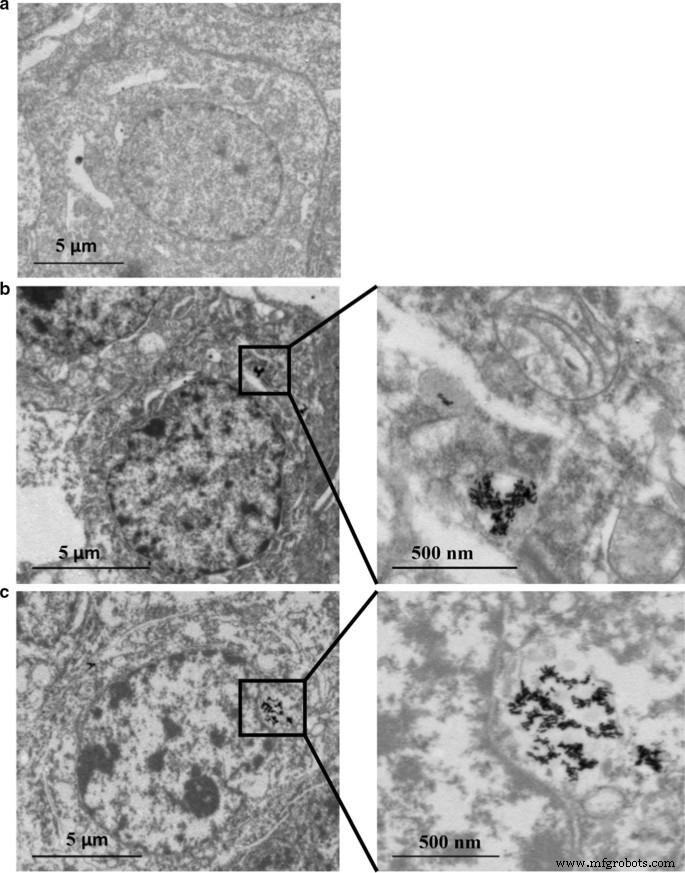
Интернализация Au NR и Au @ Ag NR:HepaRG с помощью ПЭМ при 80 кВ после 24 часов воздействия 16 мкг мл -1 Au NR и 20 мкг мл -1 Au @ Ag NR. а Управление автомобилем; б Au NR; c Au @ Ag NR
Повреждение ДНК
Повреждение ДНК, вызванное Au @ Ag NR, оценивалось как анализом комет, так и анализом γH2AX (рис. 3). Анализ комет показал, что от 0,8 до 20 мкг мл -1 Au @ Ag NR может привести к значительному повреждению ДНК. После 24- или 72-часового воздействия Au @ Ag NR как% хвостовой ДНК, так и ОТМ клеток увеличивались как во времени, так и в зависимости от концентрации. Кроме того, повреждение ДНК, связанное с индукцией окислительного стресса, наблюдалось в клетках, обработанных 20 мкг мл -1 Au @ Ag NR с помощью ферментно-модифицированного анализа комет (рис. 3а, б). Для оценки степени двухцепочечного разрыва, который представляет более высокую корреляцию с генезом рака, анализировали как γ-H2AX-положительные клетки, так и среднюю интенсивность флуоресценции в γ-H2AX-положительных клетках. После 24-часового воздействия Au @ Ag NR не было обнаружено различий между группами в γ-H2AX-положительных клетках. Однако 4 мкг мл -1 Группа Au @ Ag NR вызвала значительное увеличение после 72-часовой обработки. Значительное увеличение интенсивности флуоресценции наблюдалось во всех группах Au @ Ag NR через 72 часа по сравнению с контролем с носителем (рис. 3c – e, P <0,05).
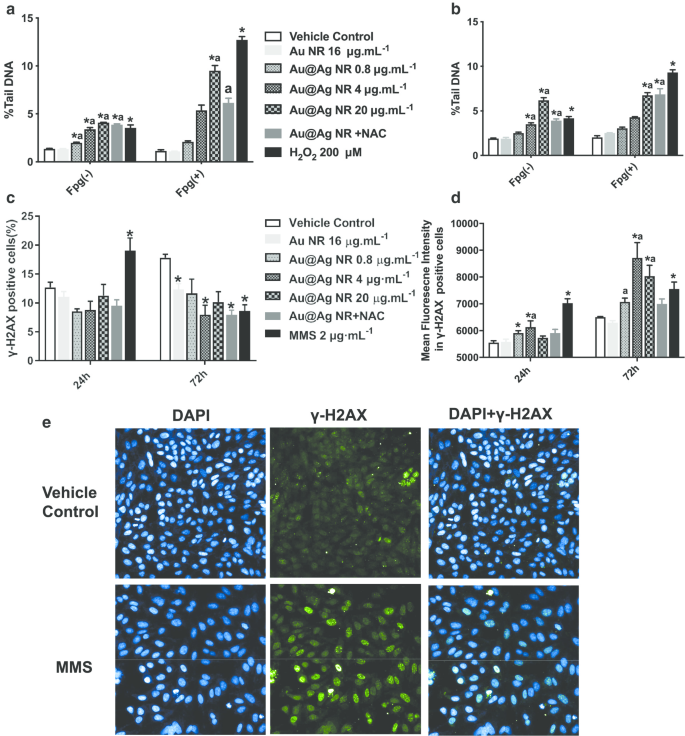
Повреждение ДНК, вызванное Au @ Ag NR. Клетки HepaGR подвергались воздействию Au @ Ag NR в различных концентрациях (от 0,8 до 20 мкг мл -1 ) в течение 24 и 72 часов соответственно. а Средний% хвостовой ДНК после воздействия Au @ Ag NR в течение 24 часов; б средний% хвостовой ДНК после воздействия Au @ Ag NR в течение 72 часов; c процент положительных клеток с очагами γ-H2AX, оцененный с помощью проточной цитометрии; г средняя интенсивность флуоресценции в клетках с фокусами γ-H2AX, оцененная с помощью иммунофлуоресцентного окрашивания. * P <0,05 по сравнению с контролем с носителем; а P <0,05 по сравнению с Au NR. 2 мкМ мл -1 MMS использовали в качестве положительного контроля
Хромосомное повреждение
Формирование микроядер является важным биомаркером для выявления хромосомного повреждения, которое является более серьезным повреждением генетического материала, чем разрыв ДНК. Соотношение двухъядерных клеток, содержащих микроядра, оценивали, как показано на фиг. 4c. Au @ Ag NR увеличивал образование микроядер в зависимости от концентрации. После 24-часового воздействия соотношение микроядер, наблюдаемое в клетках, обработанных 4 мкг мл -1 Au @ Ag NR и 20 мкг мл -1 NR Au @ Ag составлял 1,133 ± 0,145% и 1,567 ± 0,318%, соответственно, оба из которых были значительно выше, чем в контрольной группе носителя. После 72-часовой экспозиции соотношение микроядер в клетках, обработанных 4 мкг мл -1 NR Au @ Ag составлял 1,767 ± 0,233%, что было значительно выше, чем в контрольной группе с наполнителем; соотношение микроядер в клетках, обработанных 20 мкг мл -1 NR Au @ Ag составлял 2,167 ± 0,252%, что было значительно выше, чем в контрольной группе носителя и 16 мкг мл -1 . Группа Au NR (0,700 ± 0,153%). Напротив, не было обнаружено различий между клетками, обработанными 20 мкг мл -1 . Au @ Ag NR + NAC и контроль носителя, что предполагает участие ROS в разрушении хромосом, вызванном Au @ Ag NR.
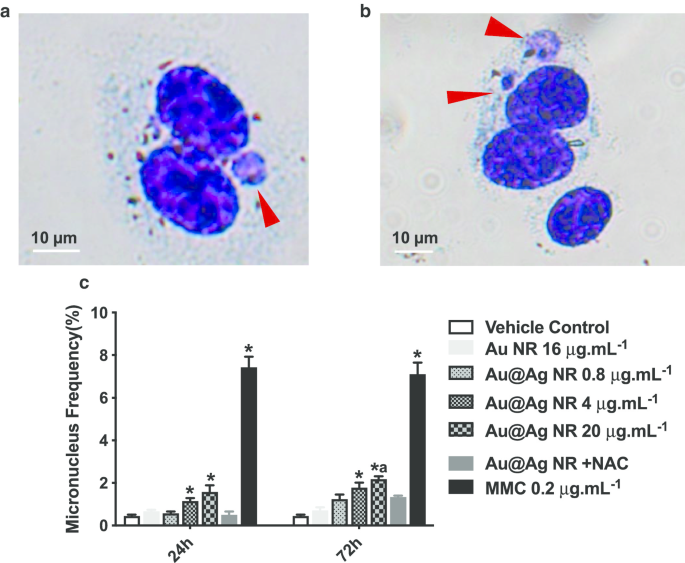
Повреждение хромосомы, вызванное Au @ Ag NR. Клетки HepaGR подвергались воздействию Au @ Ag NR в различных концентрациях от 0,8 мкг / мл -1 до 20 мкг мл -1 на 24 часа и 72 часа. а , b Репрезентативные изображения микронуклеуса (красная стрелка); c частота микронуклеусов (%). * P <0,05 по сравнению с контролем с носителем; а P <0,05 по сравнению с Au NR. 0,2 мкг мл -1 митомицин C использовали в качестве положительного контроля
Влияние Au @ Ag NR на образование ROS
Для дальнейшего изучения роли образования АФК в повреждениях ДНК и хромосом, вызванных Au @ Ag NR, были оценены уровни MDA, GSH и SOD. Значительное увеличение образования MDA ( P <0,05) наблюдалась после воздействия 20 мкг / мл -1 Au @ Ag NR как в течение 24, так и 72 ч (рис. 5а). Кроме того, уровни GSH и SOD в клетках, подвергнутых воздействию Au @ Ag NR, показали значительное снижение ( P <0,05) в зависимости от времени и концентрации. Эти результаты свидетельствуют о дисбалансе между окислением и антиокислением, вызванном воздействием Au @ Ag NR (рис. 5b, c).
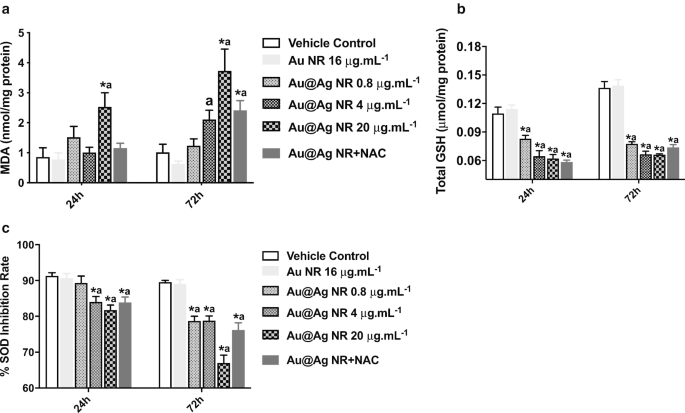
Влияние НК Au @ Ag на образование АФК. Клетки HepaGR подвергались воздействию Au @ Ag NR в различных концентрациях от 0,8 мкг / мл -1 до 20 мкг мл -1 на 24 часа и 72 часа. а Уровень MDA; б Уровень GSH; c Уровень SOD. * P <0,05 по сравнению с контролем с носителем; а P <0,05 по сравнению с Au NR
Влияние Au @ Ag NR на клеточный цикл и апоптоз
После 72-часового воздействия Au @ Ag NR увеличение количества клеток в фазе G2 / M наблюдалось в 4 мкг мл -1 Au @ Ag NR, 20 мкг мл -1 Группа Au @ Ag NR и Au @ Ag NR + NAC с долями 32,63% ± 1,77%, 32,267% ± 2,17% и 32,967% ± 4,25% соответственно (рис. 6a, b), что было значительно больше, чем в группе контрольная группа носителя (22,37% ± 0,92%). Между тем апоптоз клеток, индуцированный Au @ Ag NR, можно было наблюдать после 72-часового воздействия, а скорость позднего апоптоза клеток, обработанных 20 мкг мл -1 Au @ Ag NR и 20 мкг мл -1 Au @ Ag NR + NAC составило 78,90 ± 1,19% и 70,20 ± 4,50% соответственно (рис. 6в, г). Au @ Ag NR вызывает более поздний апоптоз, чем ранний, а лечение NAC может снизить скорость позднего апоптоза, вызванного Au @ Ag NR.
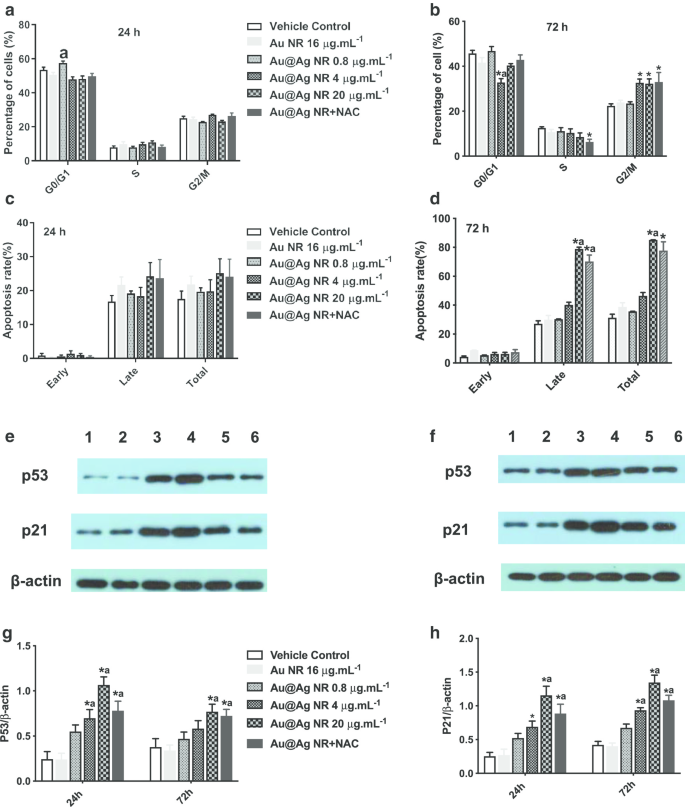
Влияние Au @ Ag NR на клеточный цикл и апоптоз. Влияние Au @ Ag NR на клеточный цикл ( a , b ) и апоптоз ( c , d ) после экспонирования в течение 24 и 72 часов соответственно; репрезентативные данные уровней экспрессии p53 и p21 в клетках HepaRG разных групп ( e , f Дорожка 1:управление транспортным средством; Дорожка 2:Au NR; Дорожка 3:Au @ Ag NR + NAC; Дорожка 4:Au @ Ag NR 20 мкг мл -1 ; Дорожка 5:Au @ Ag NR 4 мкг мл -1 ; Дорожка 6:Au @ Ag NR 0,8 мкг мл -1 ); усредненный относительный уровень экспрессии p53 и p21 по отношению к β-актину в разных группах был обобщен в ( g , f ). * P <0,05 по сравнению с контролем с носителем; а P <0,05 по сравнению с Au NR
Уровни экспрессии p21 и p53 определяли с помощью вестерн-блоттинга, и наблюдали аналогичную картину. Уровни экспрессии p53 и p21 в клетках, обработанных 4 мкг мл -1 и 20 мкг мл -1 NR Au @ Ag были заметно увеличены ( P <0,05) и значительно снизились в клетках, обработанных как 20 мкг мл -1 Au @ Ag NR и NAC ( P <0,05 по сравнению с 20 мкг мл -1 Группа Au @ Ag NR, рис. 6д – з). Известно, что белок p53 представляет собой ядро молекулы, опосредующей активацию контрольной точки G2 / M в ответ на повреждение ДНК, а p21 распознается как p53-зависимый ингибитор клеточного цикла. Таким образом, Au @ Ag NR может мешать репликации ДНК и препятствовать репарации ДНК за счет остановки клеточного цикла.
Обсуждение
В настоящее время роли выпущенного Ag + и AgNP в генерации генотоксичности далеко не ясны. Предыдущие исследования нашей группы [21] и других [13] показали, что в то время как Ag + является основным источником токсичности, наночастицы также могут быть очень токсичными. Например, AgNP могут вносить вклад в генотоксичность, индуцируя образование гидроксильных радикалов [13]. Кроме того, AgNP привел к более серьезному повреждению хромосом, окислительному стрессу и апоптозу по сравнению с Ag + только [23], предполагая, что могут быть задействованы разные пути. Мы использовали Au @ Ag NR в качестве модельного материала для понимания форм и распределения AgNP в клетках, а количество внутриклеточного Ag и Au определяли с помощью ICP-MS. Массовое соотношение Ag / Au полученного Au @ Ag NR составило 2.3. However, after a 24-h exposure, it sharply increased to 16.5 in the cells treated with Au@Ag NR, suggesting that large amount of Ag was released from the shell of Au@Ag NR within that period. When the exposure period of Au@Ag NR was extended to 72 h, the Au/Ag weight ratio was decreased to 1.7, indicating that the Ag + was released from the cell and the nanorod was the major form of Au@Ag NR in the cell at that stage. Therefore, it could be deduced that once the Au@Ag NR entered the cell, Ag + rapidly dissolved from its shell within 24 h and gradually released to the extracellular environment, while the Au@Ag NR itself retained in the cell for a longer period.
Oxidative stress is deemed as one of the most important toxicological mechanisms of nanoparticles [24]. N-acetylcysteine (NAC) is a thiol, a mucolytic agent and precursor of l-cysteine which reduced glutathione. NAC is also a source of sulfhydryl groups in cells and exerts the ROS scavenger activity by interacting with OH · и H 2 О 2 [25]. In this study, the GSH and SOD levels were significantly decreased after exposure to Au@Ag NR, while the MDA level increased in a concentration- and time-dependent manner, indicating that the Au@Ag NR introduced the oxidative stress in the cells.
The potentials of Ag and Au@Ag NR in interfering with the genetic materials were further investigated by a series of genotoxicity assays. It is noteworthy that co-culturing the NAC with Au@Ag NR could ameliorate the ROS formation, which in turn supports the participation of oxidative stress in the genotoxicity triggered by Au@Ag NR. In this study, comet and γ-H2AX assays were performed to confirm that Au@Ag NR could interact with DNA and induce certain DNA damage, and the repair endonuclease Fpg was included in the comet assay to identify the oxidative DNA damage [26]. The Fgp could recognize oxidized pyrimidines and remove oxidized purines, e.g., 8-hydroguanine, so as to create apurinic or apyrimidinic sites that could introduce gaps in the DNA strands. The oxidative stress-induced DNA breakage could be determined subsequently by another comet assay [27]. The further DNA breakage detected by the additional Fgp in the comet assay suggested that the Au@Ag NR could cause DNA damage. Mei et al. [28] observed that 5-nm-sized AgNPs induced oxidative lesion-specific DNA damage by employing the hOGG1, EndoIII and Fpg endonucleases in the comet assay. Ли и др. [29] also suggested that both PVP- and silica-coated AgNPs (15–100 nm and 10–80 nm, respectively) could lead to a significant increase in DNA breakage in mice hepatocytes in the presence of hOGG1and EndoIII. The formation of γ-H2AX foci, which represents an early cellular response to genotoxic stress, is the most sensitive and specific biomarker for detecting DSBs [30]. As demonstrated in this study, γ-H2AX foci in cells exposed to Au@Ag NR were markedly increased after 24 h, and a further increase could be observed after 72 h. The reduction in the 20 µg mL −1 group might be due to the cytotoxicity to the HepaRG cells at higher concentration. Similar results were observed for AgNPs with different coatings [31, 32]. Further, our results suggest that Au@Ag NR could induce chromosome damage in HepaRG cells, as the micronucleus rates were significantly increased. This is consistent with previous studies, where AgNPs-induced increased micronucleus rate was reported in HaCaT and TK6 cells [33]. In contrast, the addition of oxidative radical scavenger NAC could inhibit the formation of micronucleus induced by Au@Ag NR. Taken together, these data suggest the participation of oxidative stress in AgNP-introduced clastogenicity risk in vitro.
Previous studies have investigated the cell cycle arrest and cytotoxicity induced by AgNPs [33,34,35]. With prolonging the exposure time, the impact of AgNPs on cell cycle and apoptosis might be enhanced and in turn aggravate the cytotoxicity and genotoxicity. Usually, the cell cycle checkpoints (e.g., G2/M) were initiated by cells when experiencing DNA damage, and this mechanism serves to prevent the cell from entering mitosis (M phase). The G2/M cell cycle arrest indicates that an increasing percentage of cells is hindered in G2 phase for DNA repairing. Cells experiencing successful DNA repairing would further proceed to mitosis; however, for those with fatal damages, irreversible G2/M cell cycle arrest and cells apoptosis would take place [36]. We observed that Au@Ag NR could arrest the majority of HepaRG cells in G2/M phase, induce late cell apoptosis and increase the expression levels of p53 and p21, which are important proteins associated with the regulation of cell cycles [37]. As p53 could also induce apoptosis, when the DNA cannot be repaired properly [38], the p21 might indirectly participate in cell apoptosis by cell cycle arrest in a p53-dependent pathway via down-regulating the nuclear protein ICBP90 for DNA replication and cell cycle regulation [39]. Furthermore, apoptosis and a G2/M arrest induced by activation of the p53/p21 system have been reported in HepG2 cells following the administration of garlic extracts [40]. Thus, it could be inferred that the oxidative stress-triggered DNA/chromosome damages might facilitate the expression of p53 and p21, which subsequently induces cell cycle arrest. Extending the exposure period of Au@Ag NRs to the DNA/chromosome during replication may further aggravate the genotoxicity or apoptosis.
Conclusion
Genotoxicity induced by AgNPs may be attributed to the oxidative stress induced by the nanoparticles as well as the released ions [41]. This study employed Au@Ag NR as a model to determine the distribution and release behavior of Ag after the nanoparticles enter into the cells. Considering the disparate forms of Au@Ag NR in the cell, after its exposure the Ag + was rapidly dissolved from the silver shell. Ag + and Au@Ag NR could introduce cytotoxicity and genotoxicity (clastogenicity) in the cells, and the Au@Ag NR retained in the nucleus may further release Ag + to aggravate the damage, which are mainly caused by cell cycle arrest and ROS formation (summarized in Fig. 7). Collectively, these data reveal the correlation between the intracellular accumulation, Ag + release as well as the potential genotoxicity of AgNPs.

Schematic diagram of the possible mechanism of genotoxicity introduced by AgNP in vitro
Доступность данных и материалов
All data and materials are available without restriction.
Наноматериалы
- Разница между вольфрамовым серебряным сплавом и вольфрамово-медным сплавом
- Получение наночастиц mPEG-ICA, нагруженных ICA, и их применение в лечении LPS-индуцированного повреждения клеток H9c2…
- Изучение роли размера капель эмульсии и поверхностно-активного вещества в процессе изготовления мицеллярны…
- Влияние распределения наночастиц золота в TiO2 на оптические и электрические характеристики сенсибилизирован…
- Получение наноструктуры Au @ TiO2 - оболочка и ее применение для разложения и обнаружения метиленового синего
- Стимуляция роста клеток SH-SY5Y наночастицами золота, модифицированными 6-меркаптопурином и проникающим в нейро…
- Глубокие межфазные эффекты в наночастицах ядра / оболочки CoFe2O4 / Fe3O4 и Fe3O4 / CoFe2O4
- Сапонины Platycodon из Platycodi Radix (Platycodon grandiflorum) для зеленого синтеза наночастиц золота и серебра
- Конъюгированные с антителами золотые наностержни, модифицированные кремнеземом, для диагностики и фототерм…
- Оценка свойств доставки генов в антимикробные, апоптотические и раковые клетки для белковых наночастиц золо…