


Восстановленная микроРНК-133a-3p или истощенный PSAT1 сдерживает внутричерепную аневризму, вызванную повреждением эндотелиальных клеток, посредством подавления пути GSK3β / β-катенина
Аннотация
Неясно функциональная роль микроРНК-133a-3p (miR-133a-3p) при внутричерепной аневризме (IA). Следовательно, целью настоящего исследования было изучить регуляторную роль miR-133a-3p в регуляции ИА, вызванного повреждением эндотелия сосудов, посредством передачи сигналов фосфосерин-аминотрансферазы 1 (PSAT1) / гликоген-синтазы-киназы 3β (GSK3β) / β-катенина. путь. Нормальные ткани внутричерепных артериол и ткани IA были взяты у пациентов с травмой головного мозга и IA. Экспрессию miR-133a-3p, PSAT1, GSK3β и β-катенина в тканях определяли с помощью RT-qPCR и вестерн-блоттинга. Эндотелиальные клетки (ЭК) человеческого IA культивировали и обрабатывали миметиком miR-133a-3p и si-PSAT1 для определения их функций в миграции, апоптозе и пролиферации эндотелиальных клеток. Экспрессию miR-133a-3p, PSAT1, GSK3β, β-катенина, Ki-67, CyclinD1, Bax и Bcl-2 в ЭК тестировали с помощью RT-qPCR или вестерн-блоттинга. Более того, модель крыс IA была создана для обнаружения патологических изменений и экспрессии miR-133a-3p, PSAT1, GSK3β, β-катенина, VEGF и MMP-9 в тканях IA in vivo. Экспрессия miR-133a-3p была связана с количеством и размером IA. Экспрессия MiR-133a-3p снижена, а экспрессия PSAT1, GSK3β и β-катенина повышена в IA. Восстановленный miR-133a-3p и истощенный PSAT1 облегчили патологическое изменение; снижение экспрессии PSAT1, GSK3β и β-катенина в IA; подавлял апоптоз и увеличивал пролиферацию и миграцию ЭК, а также снижал экспрессию VEGF и MMP-9 в тканях IA in vivo. Наше исследование предполагает, что избыточная экспрессия miR-133a-3p или подавление PSAT1 сдерживает повреждение эндотелиальных клеток и способствует пролиферации эндотелиальных клеток посредством ингибирования пути GSK3β / β-катенина в IA. MiR-133a-3p может быть потенциальным кандидатом в маркер и терапевтической мишенью для IA.
Презентация гипотезы
В настоящем исследовании мы могли бы предположить, что ось miR-133a-3p / PSAT1 может влиять на IA, вызванное повреждением эндотелиальных клеток, посредством модуляции пути GSK3β / β-катенина.
Проверка гипотезы
Для проверки этой гипотезы мы собрали клинические образцы, эндотелиальные клетки (ЭК) ИА человека и установили модели ИА на крысах, чтобы выяснить функции miR-133a-3p и PSAT1 в процессе ИА.
Следствия гипотезы
Наше исследование подтверждает нашу гипотезу о том, что избыточная экспрессия miR-133a-3p или подавление PSAT1 сдерживают повреждение эндотелиальных клеток и ускоряют пролиферацию эндотелиальных клеток посредством ингибирования пути GSK3β / β-катенина в IA. Эти результаты позволяют по-новому взглянуть на новую таргетную терапию ИА.
Введение
Внутричерепная аневризма (ИА) представляет собой разновидность цереброваскулярного заболевания, которое проявляется нетрадиционным выпуклостью артерии в головном мозге, а также субарахноидальным кровоизлиянием (САК), вызванным разрывом ИА, сопровождающимся высокой летальностью и заболеваемостью [1]. Как деструктивное заболевание, патогенез ИА не выяснен [2]. IA - редкая семейная форма, но обычно считается результатом приобретенного сосудистого повреждения, вызванного гипертонией, курением и другими традиционными факторами риска [3]. Эндоваскулярная намотка или микрохирургическое клипирование использовались для предотвращения разрыва неразорвавшейся аневризмы в будущем у пациентов с высоким риском разрыва [4]. Хотя в хирургической операции ИА был достигнут значительный прогресс, у пациентов с ИА все еще наблюдается плохое послеоперационное восстановление [5]. Тяжелая ситуация с лечением ИА требует дальнейшего изучения механизма и поиска новой терапевтической стратегии.
МикроРНК (MiRNA) представляют собой класс некодирующих РНК, которые модулируют экспрессию генов-мишеней посредством ингибирования трансляции на посттранскрипционном уровне или опосредования деградации мРНК [6]. Было обнаружено, что miR-133a-3p в качестве ингибитора опухолей при некоторых злокачественных новообразованиях и сверхэкспрессия miR-133a-3p может подавлять рост клеток колоректального рака (CRC) [7]. В предыдущем исследовании утверждалось, что в свете блокирования опосредованного аутофагией глутаминолиза miR-133a-3p дополнительно подавляет метастазирование и рост рака желудка [8]. Также в исследовании было показано, что miR-133a-3p участвует в регуляции развития сердца и сердечной гипертрофии [9]. Исследование продемонстрировало, что повышающая регуляция miR-195-5p снижает ангиогенез и устойчивость к цисплатину при раке яичников посредством подавления фосфосерин-аминотрансферазы 1 (PSAT1) -зависимого пути передачи сигналов GSK3β / β-катенина [10]. В другом исследовании сообщается, что miR-365 подавляет клеточную инвазию и рост в плоскоклеточной карциноме пищевода (ESCC) посредством модуляции PSAT1 [11]. PSAT1 - это фермент, участвующий в биосинтезе серина; он первоначально был очищен из мозга овец и имеет высокие уровни во многих тканях [12]. Было документально подтверждено, что PSAT1, как обнаружено, опосредует прогрессирование клеточного цикла при раке груди посредством модуляции сигнального пути GSK3β / β-catenin [13]. Лю и др. также указывает на то, что PSAT1 влияет на развитие ESCC и прогнозирует плохую выживаемость; следовательно, он может быть многообещающей мишенью для противоопухолевой терапии [14]. Учитывая вышеупомянутый анализ, ожидалось, что это исследование внесет вклад в новый подход к функциональной роли оси miR-133a-3p / PSAT1 / GSK3β / β-катенина в IA.
Материалы и методы
Этика
Исследование было одобрено институциональным наблюдательным советом первой клинической больницы Тяньцзиньского университета традиционной китайской медицины. Все участники подписали документ об информированном согласии. Все эксперименты на животных согласованы с Руководством по уходу и использованию лабораторных животных международных комитетов.
Темы исследования
С января 2016 г. по март 2018 г. были отобраны случаи САК, вызванные ИА, леченные в нейрохирургии Первой клинической больницы Тяньцзиньского университета традиционной китайской медицины. Были собраны патологические образцы 75 случаев ИА, полученные с помощью микрохирургии, и отнесены к группе ИА, включая 29 мужчин и 46 женщин в возрасте 31–55 лет, средний возраст 44,98 ± 6,79 года. В качестве контрольной группы были выбраны пациенты с черепно-мозговой травмой, проходящие лечение в нейрохирургии одновременно в Первой клинической больнице Тяньцзиньского университета традиционной китайской медицины. Также 75 случаев здоровых тканей внутричерепных артериол были накоплены в результате травматической операции или внутренней декомпрессии, в том числе 43 мужчины и 32 женщины в возрасте 34–56 лет, средний возраст 48,14 ± 8,68 лет. Пациенты были исключены, если у них в анамнезе была гипертония, диабет или опухоли. Не было заметной разницы по полу и возрасту между группой IA и контрольной группой (обе P > 0,05).
Образцы обработки и сохранения
После хирургической резекции некоторые образцы в двух группах были зафиксированы формальдегидом, обезвожены градиентом спирта от низкой до высокой концентрации и залиты парафином. Затем образцы были нарезаны для окрашивания гематоксилин-эозином (HE) и иммуногистохимического окрашивания. Некоторые образцы быстро помещали в резервуары с жидким азотом, а затем переносили в криогенный холодильник с температурой - 80 ° C для обнаружения вестерн-блоттинга и количественной полимеразной цепной реакции с обратной транскрипцией (RT-qPCR). Некоторые образцы фиксировали глутаральдегидом для наблюдения под электронным микроскопом, а некоторые образцы использовали для выделения ЭК.
Наблюдение под электронным микроскопом
Образцы фиксировали 3% глутаровым альдегидом, а затем повторно фиксировали 1% тетроксидом осмия. Образцы обезвоживали ацетоном, заливали Epon812 и разрезали на полутонкие срезы толщиной 3 мкм. Наконец, образцы дважды окрашивали уранилацетатом и цитратом свинца и наблюдали с помощью просвечивающего электронного микроскопа H-600IV (Hitachi, Токио, Япония).
Окрашивание HE
Подготовленные парафиновые срезы запекали при 60 ° C в течение 30 мин. После завершения вышеуказанных этапов срезы ткани фиксировали в ксилоле, обезвоживали градиентом абсолютного спирта и очищали фосфатно-солевым буфером (PBS). Срезы ткани окрашивали гематоксилином, обрабатывали аммиаком в течение нескольких секунд, окрашивали эозином в течение 2 минут, обезвоживали и очищали. Затем на срезы ткани капали нейтральную жевательную резинку и закрывали покровным стеклом. Наконец, для наблюдения и записи был принят микроскоп (Nikon, Токио, Япония).
Иммуногистохимическое окрашивание
Набор для иммуногистохимии был произведен Zymed Laboratories (Сан-Франциско, Калифорния, США). Срезы парафина депарафинизировали и гидратировали, а срезы парафина погружали в раствор ксилола на 5 мин × 3 раза. Срезы помещали в 100% -ный абсолютный спирт на 3 мин × 2 раза, а затем вымачивали в 95–75% -ном спирте по очереди 3 мин. После депарафинизации срезы инкубировали с 3% перекисью водорода в течение 15 мин для устранения активности эндогенной пероксидазы. На срезы капали блокирующий раствор и инкубировали с нормальным рабочим раствором козьей сыворотки в течение 15 мин, затем зондировали первичным антителом против матриксной металлопротеазы (ММП) -9 (5 мкг / мл) и фактором роста эндотелия сосудов (VEGF) (1:250). , Abcam, Cambridge, MA, USA) (PBS для отрицательного контроля (NC)) и инкубировали в течение 1-2 часов. Срезы повторно зондировали рабочим раствором вторичных антител биотинилирования в течение 30–60 мин. В срезы добавляли рабочую жидкость стрептавидин / пероксидаза, меченную пероксидазой хрена, капали свежеприготовленный раствор диаминобензидина (DAB), окрашивали и блокировали. Изображение было обработано системой обработки изображений Nikon SPOT FlexTM. Площадь экспрессии белка MMP-9 и VEGF измеряли с помощью программного обеспечения для иммуногистохимического количественного анализа. В зоне скопления положительных клеток в каждом образце случайным образом были обнаружены пять мощных полей зрения, и среднее значение поглощения каждого экрана использовалось в качестве среднего значения для статистического анализа.
Изоляция и культура EC
ЭК выделяли из нормальных тканей внутричерепных артериол и тканей IA и культивировали. Ткань была разрезана на 3 мм 2 фрагменты и инкубировали в течение 25 мин в 0,1% коллагеназе B / 0,1% диспазе (Roche, Базель, Швейцария). Ткань отделяли, растирали в течение 2 минут пипеткой на 2 мл и фильтровали через фильтр 100 мкм (BD Biosciences, Нью-Джерси, США) для выделения ЭК. Суспензию клеток центрифугировали, а затем ресуспендировали в культуральной среде MV2, содержащей факторы роста и 20% фетальной бычьей сыворотки (PromoCell, Heidelberg, Германия). Затем клетки высевали на чашки, покрытые фибронектином (Sigma Aldrich Inc., Сент-Луис, Миссури, США), плотностью 10 4 клеток / см 2 (1 мкг / см 2 ) и выращивали в течение 1 дня с 5% CO 2 . На следующий день после посева клетки промывали PBS для удаления неприкрепленных клеток и помещали в свежую среду. При достижении 80–100% слияния культуры подвергали иммуносепарированию с помощью бусинок с покрытием Ulex europaeus Agglutinin I (UEA) (Vector Laboratories, Ltd., Питерборо, Великобритания) (Dynabeads M-450 Tosylactivated, Oxoid, Hampshire, UK), чтобы получить чистые ЭК. ЭК, связанные с покрытыми лектином гранулами, собирали с помощью концентратора магнитных частиц, в то время как несвязанные клетки удаляли путем двукратной промывки базальной средой. UEA-положительные клетки ресуспендировали в культуральной среде и высевали на покрытые фибронектином чашки для улучшения их адгезии и роста. Культуры стали сливаться в течение 4–6 дней.
Идентификация EC
ЭК были идентифицированы иммуноцитохимическим окрашиванием антителом к CD31 клеточной поверхности и антигеном, связанным с фактором FVII. Клетки дважды очищали PBS, фиксировали 4% параформальдегидом, инкубировали с 3% H 2 . О 2 в течение 10–15 мин для устранения эндогенной пероксидазной активности, а затем инкубировали с 0,1% Triton X-100 в течение 10 мин до перфорированных клеток. В клетки капали специфическое первичное антитело:фактор VII (1:200), CD31 (1:400, Roche, Базель, Швейцария) и инкубировали при 4 ° C в течение ночи. Затем в клетки капали иммуноглобулин G (1:50), меченный вторичным антителом к пероксидазе хрена. Клетки инкубировали при 37 ° C в течение 45 минут и проявляли с помощью DAB, избегая света, в течение 4 минут. Затем проявление цвета прекращали добавлением дистиллированной воды, и фотографию наблюдали под микроскопом. Клетки наблюдали под флуоресцентным инвертированным фазоразностным микроскопом, и положительные клетки и общее количество клеток подсчитывали случайным образом из 10 полей зрения. Скорость положительного окрашивания клеток =(количество положительных клеток / общее количество клеток) × 100%. Была создана соответствующая группа NC, и первичное антитело было заменено PBS, а остальные шаги были выполнены, как указано выше.
Группировка и трансфекция клеток
Чтобы изучить эффекты между miR-133a-3p и PSAT1 на ЭК IA, ЭК были сгруппированы в контрольную группу (нормальные сосудистые ЭК без какой-либо трансфекции), группу IA (сосудистые ЭК внутренних органов без какой-либо трансфекции), группу имитации NC (трансфицированные miR -133a-3p миметик NC), миметическая группа miR-133a-3p (трансфицированная миметиком miR-133a-3p), группа малой интерферирующей РНК (si) -NC (трансфицированная si-PSAT1 NC), группа si-PSAT1 (трансфицированная с si-PSAT1), и группа миметика miR-133a-3p + сверхэкспрессия (oe) -PSAT1 (трансфицированная миметиком miR-133a-3p и oe-PSAT1). Среди них миметик NC, миметик miR-133a-3p, si-PSAT1, si-NC и oe-PSAT1 были разработаны и составлены GenePharma Co., Ltd. (Шанхай, Китай). Трансфекцию проводили в строгом соответствии с инструкциями Lipofectamine TM Реагент для трансфекции 2000 (Thermo Fisher Scientific, MA, США).
Проточная цитометрия
Среду в культуральной чашке отбрасывали, и клетки дважды промывали PBS. Клетки отделяли 0,25% трипсином, центрифугировали при 800 об / мин в течение 5 минут и суспендировали с 1 × связывающим буфером, а плотность клеток доводили до 1 × 10 7 клеток / мл. Суспензию клеток (100 мкл) инкубировали с 5 мкл йодида пропидия (PI, 20 мкг / мл) и аннексином V-FITC в течение 20 мин, затем смешивали с 400 мкл 1 × связывающего буфера. Проточный цитометр (сортировщик клеток BD FACSArial I) использовали для обнаружения апоптоза клеток в течение 1 часа. Результаты показали, что левый нижний квадрант (Q4) на точечной карте показывает здоровые живые клетки (FITC - / PI - ), правый нижний квадрант (Q3) как ранние апоптотические клетки (FITC + / PI - ), а правый верхний квадрант (Q2) - клетки позднего апоптоза и апоптоза (FITC + / PI + ); Скорость апоптоза =процент раннего апоптоза (Q3) + процент позднего апоптоза (Q2).
Анализ 3- (4, 5-диметилтиазол-2-ил) -2,5-дифенилтетразолийбромида (МТТ)
Клетки отделяли трипсином для приготовления клеточной суспензии. Клетки подсчитывали под инвертированным микроскопом. Концентрацию клеток довели до 5 × 10 4 . клеток / мл. Клетки высевали в 96-луночный культуральный планшет. Через 48 ч клетки инкубировали с 20 мкл раствора МТТ в течение 4 ч. МТТ в каждой лунке растворяли в 150 мкл диметилсульфоксида. Величину оптической плотности (OD) ЭК измеряли при длине волны 570 нм. Скорость распространения ЭК рассчитывалась с учетом значения OD.
Скретч-тест
Клетки в каждой группе засевали в 24-луночный планшет с 2 × 10 5 ячеек / лунка. В каждой группе было установлено по три параллельные лунки. При достижении слияния примерно 90% плоскость роста клеток царапали стерилизованным одноразовым наконечником микропипетки на 1 мл; каждую лунку поцарапали один раз, и длина царапины и глубина каждой лунки были одинаковыми. После царапания плавающие клетки удаляли, культуральную среду заменяли свежей, и расстояние между царапинами наблюдали под микроскопом через 24 часа культивирования. Зона заживления царапины просчитывалась с помощью программного обеспечения National Instrument Vision Assistant 8.6. Миграция клеток =площадь заживления раны / площадь начальной царапины × 100%.
Экспериментальные животные и создание моделей крыс IA
Были отобраны 84 крысы Sprague-Dawley (SD) в возрасте 7 недель и массой от 180 до 200 г (Центр лабораторных животных Академии военно-медицинских наук, Пекин, Китай). Крыс содержали в экспериментальном центре на животных. Условия кормления контролировались при температуре 22–25 ° C и влажности 50–60% при естественном освещении. Всех крыс кормили в стандартных клетках по 4 крысы в клетке. Крыс кормили городской санитарной питьевой водой и кормом для крыс. Подушечки меняли каждые 3 дня, а клетку промывали и стерилизовали. Крысы IA моделировали согласно [15]. Разрыв аневризмы был идентифицирован, когда у крыс проявились следующие симптомы [16]:1, снижение активности в еде или питье, вызванное потерей веса (потеря веса около 10%) в течение 24 часов; 2 - сгибание туловища и передних конечностей при подъеме; 3, ходьба на боку в нормальной позе; 4, наклонившись на одну сторону в покое, без спонтанной активности. Крыс с этими симптомами умерщвляли через 3 месяца после операции. Ткани IA были получены во время операции и перфузированы PBS, а синий краситель, содержащий глутаминовую кислоту, был перфузирован в мозговую артерию.
Лечение и вмешательство крыс IA
Вышеупомянутые 84 крысы были случайным образом разделены на 7 групп по 12 крыс в каждой группе. Методы лечения:нормальная группа (моделирование не проводилось); Группа IA (стереотаксическая инъекция 100 мкл смеси PBS и липофектамина 2000); группа миметиков NC (стереотаксическая инъекция 100 мкл смеси miR-133a-3p миметика NC и липофектамина 2000); группа миметика miR-133a-3p (стереотаксическая инъекция 100 мкл смеси миметика miR-133a-3p и липофектамина 2000); группа si-NC (стереотаксическая инъекция 100 мкл смеси si-PSAT1 NC и липофектамина 2000); группа si-PSAT1 (стереотаксическая инъекция 100 мкл смеси si-PSAT1 и липофектамина 2000); и группа миметика miR-133a-3p + oe-PSAT1 (стереотаксическая инъекция 100 мкл смеси миметика miR-133a-3p и oe-PSAT1 и липофектамина 2000). Все вышеуказанные инъекции выполнялись один раз в день, и эти крысы выращивались в лаборатории на животных, свободной от конкретных патогенов (SPF), в течение 12 недель. Через 12 недель крыс в каждой группе анестезировали и открывали грудную полость, как описано выше. Из левого желудочка, интубированного в аорту, кровь выделялась путем разрезания полой полости. В то же время через проток перфузировали 30 мл физиологического раствора, содержащего гепарин натрия (37 ° C), а затем через проток медленно вводили в мозг 10% полиформальдегид / 0,1 М фосфатный буфер (pH 7,4). После восстановления перфузии мозг вскрыли. Артериальное кольцо у основания черепа было отделено и удалено под хирургическим микроскопом, под микроскопом наблюдались изменения аневризм и изучены патологические характеристики. Мимик NC, миметик miR-133a-3p, si-NC, si-PSAT1 и oe-PSAT1 были составлены Shanghai Sangon Biotechnology Co., Ltd. (Шанхай, Китай).
Обнаружение гемодинамики
Скорость кровотока в конце левой общей сонной артерии крыс исследовали до 3 дней операции и 12 недель после интервенционного лечения. Метод заключался в следующем:крыс помещали в раму анестезиологического аппарата для ингаляции и регулировали параметры потока. После того, как крысы стали дышать стабильно и не было очевидной реакции при прикосновении к хвосту крыс, крыс фиксировали на экспериментальном операционном столе с помощью резиновой ленты. Волосы на шее крысы сбривали электробритвой. Детектор цветного доплеровского ультразвука был включен, скорость кровотока на конце левой общей сонной артерии была измерена, и данные были записаны после того, как зонд был смазан соответствующим связывающим агентом. После измерения крыс осторожно поместили обратно в клетку, чтобы дыхательные пути не были заблокированы, пока крысы не проснулись после анестезии.
RT-qPCR
Суммарную РНК выделяли на основе набора для экстракции простой общей РНК (TIANGEN Biotechnology Co., Ltd., Пекин, Китай). Высокое качество РНК было подтверждено ультрафиолетовым анализом и электрофорезом денатурации формальдегида, и РНК была обратно транскриптирована в комплементарную ДНК с помощью набора реагентов PrimeScript RT. Реакцию ПЦР проводили с помощью SYBR Permix Ex Taq II . (Такара, Далянь, Ляонин, Китай). Праймеры для ПЦР были разработаны и составлены компанией Beijing ComWin Biotech Co., Ltd. (Пекин, Китай) (таблица 1). U6 был выбран в качестве внутреннего параметра для miR-133a-3p, PSAT1, GSK3β, β-катенина, Bax, Bcl-2, Ki-67 и CyclinD1 с глицеральдегидфосфатдегидрогеназой (GAPDH) в качестве внутреннего параметра. Данные были измерены с помощью 2 −ΔΔCt .
Вестерн-блоттинг
Полные белки экстрагировали из клеток и тканей, и образцы белков количественно определяли с помощью набора для анализа белков бицинхониновой кислоты (Институт биотехнологии Beyotime, Шанхай, Китай). Образцы смешивали с 1/4 объема 5-кратного буфера для образцов и кипятили в течение 5 мин. Для электрофореза были выбраны 10% разделительный гель и 5% концентрированный гель. Мембрану вылупляли в 5% сухом обезжиренном молоке в течение 60 мин. К мембране добавляли первичные антитела PSAT1 (1:500), GSK3β (1:500), β-катенин (1:5000), Bax (1:1000), Bcl-2 (1:1000), CyclinD1 (1:200), Ki-67 (1:5000), MMP-9 (1 мкг / мл), VEGF (1:1000) (все от Abcam, Кембридж, Массачусетс, США). Затем через мембрану добавляли вторичное антитело (1:2000) в течение 60 мин. Мембрану погружали в раствор для реакции электрохемилюминесценции (Beyotime Institute of Biotechnology, Шанхай, Китай) на 1 мин, затем закрывали пищевой пленкой после удаления жидкости. Мембрану подвергли рентгеновскому облучению, и результат наблюдали после проявления и фиксации. GAPDH (1:10000, Abcam) использовали в качестве контроля загрузки, а изображение белка анализировали с помощью программного обеспечения ImageJ2x.
Анализ двойного люциферазного репортерного гена
Целевые отношения между miR-133a-3p и PSAT1 и сайт связывания между miR-133a-3p и 3'-нетранслируемой областью PSAT1 (3'UTR) были спрогнозированы сайтом биоинформатики (https://cm.jefferson.edu/rna22). / Предварительно вычислено /). Последовательность промоторной области 3'UTR PSAT1, содержащей сайт связывания miR-133a-3p, была амплифицирована и клонирована в плазмиду pGL3-основной люциферазы (Takara Bio Inc., Otsu, Shiga, Japan) для конструирования плазмиды дикого типа (WT) ( PSAT1-WT) PSAT1 3'UTR, тогда как мутантная (MUT) рекомбинантная плазмида PSAT1-MUT была получена путем мутации сайта связывания miR-133a-3p на PSAT1-WT с помощью набора для точечной мутации (Takara Bio Inc., Otsu, Shiga , Япония). ЭК сосудов в фазе логарифмического роста высевали в 96-луночный планшет. Когда слияние достигает примерно 70%, плазмиды PSAT1-WT и PSAT1-MUT были смешаны с мимическими плазмидами NC и miR-133a-3p с помощью Lipofectamine 2000 и котрансфицированы в сосудистые ЭК. Клетки собирали и лизировали через 48 часов после трансфекции, и активность люциферазы проверяли с помощью набора для определения люциферазы (Promega Corporation, Мэдисон, Висконсин, США).
Статистический анализ
Все данные были эксплицированы с помощью программного обеспечения SPSS 21.0 (IBM Corp., Армонк, Нью-Йорк, США). Данные подсчета обозначались как процент или процент, а анализ проводился с помощью критерия хи-квадрат или критерия Фишера. Данные измерений, подвергнутые нормальному распределению, были представлены как среднее ± стандартное отклонение. Сравнение между двумя группами было проведено t тест, тогда как сравнение между несколькими группами было проанализировано с помощью одностороннего дисперсионного анализа (ANOVA) с последующим апостериорным тестом Тьюки. А P значение <0,05 считалось значимым.
Результаты
Общие данные пациентов с ИА
Как показано в таблице 2, сравнивались общие данные группы IA и контрольной группы. Конкретная информация указана в Таблице 2.
Выражение miR-133a-3p связано с количеством и размером IA
Анализируя взаимосвязь между экспрессией miR-133a-3p и клинико-патологическими особенностями IA, в таблице 3 было подробно показано, что в свете средней относительной экспрессии miR-133a-3p в IA, 75 случаев IA были разделены на два группы:группа высокой экспрессии miR-133a-3p ( n =47) и группа с низкой экспрессией miR-133a-3p ( n =28). Взаимосвязь между miR-133a-3p и различными клинико-патологическими параметрами была статистически проанализирована с помощью критерия хи-квадрат или критерия Фишера. Результаты показали, что экспрессия miR-133a-3p не связана с возрастом, полом, формой и положением аневризмы (все P > 0,05), но связано с количеством и размером аневризмы (оба P <0,05).
Экспрессия MiR-133a-3p отклоняется, а экспрессия PSAT1, GSK3β и β-катенина повышается в тканях IA
Экспрессию miR-133a-3p в IA определяли с помощью RT-qPCR, и результаты показали, что по отношению к нормальным тканям внутричерепных артериол (контрольная группа) экспрессия miR-133a-3p была снижена в тканях IA ( Группа IA) ( P <0,05) (рис. 1а). Анализ RT-qPCR и вестерн-блоттинга показал, что экспрессия PSAT1, GSK3β и β-катенина была повышена в тканях IA по сравнению с нормальными тканями внутричерепных артериол (все P <0,05) (рис. 1а – в).
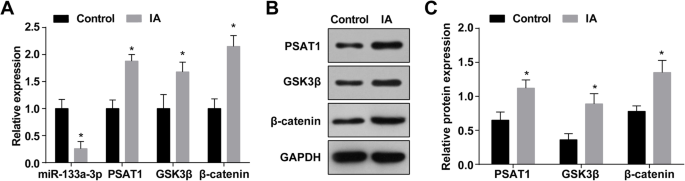
Экспрессия MiR-133a-3p снижена, а экспрессия PSAT1, GSK3β и β-катенина повышена в тканях IA. а Экспрессия miR-133a-3p, PSAT1, GSK3β и β-катенина в группе IA и нормальной группе. б Белковая полоса экспрессии PSAT1, GSK3β и β-катенина. c Экспрессия белков PSAT1, GSK3β и β-катенина в группе IA и нормальной группе методом вестерн-блоттинга. нет =75, * P <0,05 по сравнению с контрольной группой. Данные измерений были представлены как среднее значение ± стандартное отклонение, а сравнения между двумя группами проводились с помощью независимой выборки t тест
Патологические изменения аневризмы и экспрессии MMP-9 и VEGF в тканях IA
Непосредственно наблюдая за нормальной тканью внутричерепных артериол и тканями IA, было показано, что в контрольной группе сосуды в тканях артерий были ярко-красными, а в просвете не было обнаружено явных атеросклеротических бляшек и боковых тромбов. Опухоль тканей аневризмы группы IA в основном была коричневой или темно-красной, на вид была скротеформной или веретенообразной, а текстура была в основном жесткой. Когда опухоль была разрезана, на опухолевой стенке некоторых образцов опухоли появлялись белые или темно-красные атеросклеротические бляшки, которые имели плоскую, круглую или овальную форму. В некоторых образцах опухоли в полости опухоли имелся настенный тромб, и текстура тромба была мягкой. Толщина стенки опухоли постепенно уменьшалась от шейки опухоли, некоторые из которых имели только тонкую волокнистую мембрану на вершине опухоли, а некоторые из них даже разорвались. Расщелина разорванной аневризмы располагалась на вершине опухоли или рядом с ней.
Окрашивание HE показало, что под световым микроскопом толщина стенки нормальных тканей внутричерепной артериолы была однородной; анатомическое строение внутреннего, среднего и внешнего слоев четкое и неповрежденное; морфология клеток в каждом слое была нормальной; сарколемма соседних клеток часто формируется тесно; и воспалительные клетки стенки были редкими. В группе ИА локальные выпячивания, образовавшиеся в дистальной боковой сосудистой полости на вершине бифуркации внутричерепной артерии в стенке аневризмы, стали тупыми и меньшего размера, а локальные ЭК были утрачены. Небольшое количество образцов показало миграцию из слоя гладкомышечных клеток в слой интимы и миогенную пролиферацию клеток интимы. ЭК уменьшились или даже исчезли. Слой эндотелиальных клеток состоит из гиперпластических миоинтимальных клеток и линейно расположенных ЭК или из апоптотических ЭК и клеток крови, прикрепленных к просвету. Его вакуоль дегенерировала и прерывалась. Некоторые из них отслоились вместе с базальной мембраной, а коллагеновые волокна интимы были увеличены. Изменен атеросклероз, стенка артериолы явно тоньше и заполнена большим количеством соединительных лестничных тканей. Инфильтрация и частичная диффузия воспалительных клеток наблюдались во всех слоях, в основном в средней и внешней мембранах. В некоторых клетках наблюдались отложения кристаллов липидов и холестерина. Некоторые стенки опухоли были полностью или локально истончены и расширены наружу (рис. 2а, б).
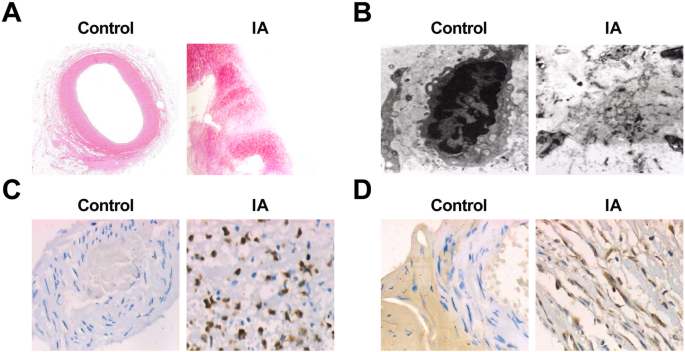
Патологические изменения аневризмы и экспрессия MMP-9 и VEGF в IA. а Нормальные срезы ткани внутричерепных артериол в контрольной группе при окрашивании HE (× 10). б Срезы тканей IA, окрашенные HE (× 10). c Ультраструктура нормальных тканей внутричерепных артериол в контрольной группе под электронным микроскопом (× 10 000). г Ультраструктура тканей IA под электронным микроскопом (× 10 000). е Экспрессия MMP-9 в контрольной группе и группе IA путем иммуногистохимического окрашивания (× 200). е Экспрессия VEGF в контрольной группе и группе IA методом иммуногистохимического окрашивания (× 200)
Срезы нормальных тканей внутричерепных артериол и тканей IA наблюдались под электронным микроскопом, и он показал, что в нормальных тканях внутричерепных артериол можно было четко увидеть матричные волокна стенки сосудов головного мозга, и не было эндотелиального повреждения, клеточного пикноза. , или вырождение. В тканях IA наблюдались явное повреждение эндотелиальных клеток, пикноз клеток или дегенерация вакуолей, количество средних гладкомышечных клеток уменьшалось, появлялся пикноз ядра, наблюдалась агрегация хроматина и апоптотические тельца. В некоторых клетках наблюдалось набухание митохондрий и исчезновение нормальной внутренней структуры. Внеклеточный матрикс, образующий цитоскелет, был размытым и имел аморфный хлопьевидный белок. В недостающих частях ячеек было много фрагментов (рис. 2в, г).
Иммуногистохимическое окрашивание использовали для тестирования экспрессии MMP-9 и VEGF, и результаты показали, что в 75 случаях контрольной группы не было экспрессии MMP-9 и VEGF. Было 60 случаев положительной экспрессии MMP-9 в 75 случаях образцов IA. Положительная экспрессия MMP-9 проявлялась во внутренней и внешней мембранах стенки IA, но экспрессия не была однородной. Положительная экспрессия в основном характеризовалась коричневато-желтой цитоплазмой. Положительная экспрессия VEGF составила 66 случаев в 75 случаях образцов IA. В стенке IA была высокая положительная экспрессия в средней и наружной мембранах и низкая положительная экспрессия в интиме. Положительная экспрессия также в основном характеризовалась коричневато-желтой цитоплазмой (рис. 2e, f). Экспрессия MMP-9 и VEGF в двух группах показана в таблице 4.
Идентификация сосудистых ЭК
Экспрессию фактора VIII и CD31 в ЭК анализировали иммуногистохимическим окрашиванием. Результаты показали, что ЭК сосудов положительно реагировали на фактор VIII и антитела к антигену CD31, и положительная частота составила 95%. Кроме того, в цитоплазме было большое количество коричневых частиц, и пятый пассаж клеток с коричневым окрашиванием был значительно выше, чем первичный пассаж клеток (рис. 3a, b).
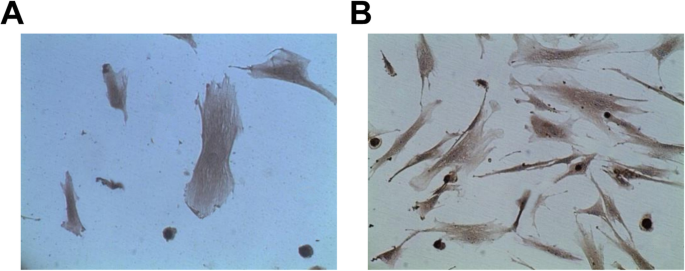
ЭК сосудов положительно реагируют на антитела к антигенам FVIII и CD31. а Идентификация ЭК по CD31. б Идентификация ЭК по FVII
Повышающая регуляция miR-133a-3p и понижающая регуляция PSAT1 подавляют апоптоз и ускоряют пролиферацию и миграцию ЭК в IA
Проточная цитометрия, RT-qPCR и вестерн-блоттинг-анализ были приняты для наблюдения за апоптозом и экспрессией Bax и Bcl-2 в ЭК IA после обработки миметиком miR-133a-3p или si-PSAT1. Было указано, что по сравнению с контрольной группой скорость апоптоза клеток и экспрессия Bax были повышены в группе IA, а экспрессия Bcl-2 была снижена (все P <0,05). Апоптоз клеток и экспрессия Bax и Bcl-2 в группе IA, группе имитации NC и группе si-NC не претерпели значительных изменений (все P > 0,05). По сравнению с группой si-NC и группой миметиков NC, скорость апоптоза клеток в группе si-PSAT1 и группе миметиков miR-133a-3p была подавлена, экспрессия Bax снижалась, а экспрессия Bcl-2 повышалась ( все P <0,05). По сравнению с группой миметика miR-133a-3p скорость апоптоза и экспрессия Bax были увеличены, а экспрессия Bcl-2 была снижена в группе миметика miR-133a-3p + oe-PSAT1 (все P <0,05) (рис. 4a – e).
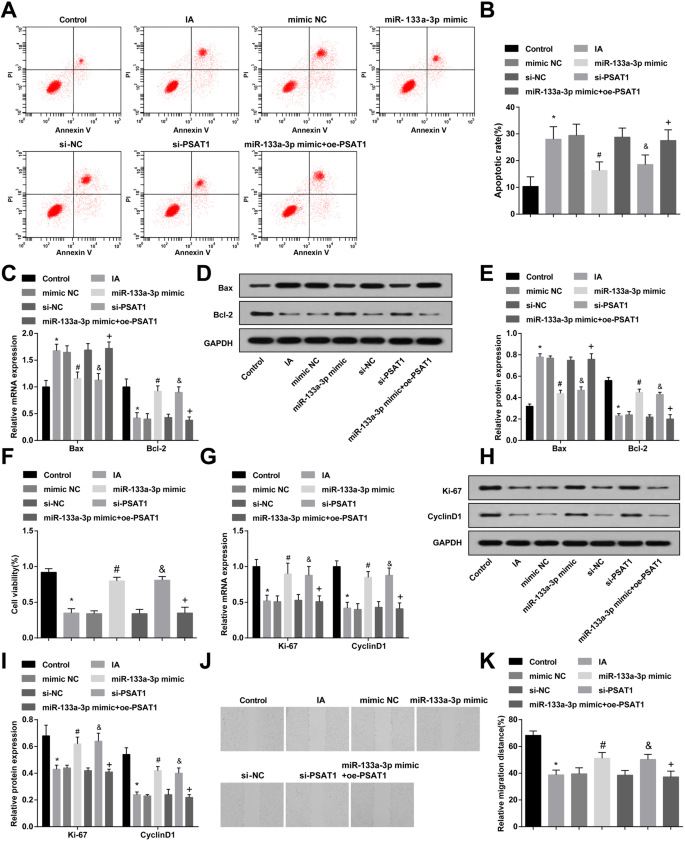
Высокоэкспрессируемый miR-133a-3p и низкоэкспрессированный PSAT1 ингибируют апоптоз и способствуют пролиферации и миграции IA ECs. а Обнаружение апоптоза ЭК методом проточной цитометрии. б Определение скорости апоптоза ЭК в каждой группе. c Экспрессия Bax и Bcl-2 в ЭК, обнаруженная с помощью RT-qPCR. г Белковая полоса экспрессии Bax и Bcl-2. е Экспрессия белков Bax и Bcl-2 в ЭК, обнаруженная с помощью вестерн-блоттинга. е МТТ-анализ использовали для определения пролиферативной активности ЭК в каждой группе. г RT-qPCR использовали для обнаружения экспрессии Ki-67 и CyclinD1 в каждой группе EC. ч Белковая полоса экспрессии Ki-67 и CyclinD1. я Экспрессия белков Ki-67 и CyclinD1 в ЭК, обнаруженная с помощью вестерн-блоттинга. j Обнаружение миграции ЭК в каждой группе с помощью скретч-теста. к Статистические результаты миграции эндотелиальных клеток в каждой группе. N =3, * P <0,05 по сравнению с контрольной группой. # P <0,05 по сравнению с группой мимических NC. & P <0,05 по сравнению с группой si-NC. + P <0,05 по сравнению с группой миметиков miR-133a-3p. Данные измерений были представлены как среднее ± стандартное отклонение; данные были оценены с помощью одностороннего дисперсионного анализа с последующим апостериорным тестом Тьюки
МТТ-анализ, RT-qPCR и вестерн-блот-анализ использовали для наблюдения за пролиферацией и экспрессией Ki-67 и CyclinD1 в ЭК IA после обработки миметиком miR-133a-3p или si-PSAT1. Было показано, что в отличие от контрольной группы, активность пролиферации и экспрессия Ki-67 и CyclinD1 были снижены в группе IA (все P <0,05). Не было значительных различий в активности пролиферации и экспрессии Ki-67 и CyclinD1 в группе IA, группе имитации NC и группе si-NC (все P > 0,05). Что касается группы si-NC и группы миметиков NC, активность пролиферации и экспрессия Ki-67 и CyclinD1 были повышены в группе si-PSAT1 и группе миметиков miR-133a-3p (все P <0,05). По сравнению с группой миметиков miR-133a-3p, активность пролиферации и экспрессия Ki-67 и CyclinD1 были снижены в группе миметиков miR-133a-3p + oe-PSAT1 (все P <0,05) (рис. 4f, i).
Миграцию ЭК в каждой группе после обработки миметиком miR-133a-3p или si-PSAT1 в течение 24 часов наблюдали с помощью скретч-теста. Выявлено, что миграция клеток в группе IA ингибировалась по сравнению с таковой в контрольной группе ( P <0,05). Не было заметных изменений в миграции клеток в группе IA, группе si-NC и мимической группе NC (все P > 0,05). По сравнению с группой si-NC и группой миметиков NC миграция клеток в группе si-PSAT1 и группе миметиков miR-133a-3p была увеличена (обе P <0,05). По сравнению с группой миметиков miR-133a-3p миграция клеток была снижена в группе миметиков miR-133a-3p + oe-PSAT1 ( P <0,05) (рис. 4j, k).
Восстановленный miR-133a-3p и истощенный PSAT1 снижают экспрессию PSAT1, GSK3β и β-катенина в ЭК IA
RT-qPCR использовали для обнаружения экспрессии miR-133a-3p в ЭК IA; Было установлено, что по сравнению с контрольной группой экспрессия miR-133a-3p в группе IA была снижена ( P <0,05). Экспрессия miR-133a-3p в группе IA, группе имитации NC и группе si-NC существенно не изменилась ( P > 0,05). Экспрессия MiR-133a-3p в мимической группе miR-133a-3p была усилена по сравнению с таковой в мимической группе NC ( P <0,05). В отличие от группы si-NC, не было явных изменений в экспрессии miR-133a-3p в группе si-PSAT1 ( P > 0,05). По сравнению с группой миметиков miR-133a-3p, экспрессия miR-133a-3p не показала значительных различий в группе миметиков miR-133a-3p + oe-PSAT1 ( P > 0,05) (рис. 5а).
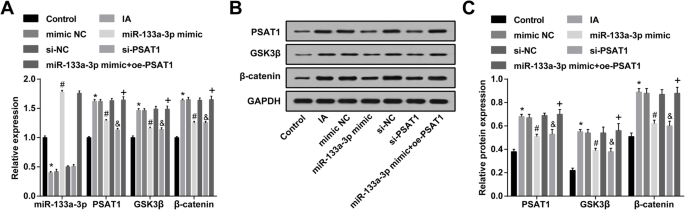
Повышающая регуляция miR-133a-3p и подавление PSAT1 снижает экспрессию PSAT1, GSK3β и β-катенина в ЭК IA. а Экспрессия miR-133a-3p, PSAT1, GSK3β и β-катенина в ЭК, обнаруженная с помощью RT-qPCR. б Белковые полосы PSAT1, GSK3β и β-катенина. c Экспрессия белков PSAT1, GSK3β и β-катенина в ЭК в каждой группе определяется с помощью вестерн-блоттинга. N =3, * P <0,05 по сравнению с контрольной группой. # P <0,05 по сравнению с группой мимических NC. & P <0,05 по сравнению с группой si-NC. + P <0,05 по сравнению с группой миметиков miR-133a-3p. Данные измерений были представлены как среднее значение ± стандартное отклонение, и данные были оценены с помощью одностороннего дисперсионного анализа с последующим апостериорным тестом Тьюки
Экспрессию PSAT1, GSK3β и β-катенина в ЭК IA проверяли с помощью вестерн-блоттинга и RT-qPCR. Было указано, что по сравнению с контрольной группой экспрессия PSAT1, GSK3β и β-катенина в группе IA была повышена (все P <0,05). Экспрессия PSAT1, GSK3β и β-катенина в группе IA, группе имитации NC и группе si-NC существенно не изменилась (все P > 0,05). Экспрессия PSAT1, GSK3β и β-катенина в группе миметика miR-133a-3p и группе si-PSAT1 была снижена по сравнению с таковой в группе миметика NC и группе si-NC (все P <0,05). По отношению к группе миметиков miR-133a-3p, экспрессия PSAT1, GSK3β и β-катенина была повышена в группе миметиков miR-133a-3p + oe-PSAT1 (все P <0,05) (рис. 5a – c).
Повышающая регуляция miR-133a-3p и понижающая регуляция PSAT1 уменьшают патологические изменения тканей IA
Проверяя гемодинамические изменения крыс после моделирования, мы отслеживали скорость кровотока крыс в каждой группе за 3 дня до операции и через 12 недель после интервенционного лечения. Было установлено, что не было очевидной разницы в скорости кровотока в каждой группе за 3 дня до операции ( P > 0,05). После 12 недель вмешательства скорость кровотока у крыс в группе IA снизилась по сравнению с таковой в нормальной группе ( P <0,05). Не было явной разницы в степени снижения скорости кровотока в группе IA, группе миметиков NC, группе si-NC и группе миметиков miR-133a-3p + oe-PSAT1 (все P > 0,05). По сравнению с группой si-NC и группой миметиков NC скорость кровотока была увеличена в группе миметика miR-133a-3p и группе si-PSAT1 (обе P <0,05). В отличие от группы миметиков miR-133a-3p, скорость кровотока была снижена в группе миметиков miR-133a-3p + oe-PSAT1 ( P <0,05) (рис. 6а).
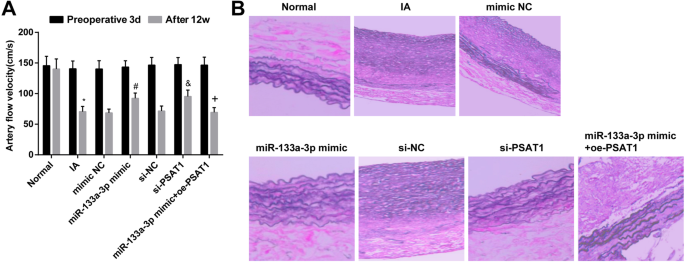
Повышенная регуляция miR-133a-3p и пониженная регуляция PSAT1 облегчают патологические изменения тканей IA. а Гемодинамические изменения в каждый момент времени после успешного моделирования на крысах. б Изменения ткани IA после трансфекции. нет =12, * P <0,05 по сравнению с нормальной группой. # P <0,05 по сравнению с группой мимических NC. & P <0,05 по сравнению с группой si-NC. + P <0,05 по сравнению с группой миметиков miR-133a-3p. Данные измерений были представлены как среднее значение ± стандартное отклонение, и данные были оценены с помощью одностороннего дисперсионного анализа с последующим апостериорным тестом Тьюки.
Изменения тканей IA подтверждены окрашиванием HE. Результаты показали, что в нормальной группе эластические волокна в среднем слое внутричерепной сосудистой ткани были аккуратными, появилась нормальная волнообразная структура эластичного белка, не было разрывов и деградации. По сравнению с нормальной группой просвет внутричерепной сосудистой ткани был увеличен, нормальная эластическая белковая волнообразная структура исчезла, эластичное волокно в среднем слое местного эластического белкового сосуда было разорвано, а некоторые из эластических волокон полностью разрушились. деградировал в группе ИА. Не было явных изменений в морфологии тканей IA в группе si-NC, группе миметиков NC, группе IA и группе miR-133a-3p mimic + oe-PSAT1. В отличие от группы si-NC и группы мимических NC, волновая структура эластичного белка во внутричерепной сосудистой ткани крыс в группе миметиков miR-133a-3p и группе si-PSAT1 существовала, а локальный эластический белок в сосудистых тканях существовал. структура была немного неупорядоченной, но не было разрушения и растворения (рис. 6б).
Высокоэкспрессируемый miR-133a-3p и низкоэкспрессируемый PSAT1 снижают экспрессию PSAT1, GSK3β, β-катенина, VEGF и MMP-9 в тканях IA in Vivo
Экспрессия miR-133a-3p в тканях IA in vivo была протестирована с помощью RT-qPCR, было высказано предположение, что по сравнению с нормальной группой экспрессия miR-133a-3p была снижена в группе IA ( P <0,05). Экспрессия miR-133a-3p в группе IA, группе имитации NC и группе si-NC явно не изменилась (все P > 0,05). Экспрессия MiR-133a-3p в мимической группе miR-133a-3p была повышена по сравнению с таковой в мимической группе NC ( P <0,05). По сравнению с группой si-NC, не было заметных изменений в экспрессии miR-133a-3p в группе si-PSAT1 ( P > 0,05). По сравнению с группой миметиков miR-133a-3p, экспрессия miR-133a-3p не показала значительных различий в группе миметиков miR-133a-3p + oe-PSAT1 ( P > 0,05) (рис. 7а).
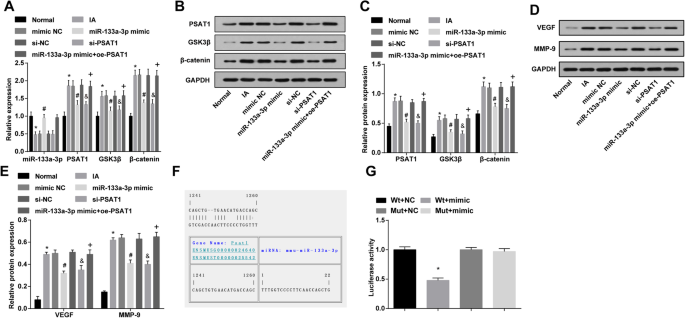
Сверхэкспрессия miR-133a-3p и низкая экспрессия PSAT1 снижают экспрессию PSAT1, GSK3β и β-катенина, VEGF и MMP-9 в тканях IA in vivo, а PSAT1 является геном-мишенью miR-133a-3p. а Обнаружение экспрессии miR-133a-3p, PSAT1, GSK3β и β-катенина в тканях IA крыс в каждой группе с помощью RT-qPCR. б Белковые полосы PSAT1, GSK3β и β-катенина. c Обнаружение экспрессии белков PSAT1, GSK3β и β-катенина в тканях IA крыс в каждой группе методом вестерн-блоттинга. г Полосы белков VEGF и MMP-9. е Обнаружение экспрессии белков VEGF и MMP-9 в тканях IA крыс в каждой группе методом вестерн-блоттинга. е Прогнозирование целевого сайта связывания PSAT1 с соответствующей miR-133a-3p с помощью Target Scan. г Результат двойного анализа репортерного гена люциферазы. а - е , n =12; е - г , N =3, * P <0,05 по сравнению с нормальной группой / группой Wt + NC. # P <0,05 по сравнению с группой мимических NC. & P <0,05 по сравнению с группой si-NC. + P <0,05 по сравнению с группой миметиков miR-133a-3p. Данные измерений были представлены как среднее значение ± стандартное отклонение, и данные были оценены с помощью одностороннего дисперсионного анализа с последующим апостериорным тестом Тьюки
Экспрессию PSAT1, GSK3β и β-катенина в тканях IA in vivo тестировали с помощью вестерн-блоттинга и RT-qPCR. Оказалось, что в отличие от нормальной группы, экспрессия PSAT1, GSK3β и β-катенина в группе IA была увеличена (все P <0,05). Экспрессия PSAT1, GSK3β и β-катенина в группе IA, группе имитации NC и группе si-NC существенно не изменилась (все P > 0,05). Экспрессия PSAT1, GSK3β и β-катенина в группе миметика miR-133a-3p и группе si-PSAT1 была снижена по сравнению с таковой в группе миметика NC и группе si-NC (все P <0,05). По сравнению с группой миметика miR-133a-3p, экспрессия PSAT1, GSK3β и β-катенина была усилена в группе миметика miR-133a-3p + oe-PSAT1 (все P <0,05) (рис. 7a – c).
Вестерн-блоттинг использовали для проверки экспрессии VEGF и MMP-9 в тканях IA in vivo; результаты показали, что по сравнению с нормальной группой, экспрессия VEGF и MMP-9 в группе IA была увеличена (оба P <0,05). Экспрессия VEGF и MMP-9 в группе IA, группе имитации NC и группе si-NC явно не изменилась (все P > 0,05). По отношению к группе миметиков NC и группе si-NC, экспрессия VEGF и MMP-9 в группе миметиков miR-133a-3p и группе si-PSAT1 снижалась (все P <0,05). По сравнению с группой миметиков miR-133a-3p, экспрессия VEGF и MMP-9 была повышена в группе миметиков miR-133a-3p + oe-PSAT1 (все P <0,05) (рис. 7г, д).
PSAT1 является целевым геном miR-133a-3p
Программное обеспечение для онлайн-прогнозирования (https://cm.jefferson.edu/rna22/Precomputed/) использовалось для прогнозирования и анализа целевого сайта связывания PSAT1 с miR-133a-3p и последовательности области 3'UTR, объединенной PSAT1 и miR-133a-3p. Чтобы доказать, что предсказанный сайт связывания miR-133a-3p приводил к изменению активности люциферазы, были разработаны последовательность мутации и дикая последовательность PSAT1, удаляющая 3'UTR сайт связывания miR-133a-3p. Активность люциферазы подтверждали котрансфекцией миметика miR-133a-3p и рекомбинантных плазмид WT (Wt-miR-133a-3p / PSAT1) или MUT (Mut-miR-133a-3p / PSAT1) в сосудистых ЭК. Результаты показали, что миметик miR-133a-3p не оказывает заметного влияния на активность люциферазы в группе Mut-miR-133a-3p / PSAT1 ( P > 0,05), тогда как активность люциферазы в группе Wt-miR-133a-3p / PSAT1 была заметно снижена ( P <0,05) (рис. 7е, ж).
Обсуждение
IA - это аномальное расширение внутричерепной артерии, которое ослабляет артериальную стенку за счет непрерывного выталкивания сосудистой стенки наружу, что приводит к более высокому риску разрыва аневризмы [17]. В исследовании, проведенном Liu et al., Было показано, что некоторые miRNAs участвуют в модуляции клеточной пролиферации гладкомышечных клеток сосудов, которая тесно связана с IA [18]. Кроме того, недавнее исследование предоставило доказательство того, что циркулирующие miRNAs могут использоваться в качестве нового биомаркера для оценки возможности возникновения IA у лиц с высоким риском [19]. Принято считать, что PSAT1 может участвовать в состояниях шизофренического спектра и изменять метаболизм серина [20]. Настоящее исследование было разработано для изучения регуляторной роли модулированного miR-133a-3p повреждения эндотелия сосудов и запуска IA посредством модуляции пути передачи сигналов PSAT1 / GSK3β / β-катенина.
В данном исследовании была проанализирована взаимосвязь между экспрессией miR-133a-3p и клинико-патологическими особенностями IA, и результаты показали, что экспрессия miR-133a-3p не связана с возрастом, полом, формой и положением аневризмы, а связано с количеством и размером аневризмы. Некоторые ученые считали, что напряжение сдвига регионального кровотока в стенке артерии индуцировало индукцию экспрессии хемоаттрактантного белка-1 моноцитов (MCP-1) и макрофагального воспалительного белка 1α (MIP-1α) фибробластами и ЭК сосудов внутри сосудистой стенки. Высокореактивные хемотаксические факторы MCP-1 и MIP-1α вызывали агрегацию макрофагоцитов в сосудистой стенке и опосредовали воспалительную реакцию, затем вызывали возбуждение ядерного фактора транскрипции c-Jun и затем регулировали активацию активированного белка 1 (AP- 1), затем активировал промотор MMP-9 в его структурном домене, чтобы повысить экспрессию мРНК MMP-9, и, наконец, вызвал растворение внеклеточного матрикса сосудистой стенки, вызывая образование внутричерепной аневризмы [21,22,23]. Сайто и др. [24] обнаружили, что MMP-9-положительные клетки происходили в основном из средней и внешней мембран артериальных макрофагов, что свидетельствовало о том, что MMP-9, экспрессируемый макрофагами, опосредует дегенерацию артериальной стенки, приводящую к образованию артериальной аневризмы. Вышеупомянутые исследования показали, что MMP-9 был связан с образованием IA. Результаты нашего исследования показали, что MMP-9 активируется в IA; таким образом, мы предположили, что mR-133a-3p может участвовать в возникновении и развитии IA путем регуляции пути PSAT1 / GSK3β / β-catenin и дальнейшей регуляции MMP-9. В нашем исследовании мы обнаружили, что восстановление miR-133a-3p снижает экспрессию PSAT1, GSK3β, β-катенина и MMP-9 в тканях внутричерепной аневризмы. Мы проведем соответствующее исследование в будущем исследовании, чтобы проверить наши выводы.
Наше исследование предоставило существенные доказательства в отношении того, что экспрессия miR-133a-3p была снижена, а PSAT1, GSK3β и β-катенин были повышены в IA. Новые данные свидетельствуют о том, что miR-133a-3p играет подавляющую роль в различных типах опухолей. Недавнее исследование показало, что экспрессия miR-133a-3p резко деградировала в тканях рака груди по сравнению с таковой в нераковых тканях [25]. Другое исследование показало, что экспрессия miR-133a-3p снижена в тканях распространенного рака простаты (РПЖ) по сравнению с таковой в прилегающих нормальных тканях или тканях доброкачественных поражений простаты, особенно в тканях с метастатическим РПЖ в кости [26]. Стимулирующий эффект PSAT1 при других типах заболеваний описан в некоторых литературных источниках. Сообщается, что экспрессия PSAT1 была заметно повышена при немелкоклеточном раке легкого (НМРЛ) и предсказывала плохой клинический исход у пациентов с НМРЛ [27]. Кроме того, PSAT1 считается самым активным геном в опухолях CRC, а также высоко экспрессируется у пациентов с химиорезистентными заболеваниями [28]. Было показано, что активность GSK3β повышена в раковых тканях [29]. Более того, уровень фосфорилирования GSK3β, а также экспрессия ядерного β-катенина также увеличиваются, что позволяет предположить, что путь GSK3β / β-катенина может участвовать в регуляции остеопонтина [30].
Другие результаты, полученные из наших данных, свидетельствуют о том, что повышающая регуляция miR-133a-3p и понижающая регуляция PSAT1 подавляли апоптоз и повышали пролиферацию и миграцию IA ECs, снижали экспрессию VEGF и MMP-9 в тканях IA. Ранее предполагалось, что сверхэкспрессия miR-133a-3p повторно тренирует инвазию, рост и митоз клеток плоскоклеточной карциномы полости рта, воздействуя на коллаген типа I alpha 1 (COL1A1) [31]. Сообщается, что высокоэкспрессируемая miR-133a-3p может подавлять размножение клеток ESCC, ускорять апоптоз клеток и снижать миграцию и инвазию клеток ESCC путем нацеливания на COL1A1 [32]. Другое исследование подтвердило, что временная повышающая регуляция miR-133a-3p подавляет способность к миграции, инвазии и росту клеток карциномы желчного пузыря посредством прямого воздействия на белок Jκ, связывающий сигнал рекомбинации [33]. Сходным образом это исследование предполагает, что miR-133a-3p проявляет свою роль в IA посредством нацеливания на PSAT1. Показано, что сверхэкспрессия PSAT1 усиливает рост клеток ESCC и инвазию матригеля in vitro, а инъекция мышам клеток ESCC с высокой экспрессией PSAT1 индуцирует образование опухоли in vivo [14]. Другое исследование также показало, что PSAT1 высоко экспрессируется и прогнозирует плохой клинический исход для пациентов, а также усиливает опухолегенез и пролиферацию клеток in vivo и in vitro [13]. Предыдущие исследования в целом подтверждают, что PSAT1 способствует прогрессированию клеточного цикла, пролиферации и онкогенезу посредством экспериментов с потерей и увеличением функции [27]. Было указано, что ММП состоят из ряда ферментов, которые расщепляют белковые субстраты на основе консервативного механизма, относящегося к активации связанной с активным сайтом молекулы воды через Zn 2+ ion [34]. MMP-9 - это отдельная протеаза, которая оказывает огромное влияние на многие биологические процессы [35]. В исследовании утверждается, что MMP-9 повышен в группах с аневризмой по сравнению с контрольной группой [36]. Фактор роста эндотелия сосудов-A (VEGF-A) признан ключевым модулятором эндоваскулярной дифференцировки трофобласта [37]. Исследование показало, что подавленная экспрессия PSAT1 подавляет экспрессию фосфорилирования VEGF, β-катенина и GSK3β [10].
Заключение
Вкратце, наше исследование подтверждает нашу гипотезу о том, что избыточная экспрессия miR-133a-3p или подавление PSAT1 сдерживают повреждение эндотелиальных клеток и ускоряют пролиферацию эндотелиальных клеток посредством ингибирования пути GSK3β / β-catenin в IA. Эти результаты позволяют по-новому взглянуть на новую таргетную терапию ИА. Эти находки подчеркивают роль miR-133a-3p в IA в отношении пути PSAT1 / GSK3β / β-catenin. Однако нельзя сделать однозначный вывод об эффектах miR-133a-3p и PSAT1 из-за ограниченного количества известных исследований по этому вопросу. Его необходимо тщательно контролировать и соответствующим образом сообщать в будущих клинических испытаниях.
Доступность данных и материалов
Не применимо
Сокращения
- miR-133a-3p:
-
МикроРНК-133a-3p
- IA:
-
Внутричерепная аневризма
- PSAT1:
-
Фосфосерин аминотрансфераза 1
- GSK3:
-
киназа β-гликогенсинтазы 3β
- SAH:
-
Субарахноидальное кровоизлияние
- MiRNA:
-
МикроРНК
- CRC:
-
Колоректальный рак
- ESCC:
-
Плоскоклеточный рак пищевода
- HE:
-
Гематоксилин-эозин
- PBS:
-
Фосфатный буферный раствор
- MMP:
-
Матрикс-металлопротеиназа
- VEGF:
-
Фактор роста эндотелия сосудов
- NC:
-
Отрицательный контроль
- DAB:
-
Диаминобензидин
- FBS:
-
Фетальная бычья сыворотка
- UEAI:
-
Агглютинины Ulex europaeus I
- PI:
-
Иодид пропидия
- FITC:
-
V-флуоресцеина изотиоцианат
- DMSO:
-
Диметилсульфоксид
- OD:
-
Оптическая плотность
- SD:
-
Спраг-Доули
- SPF:
-
Отсутствие специфических патогенов
- GAPDH:
-
Глицеральдегидфосфатдегидрогеназа
- BCA:
-
Бицинхониновая кислота
- 3′UTR:
-
3 ′ Непереведенный регион
- WT:
-
Дикий тип
- MUT:
-
Мутант
- ANOVA:
-
Дисперсионный анализ
- NSCLC:
-
Немелкоклеточный рак легкого
- VEGF-A:
-
Фактор роста эндотелия сосудов-А
Наноматериалы
- Усилитель с общим эмиттером
- Получение наночастиц mPEG-ICA, нагруженных ICA, и их применение в лечении LPS-индуцированного повреждения клеток H9c2…
- Доставка наночастиц артесуната увеличивает противоопухолевую эффективность за счет активации митохондриал…
- Контроль роста высокоднородных нанопроволок селенида индия (In2Se3) с помощью процесса быстрого термического о…
- Легкий синтез червоточиноподобного мезопористого оксида олова за счет самосборки, вызванной испарением, и у…
- Разработка и регулировка работы выхода графена с помощью размера, модификации, дефектов и легирования:исслед…
- Исследование солнечного элемента из кристаллического кремния с черным слоем кремния на задней панели
- Настройка режима скольжения-качения углеродных нанотрубок с помощью гидроксильных групп
- Эффективное поглощение света с использованием решеток двусторонней пирамиды для тонкопленочных кремниевых…
- Эффект апоптоза наночастиц золота, модифицированных литохолевой кислотой, на раковые клетки печени