


Доставка наночастиц артесуната увеличивает противоопухолевую эффективность за счет активации митохондриально-опосредованного апоптоза клеток
Аннотация
Считалось, что артемизинин и его производные обладают широким спектром противораковых действий и вызывают значительные противораковые эффекты в опухолевых клетках. Артемизинин и его производные могли быстро всасываться, и они были широко распространены, избирательно убивая опухолевые клетки. Поскольку низкие концентрации артесуната в первую очередь зависели от онкоза для индукции гибели опухолевых клеток, его противоопухолевые эффекты были нежелательными и ограниченными. Чтобы получить лучший противоопухолевый эффект, в этом исследовании мы использовали преимущества новой нанотехнологии для разработки новых наночастиц бычьего сывороточного альбумина, нагруженных артесунатом, для достижения митохондриального накопления артесуната и индукции митохондриально-опосредованного апоптоза. Результаты показали, что по сравнению с зависимостью свободного артесуната от онкотической смерти, наночастицы бычьего сывороточного альбумина, нагруженные артесунатом, показали более высокую цитотоксичность, а их значительные апоптотические эффекты были вызваны распределением артесуната в митохондриях. Это открытие указывает на то, что наночастицы бычьего сывороточного альбумина, нагруженные артесунатом, нарушают целостность митохондрий и активируют митохондриально-опосредованный апоптоз клеток, регулируя белки, связанные с апоптозом, и способствуя быстрому высвобождению цитохрома С.
Фон
Артемизинин и его производные широко используются для лечения малярии из-за их высокой противомалярийной активности и низкой токсичности. Исследователи также обнаружили, что артемизинин и его производные демонстрируют значительную противоопухолевую активность благодаря небольшим токсическим побочным эффектам и большей переносимости пациентами [1]. Сообщалось, что артесунат (Ats) определенно ингибировал рост опухолевых клеток и, кроме того, индуцировал значительные противораковые эффекты в опухолевых клетках [2,3,4]. Некоторые эксперименты показали, что Ats вызывают разную степень апоптоза и онкоза в опухолевых клетках через 48 часов, и что степень апоптоза и онкоза зависит от дозы Ats. В низких концентрациях Ats не вызывал явного апоптоза в опухолевых клетках, а гибель клеток, вызванная Ats, сопровождалась смертью, подобной онкозу [5,6,7,8]. Для получения большего противоопухолевого эффекта применялась более высокая доза Ats, но это дополнительно подтвердило его серьезную токсичность и подавление функции костного мозга. Следовательно, необходимо найти эффективное лечение, позволяющее снизить эффективную дозировку Атс, чтобы повысить его противоопухолевую эффективность [9,10,11]. Было обнаружено, что митохондрии играют важную роль в регуляции апоптотических и онкотических эффектов Ats. Митохондрии также участвуют в регуляции процесса трансдукции широкого спектра апоптотических сигналов [12,13,14,15,16,17]. Когда митохондрии подвергались атаке лекарствами, их проницаемость увеличивалась, а мембранный потенциал снижался, что приводило к набуханию эндометрия митохондриальной мембраны и быстрому высвобождению цитохрома С из митохондрий в цитоплазму [18,19,20]. Кроме того, были активированы некоторые белки семейства каспаз, и была вызвана каскадная реакция апоптоза клеток.
Чтобы усилить противоопухолевые эффекты Ats, было предпринято множество новых попыток увеличить распределение лекарства в опухолевых клетках или улучшить адресную доставку лекарств в органеллы клеток, чтобы вызвать гибель клеток [21,22,23]. Наночастицы (НЧ) как ключевой инструмент целенаправленного лечения рака широко изучались, и они показали многообещающий потенциал. Поскольку НЧ имели меньший размер частиц и большую площадь поверхности, они могли проникать в кровоток через капилляры, проходить через щель эндотелиальных клеток и мигрировать к месту опухоли, таким образом достигая целевого распределения лекарственного средства и увеличивая биодоступность лекарственного средства. . Более того, НЧ могут контролировать высвобождение лекарства через разложение биоматериала по длинной и плавной схеме, в конечном итоге продлевая период полужизни, улучшая эффективную концентрацию в крови и уменьшая частоту дозирования. Прежде всего, НЧ, нагруженные лекарством, могут быть доставлены в определенные места внутри клеток, повышая эффективность лечения [24,25,26].
Чтобы усилить противоопухолевые эффекты Ats в низких концентрациях, мы попытались создать новые НЧ бычьего сывороточного альбумина (BSA), нагруженные Ats. Из-за низкого pH в опухолевых клетках, накопления большого количества протонов водорода, присутствующих на внешней митохондриальной мембране или в межмембранном пространстве, напротив, межмитохондриальная мембрана богата отрицательным зарядом из-за своего химического состава и матрикса митохондрий. секреция, которая создает электроположительный внешний и отрицательный внутренний трансмембранный потенциал, что может способствовать доставке BSA. Затем массовое накопление Ats в митохондриях может эффективно запускать митохондриально-опосредованный апоптоз. Результаты показали, что по сравнению с типичной онкотической смертью, вызванной свободными Ats, Ats специфически переносился в митохондрии при посредничестве НЧ BSA и способствовал опосредованной митохондриями активации белков каспаз, связанных с апоптозом. Это вызвало значительный апоптоз клеток, тем самым подчеркивая более высокую цитотоксичность.
Методы
Материалы
BSA был приобретен у Sigma-Aldrich Co. (Сент-Луис, Миссури, США), а Ats был приобретен у Guilin Pharmaceutical Corporation (Гуйлинь, Китайская Народная Республика). Клетки SMMC-7721 и клетки Plc были приобретены в Институте биохимии и клеточной биологии Китайской академии наук (Шанхай, Китайская Народная Республика). Все остальные закупленные химические вещества были аналитической чистоты; они были получены от различных поставщиков.
Подготовка и характеристика загруженных Ats NP BSA
Согласно ранее опубликованной литературе [27], НЧ БСА, нагруженные Ats, получали методом десольватации. Вкратце, НЧ БСА, нагруженные Ats, получали путем быстрого добавления 1,0 мл безводного спирта, содержащего определенное количество Ats, в 0,5 мл раствора BSA при 37 ° C до опалесценции. После удаления этанола с помощью роторного испарителя НЧ БСА, нагруженные Ats, дополнительно осаждали из среды, а затем добавляли 8% глутаральдегид в воде (0,5 мкл / мг БСА), чтобы вызвать сшивание частиц при перемешивании суспензии в течение периода времени. от 24 ч. Наконец, НЧ собирали и трижды промывали деионизированной водой для дальнейшего анализа их физических характеристик, включая их гидродинамический диаметр, индекс полидисперсности (PDI), дзета-потенциал и морфологию, используя Brookhaven Zetasizer (Brookhaven Instruments Corporation, Holtsville, NY, USA). и просвечивающий электронный микроскоп (JEM-1200EX; JEOL, Токио, Япония). Определение эффективности инкапсуляции Ат в НЧ БСА оценивалось с использованием ранее описанного метода [27].
Анализ МТТ
Два вида линий опухолевых клеток, клетки SMMC-7721 и клетки Plc, отдельно инкубировали с 20% фетальной бычьей сывороткой (FBS). Плотность роста клеток доводили до 1 × 10 6 . клеток / мл по количеству клеток, а затем клеточные суспензии разбавляли до 1 × 10 5 клеток / мл. Разбавленные суспензии дополнительно добавляли отдельно в 96-луночный планшет (100 мкл на лунку, примерно 1 × 10 4 клеток / лунку) для непрерывной инкубации в течение 24 ч при 37 ° C в условиях 5% CO 2 и 95% O 2 . Среду заменяли бессывороточной средой в присутствии либо свободных Ats, либо загруженных Ats НЧ БСА с различными концентрациями Ats, и затем инкубировали в течение 24 часов. Всего 50 мкл 3- (4,5-диметилтиазол-2-ил) -2,5-дифенилтетразолийбромида (МТТ) (5 мг / мл) добавляли в каждую лунку и инкубировали в течение 4 ч для прекращения культивирования. Когда тетразолиевый краситель МТТ был восстановлен до нерастворимого формазана, 96-луночные планшеты центрифугировали при 1000 об / мин в течение 5 минут, и супернатант сливали из каждой лунки с последующим добавлением 150 мкл диметилсульфоксида (ДМСО), что полностью растворил кристаллы. Оптическую плотность раствора измеряли с помощью считывающего устройства для микропланшетов (Syneray-2; BioTek Instruments, Inc., Winooski, VT, USA) при 490 нм.
Внутриклеточное распределение группы NP BSA в клетках
Клетки SMMC-7721 и клетки Plc в логарифмической фазе отбирали и обрабатывали расщеплением трипсином; концентрация клеток была доведена до 1 × 10 6 клеток / мл. Затем культивированные клетки добавляли в 6-луночный планшет для культивирования клеток для прилипания, и культуральную среду удаляли с последующим добавлением меченых родамином B НЧ BSA. Ядро окрашивали Hoechst (синий) в течение 15 мин при 37 ° C, а митохондрии окрашивали Mitotracker Green FM. Расположение НЧ БСА в клетках отслеживали с помощью конфокальной лазерной сканирующей микроскопии (FluoView FV10i; Olympus Corporation, Токио, Япония).
Возможное изменение митохондриальной мембраны
JC-1 можно использовать для определения изменений потенциала митохондриальной мембраны. Когда потенциал митохондриальной мембраны был высоким, JC-1 мог свободно проходить через клеточную мембрану и формировать агрегаты внутри митохондрий, проявляя красную флуоресценцию (длина волны возбуждения 525 нм; длина волны излучения 590 нм); когда потенциал митохондриальной мембраны снижался, JC-1 переносился из митохондриального матрикса в цитоплазму клетки с образованием зеленого флуоресцентного мономера (длина волны возбуждения 490 нм; длина волны излучения 530 нм). Клетки SMMC-7721 и клетки Plc соответственно высевали в конфокальные чашки для достижения плотности 1 × 10 6 клеток / мл при непрерывной инкубации в течение 12 часов. Затем культуральную среду отбрасывали и в чашку добавляли бессывороточную культуральную среду, содержащую дисперсию Ats или загруженных Ats НЧ BSA. Через 9 ч среду удаляли и клетки дважды промывали PBS с последующим добавлением 2 мл JC-1 в концентрации 2 мкмоль / л; Затем клетки инкубировали в течение 30 мин при 37 ° C в темноте. Конфокальный микроскоп с лазерным сканированием (FluoView FV10i; Olympus Corporation) использовался для наблюдения за изменениями изображения митохондриальной мембраны.
Измерение продукции ROS и окрашивание эндоплазматической сети
Клетки инкубировали с 20% FBS, и плотность роста клеток доводили до 1 × 10 6 . клеток / мл по количеству клеток; а затем клеточные суспензии разбавляли до 1 × 10 5 клеток / мл. Разбавленные суспензии дополнительно добавляли в 96-луночные планшеты (100 мкл на лунку, примерно 1 × 10 4 клеток / лунку) для непрерывной инкубации в течение 24 часов при 37 ° C и 5% CO 2 и 95% O 2 . Во-вторых, свободные Ats и загруженные Ats НЧ БСА инкубировали с клетками в течение 6, 12 и 24 часов с последующей непрерывной инкубацией с 10 мкМ 2,7-дихлорфлуоресцеина диацетата (DCFH-DA; Sigma-Aldrich Co.) для около 30 мин. Ледяной буфер PBS использовали для трехкратной промывки клеток для удаления неинтернализованных НЧ. Интенсивность внутриклеточной флуоресценции DCF, которая возбуждается при 485 нм и испускается при 530 нм, определялась с помощью считывающего устройства для микропланшетов (Synergy-2; BioTek Instruments) для исследования степени окислительного стресса. Испытуемые группы обрабатывали клетками SMMC-7721 и клетками Plc в течение 24 ч, и зонд ER-Tracker Blue-White DPX (Molecular Probes, Юджин, Орегон, США) добавляли в клетки для инкубации в течение 30 мин. После удаления загрузочного раствора и промывки клеток PBS изменение морфологии ER наблюдали с помощью конфокальной лазерной сканирующей микроскопии.
Оценка клеточного онкоза и апоптоза с помощью проточной цитометрии
Согласно протоколу нашего предыдущего исследования [28], анализ окрашивания аннексином V – флуоресцеинизотиоцианатом (FITC) / пропидиум иодидом (PI) был использован для оценки клеточного онкоза и апоптоза, вызванного свободными Ats и загруженными Ats НЧ BSA. Клетки лизировали типсином и высевали в шестилуночные планшеты в концентрации 1 × 10 6 . клеток / мл в течение 24 ч непрерывной инкубации. Затем культуральную среду удаляли и в лунки добавляли бессывороточную среду, содержащую свободные Ats и нагруженные Ats NP BSA. После обработки клетки собирали и суспендировали в буфере Николетти (Beijing 4A Biotech Co., Ltd., Пекин, Китайская Народная Республика), содержащем PI- и FITC-меченый аннексин V (AV-FITC). Морфологические изменения клеток наблюдали с помощью конфокальной лазерной сканирующей микроскопии. Для проверки скорости апоптоза и онкоза клеток, вызванных Ats-нагруженными НЧ, процентное содержание ранних апоптозных (Q4), онкотических (Q2), некротических (Q1) и живых клеток (Q3) было определено количественно с помощью проточной цитометрии.
Вестерн-блот-анализ белков, связанных с апоптозом, и цитохрома C в клетках
Для определения уровней относительных белков проводили вестерн-блоттинг, когда свободные Ats или загруженные Ats NP инкубировали с клетками SMMC-7721 в течение 24 часов. Клетки лизировали ледяным буфером для анализа радиоиммунопреципитации (RIPA), содержащим смесь ингибиторов протеаз и ингибиторов фосфатаз (Roche, Базель, Швейцария). Концентрации белка определяли с использованием модифицированного набора для анализа BSA (Thermo Fisher Scientific, Waltham, MA, USA) и нормализовали перед загрузкой с помощью электрофореза в 10% додецилсульфат натрия (SDS) в полиакриламидном геле (PAGE). Уровни целевых белков были сфотографированы и проанализированы с использованием системы анализа геля UVP (iBox Scientia 600; UVP, LLC., Upland, CA, USA).
Результаты
Характеристики NPs BSA, загруженных Ats, и исследование жизнеспособности сотовой связи
На рис. 1а, б видно, что НЧ БСА, нагруженные Ats, имели сферическую форму, и они были гомогенно диспергированы с более низким PDI на уровне 0,016. Средний размер частиц НЧ БСА, нагруженных Ats, составлял около 99,9 ± 2,3 нм, а дзета-потенциал был отрицательным и составлял около –25,6 ± 4,3 мВ. Профиль высвобождения показал плавное и продолжительное высвобождение, что показано на фиг. 1c. По сравнению с быстрым высвобождением свободных Ats в среде in vitro, Ats, захваченные в ядре НЧ БСА, медленно диффундировали из внутренней части НЧ в среду и демонстрировали плавный и устойчивый паттерн высвобождения из-за непрерывной деградации БСА. Более 85% свободных Ats было полностью высвобождено в течение первых 6 часов, в то время как общее накопленное количество лекарственного средства, высвобожденного из NP в среду в течение 48 часов, составило 78,9%. Это указывает на то, что НЧ могут контролировать высвобождение лекарства за счет разложения биоматериалов по продолжительной и плавной схеме, таким образом продлевая период полувыведения, улучшая эффективную концентрацию в крови и уменьшая частоту дозирования.
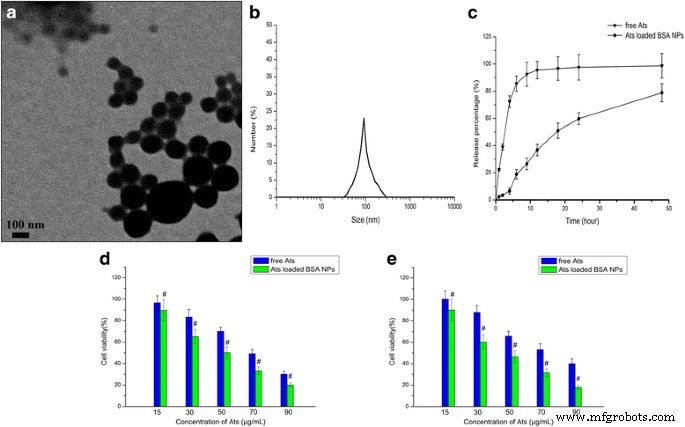
Характеристика Ats-нагруженных НЧ БСА. а ПЭМ-изображение НЧ БСА, нагруженных Ats. б Анализ динамического светорассеяния (DLS) полученных Ats-нагруженных НЧ BSA. c Профиль высвобождения Ats-нагруженных НЧ БСА in vitro в фосфатно-солевом буфере с pH 7,4 при 37 ° C в течение 48 часов. Жизнеспособность клеток SMMC-7721 ( d ) и ячейки Plc ( e ) после инкубации с различными количествами свободных Ats и НЧ BSA, нагруженных Ats, в течение 24 часов. Данные представлены как среднее значение ± стандартное отклонение ( n =3). # P <0,05 по сравнению с соответствующими бесплатными Ats
МТТ использовали для исследования ингибирующих эффектов свободных Ats и загруженных Ats НЧ БСА в клетках SMMC-7721 и клетках Plc в различные интервалы времени. Результаты (рис. 1d, e) показали, что цитотоксичность свободных Ats увеличивалась с увеличением концентрации лекарственного средства, а нагруженные Ats НЧ BSA демонстрировали постепенное усиление цитотоксичности. Это доказало, что Ats и нагруженные Ats NP BSA ингибируют рост опухолевых клеток и что коэффициент ингибирования зависит от дозы Ats. По сравнению со свободными Ats, нагруженные Ats НЧ БСА продемонстрировали более высокую цитотоксичность и более высокую чувствительность в обеих клетках, и они привели к большему ингибированию клеток. Как показано на фиг. 1d, e, обработка обеих клеток Ats-нагруженными НЧ BSA вызвала значительное снижение жизнеспособности клеток через 24 часа по сравнению с таковой для свободных Ats. Значения максимальной ингибирующей концентрации 50% (IC50) для клеток SMMC-7721 и клеток Plc, обработанных Ats-нагруженными НЧ БСА, составляли 50,1 и 44,9 мкг / мл через 24 ч соответственно, что сравнивается с полученными значениями 69,2 и 74,9. мкг / мл через 24 ч в клетках, обработанных свободными Ats. Это указывает на то, что когда Ats загружается в NP BSA, он может изменить свое внутриклеточное расположение, опосредованное NP, и в конечном итоге убивать больше клеток.
Поглощение НЧ БСА в клетках in vitro
Внутриклеточное распределение и расположение НЧ БСА в обоих типах опухолевых клеток наблюдали с помощью конфокальной лазерной сканирующей микроскопии, как показано на рис. 2. После совместного культивирования НЧ, меченных родамином В, с клетками в течение 3 ч, отчетливо наблюдалась красная флуоресценция. в цитоплазме; с течением времени большинство НЧ БСА интернализовалось внутри клетки и диффундировало в цитоплазму, проявляя усиленную зависящую от времени красную флуоресценцию. Также было замечено, что НЧ БСА, расположенные в цитоплазме, были совмещены с митохондриями, о чем свидетельствует появление желтой флуоресценции, что служило указанием на то, что собственная красная флуоресценция НЧ, меченных родамином B, и зеленая флуоресценция, испускаемая митохондриальный индикатор MitoTracker® green FM был объединен. Это доказало, что интернализованные НЧ БСА могут специфически накапливаться в митохондриях, что подчеркивает возможность доставки Ат в митохондрии при посредничестве НЧ БСА.
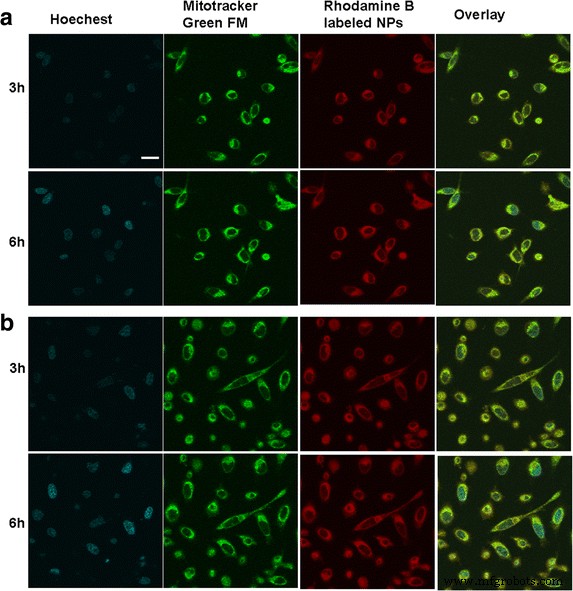
Клеточное распределение НЧ БСА in vitro после инкубации с различными опухолевыми клетками. Флуоресцентное изображение клеток SMMC-7721 ( a ) и ячейки Plc ( b ). Шкала масштаба , 100 мкм
Анализ митохондриального мембранного потенциала
Чтобы выяснить, влияют ли загруженные Ats НЧ БСА на функцию митохондрий после доставки Ats в митохондрии, были определены изменения потенциала митохондриальной мембраны. Фигура 3 продемонстрировала, что после окрашивания JC-1 большая часть митохондрий в опухолевых клетках, обработанных свободными Ats, проявляла сильную красную флуоресценцию и слабую интенсивность зеленой флуоресценции. Это предполагает, что большая часть JC-1 существует в агрегированном состоянии, что усиливает целостность митохондриальной мембраны и повышает потенциал. Напротив, когда JC-1 окрашивал нагруженные Ats клетки, обработанные НЧ BSA, митохондрии в обеих опухолевых клетках проявляли более сильную зеленую флуоресценцию, указывая на то, что митохондриальная мембрана была серьезно повреждена и ее потенциал был значительно снижен. В совокупности это доказало, что Ats был успешно доставлен в митохондрии при посредничестве НЧ БСА, что привело к деполяризации митохондриальной мембраны.
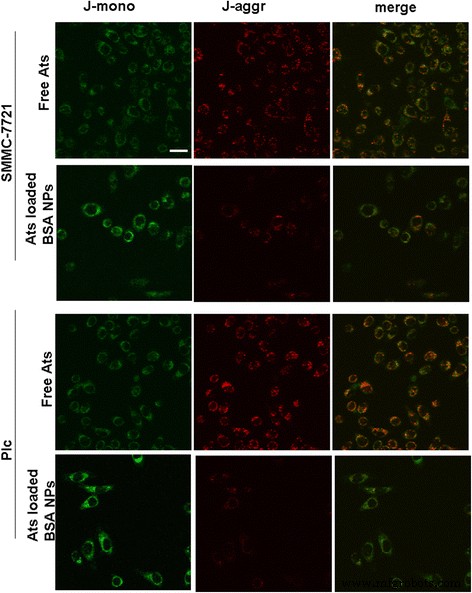
Изменение изображения митохондриального мембранного потенциала после инкубации свободных Ats и загруженных Ats НЧ BSA с клетками SMMC-7721 и клетками Plc. Шкала масштаба , 100 мкм
Измерение продукции ROS и окрашивание ER
Было широко подтверждено, что образование большого количества АФК может вызывать перекисное окисление фосфолипидов во внутренней митохондриальной мембране и что оно также может вызывать снижение потенциала митохондриальной мембраны, что приводит к быстрому высвобождению цитохрома С. Мы использовали DCFH- DA в качестве флуоресцентного зонда для обнаружения изменения ROS. DCFH-DA свободно проходил через клеточную мембрану в клетку и трансформировался в DCFH гидролизом эстеразы. Генерируемый DCFH не может проходить через клеточную мембрану и легко загружается в клетки. Внутриклеточные ROS окислили нефлуоресцентный DCFH до DCF с зеленым флуоресцентным цветом. Следовательно, обнаружение флуоресценции DCF может указывать на уровень внутриклеточных АФК.
Когда обе клетки обрабатывали свободными Ats и загруженными Ats НЧ BSA в течение определенного периода времени, количество внутриклеточных ROS также увеличивалось, показывая зависимость от времени. По сравнению со свободными Ats, генерация ROS в клетках SMMC-7721 и Plc, обработанных НЧ BSA, нагруженных Ats, была значительно увеличена. Фигуры 4a продемонстрировали, что уровни ROS в клетках SMMC-7721 и Plc, подвергнутых воздействию Ats-нагруженных НЧ BSA в течение 48 часов, были увеличены в 1,53 раза и 1,28 раза, соответственно, по сравнению с клетками SMMC-7721 и клетками Plc, обработанными. с бесплатными атс. Это подтвердило идею о том, что НЧ ускоряют продукцию внутриклеточных АФК. По сравнению с контрольной группой и свободными Ats, а также после обработки НЧ BSA, нагруженных Ats, интенсивность флуоресцентного окрашивания от ER-Tracker Blue-White DPX как ER-специфического красителя была значительно увеличена, что позволяет предположить, что стресс ER также был вызван в клетках, обработанных НЧ, нагруженных Ats, с соответствующим увеличением уровня АФК. Это открытие подчеркнуло, что Ats был специфически локализован в митохондриях, как опосредовано BSA NPs; это привело к значительному увеличению уровня бескислородных радикалов в клетках, что вызвало индукцию стресса ER и активацию митохондриального пути, чтобы вызвать каспазозависимый клеточный апоптоз.
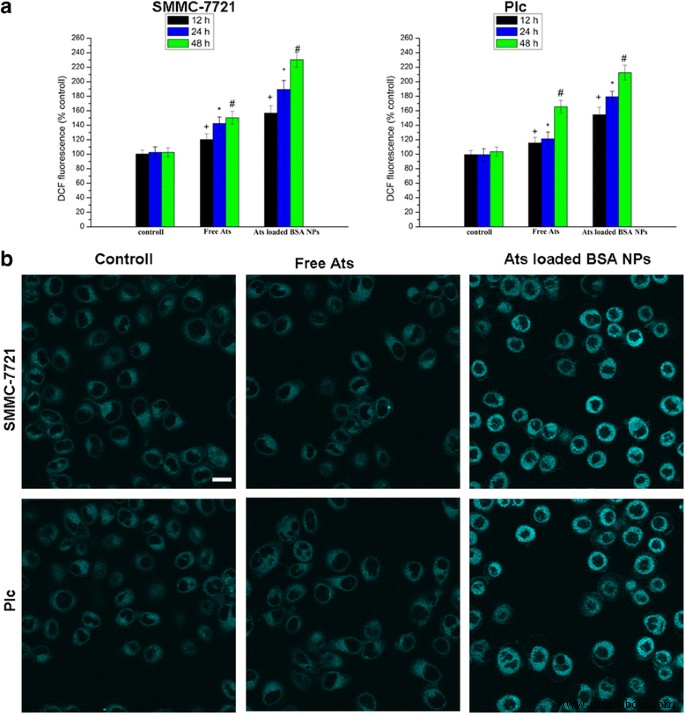
Количественная оценка генерации ROS в клетках, обработанных свободными Ats и загруженными Ats НЧ БСА в разное время ( a ). Окрашивание ER с помощью зонда ER-Tracker Blue-White DPX ( b ). Шкала масштаба , 100 мкм. Данные представлены как среднее значение ± стандартное отклонение ( n =3). + P <0,05 по сравнению с контрольной группой через 12 ч, * P <0,05 по сравнению с контрольной группой через 24 часа, # P <0,05 по сравнению с контрольной группой через 24 часа
Оценка апоптоза и некроза клеток
Клетки обрабатывали анализом окрашивания аннексином V-FITC / PI. Живые клетки не связывались с аннексином V-FITC / PI, поэтому флуоресценция не появлялась. Апоптотические клетки не связывались с PI, но их окрашивали аннексином V-FITC, что давало зеленую флуоресценцию. Напротив, для онкотических клеток их клеточные мембраны были в некоторой степени повреждены, а ядра клеток были расширены, чтобы распадаться на части, показывая зеленую и красную флуоресценцию. Как показано на фиг. 5a, по сравнению с контрольной группой, когда свободные Ats и загруженные Ats НЧ инкубировали с клетками в течение 24 ч, в клетках наблюдалась сильная зеленая и красная флуоресценция, что указывает на то, что свободные Ats и загруженные Ats НЧ БСА индуцированный онкоз опухолевых клеток и апоптоз. Особенно после обработки НЧ БСА, нагруженных Ats, интенсивности окрашивающей флуоресценции, полученные от аннексина V-FITC и PI, были значительно увеличены, что свидетельствует о значительном увеличении степени онкоза и апоптоза в клетках, обработанных НЧ, нагруженных Ats. / P> 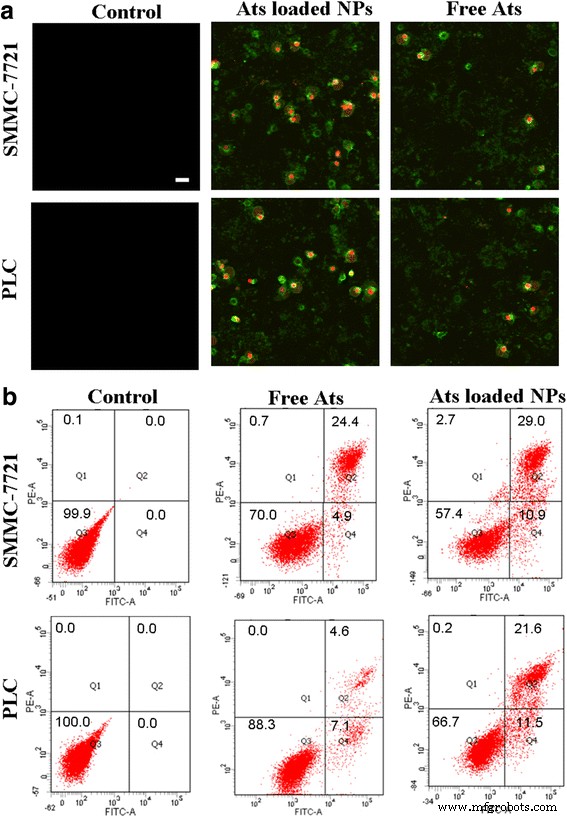
Морфология ультраструктурных изменений клеток, обработанных свободными Ats и загруженными Ats НЧ БСА с использованием окрашивания аннексином V-FITC / PI ( a ). Шкала масштаба , 100 мкм. Проточный цитометрический анализ клеточного апоптоза и онкоза через 24 часа инкубации со свободными Ats и загруженными Ats НЧ БСА, соответственно ( b )
Процент раннего апоптоза (Q4), онкотических (Q2), некротических (Q1) и живых клеток (Q3) показан на фиг. 5b. Это открытие продемонстрировало, что, когда клетки обрабатывали свободными Ats, онкотические уровни постепенно увеличивались до 24,4 и 4,6%, а скорость апоптоза оставалась на уровне 4,9 и 7,1% в клетках SMMC-7721 и Plc, соответственно, что позволяет предположить, что свободные Ats запускаются. возникновение онкоза и апоптоза, приводящих к гибели клеток. Напротив, НЧ БСА, нагруженные Ats, значительно улучшали скорость апоптоза и онкоза клеток. Отношения апоптоза были значительно увеличены до 10,9% в клетках SMMC-7721 и до 11,5% в клетках Plc. Онкотические отношения были увеличены до 29,0% в клетках SMMC-7721 и до 21,6% в клетках Plc. Это указывает на то, что митохондриальная доставка Ат при посредничестве НЧ БСА ускоряет гибель опухолевых клеток за счет усиления онкотических и апоптотических эффектов. НЧ БСА, нагруженные Ats, запускали процесс передачи апоптотического сигнала и способствовали митохондриально-опосредованной каскадной реакции клеточного апоптоза.
Вестерн-блоттинг
Чтобы изучить зависимость гибели клеток от апоптоза, индуцированного свободными Ats и нагруженными Ats NP, был проведен вестерн-блоттинг для обнаружения экспрессии белков апоптоза. Было обнаружено, что в обработанных Ats NPs клетках SMMC-7721 уровень внутриклеточной экспрессии белка Bax был значительно повышен (фиг. 6). Это открытие свидетельствует о том, что с помощью НЧ БСА Ats накапливается в митохондриях и вызывает митохондриальную дисфункцию. Цитоплазматический мономерный белок Bax переносился на внешнюю мембрану митохондрий и подвергался олигомеризации, образуя белковый канал на внешней мембране митохондрий, что в дальнейшем приводило к увеличению проницаемости мембраны. Уровень экспрессии цитохрома С в цитоплазме также был особенно и значительно повышен, и было обнаружено, что экспрессия каспазы-3 и каспазы-9 демонстрирует тенденцию к увеличению. Следовательно, благодаря более высокой проницаемости мембран митохондрий, цитохром С быстро высвобождается в цитоплазму, активируя белки, сигнализирующие о гибели клеток (каспазы), и способствуя каскадной реакции клеточного апоптоза. Напротив, свободные Ats не имели значительных различий в экспрессии белков, связанных с апоптозом, и цитохрома C, что позволяет предположить, что свободные Ats не запускали митохондриально-опосредованный апоптоз клеток и в первую очередь полагались на онкоз, чтобы привести к гибели клеток. НЧ БСА усиливали накопление препарата в митохондриях и активировали митохондриально-опосредованные апоптотические эффекты, что приводило к значительному апоптозу и увеличению экспрессии первичных белков, имеющих отношение к апоптозу, как показано в нашем Вестерн-блоттинговом анализе.
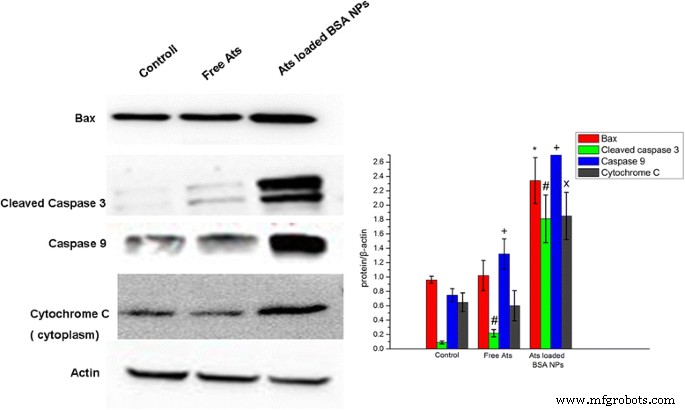
Вестерн-блоттинг-анализ уровней экспрессии расщепленной каспазы-3, каспазы-9, Вах и цитохрома С в клетках SMMC-7721. * P <0,05 по сравнению с экспрессией белка Bax в контрольной группе; # P <0,05 по сравнению с экспрессией расщепленной каспазы-3 в контрольной группе; + P <0,05 по сравнению с экспрессией белка каспазы-9 в контрольной группе; x P <0,05 по сравнению с экспрессией белка цитохрома С в контрольной группе. Данные были представлены как среднее значение ± стандартное отклонение ( n =3)
Обсуждение
Онкоз и апоптоз представляют собой два разных пути гибели клеток. Апоптоз - это активный процесс запрограммированной гибели клеток, происходящий в многоклеточных организмах. С другой стороны, онкоз описывает независимую от каспаз гибель клеток, которая характеризуется набуханием, повышенной проницаемостью и разрывом мембраны, что часто называют некрозом. Эта форма гибели клеток считается случайной и неконтролируемой. Основываясь на нашем исследовании, мы обнаружили, что Ats ингибируют рост опухолевых клеток и что коэффициент ингибирования зависит от дозы Ats. Атс в первую очередь зависел от степени онкоза и приводил к гибели клеток; он также активировал независимую от каспазы гибель клеток в форме онкоза. И наоборот, и отдельно от возникновения очевидной смерти, подобной онкозу, когда опухолевые клетки обрабатывали Ats-нагруженными НЧ БСА, нагруженные Ats НЧ БСА интернализовались в цитоплазму и быстро локализовались в митохондриях, высвобождая Ats, как опосредовано НП. Ats в митохондриях генерировали ROS и запускали стресс ER; он дополнительно активировал опосредованный митохондриями каспазозависимый клеточный апоптотический путь за счет снижения потенциала митохондриальной мембраны, высвобождения цитохрома С и стимулирования экспрессии белков Bax, расщепленной каспазы 3 и каспазы 9. Вместе взятые, загруженные Ats НЧ БСА увеличивали митохондриальная доставка Ats и увеличение степени онкоза и апоптоза, вызывая гибель клеток, тем самым увеличивая цитотоксичность препарата и вызывая значительную гибель клеток.
Выводы
Вкратце, мы пояснили, что свободный Ats в опухолевых клетках сильно зависит от степени онкоза, чтобы ингибировать пролиферацию опухолевых клеток в форме смерти, подобной онкозу; таким образом, цитотоксичность препарата была ограниченной и нежелательной. Напротив, нагруженные Ats НЧ БСА активировали митохондриальный апоптотический путь и одновременно запускали онкотические эффекты; вместе они усиливали синергетическую противоопухолевую эффективность Ats. The results of this study highlighted the significance of Ats-loaded BSA NPs in the enhancement of the cytotoxic and apoptotic effects of Ats, and they further signify the role of BSA NPs in diversifying the pathways of cell death induced by Ats. Compared with free Ats, Ats-loaded BSA NPs induced greater cytotoxicity and significant cell apoptosis effects in tumor cells.
Наноматериалы
- Усилитель с общим эмиттером
- Воздействие COVID-19 на производство
- Настройка токсичности активных форм кислорода для расширенной терапии опухолей
- Металлоорганические каркасы, реагирующие на окружающую среду, как система доставки лекарств для терапии опу…
- Влияние отношения Li / Nb на получение и фотокаталитические характеристики соединений Li-Nb-O
- Доставка ДНК-тетраэдра усиливает индуцированный доксорубицином апоптоз клеток рака толстой кишки HT-29
- Эффекты взаимодействия поверхностных плазмонных поляритонов и магнитных дипольных резонансов в метаматери…
- Исследование солнечного элемента из кристаллического кремния с черным слоем кремния на задней панели
- Оценка свойств доставки генов в антимикробные, апоптотические и раковые клетки для белковых наночастиц золо…
- Положительные эффекты автоматизации сельского хозяйства