


микроРНК-18a из макрофагов M2 ингибирует TGFBR3, способствуя прогрессированию карциномы носоглотки и росту опухоли через сигнальный путь TGF-β
Аннотация
Цели
Рак носоглотки (NPC) - это тип заболевания носоглотки с высокими метастазирующими и инвазионными свойствами. Доказано, что ассоциированные с опухолью альтернативно активированные (M2) макрофаги соединяются с NPC. Исходя из этого, данное исследование направлено на изучение механизма и участия микроРНК-18a (miR-18a) из макрофагов M2 в NPC.
Методы
Мононуклеарные клетки периферической крови были дифференцированы до макрофагов, и макрофаги были поляризованы до типа M2 с помощью интерлейкина-4. Клетки SUNE-1 и CNE2 трансфицировали восстановленным или истощенным miR-18a или рецептором трансформирующего фактора роста-бета III (TGFBR3), чтобы исследовать их роль в прогрессировании NPC с участием сигнального пути TGF-β. Затем клетки SUNE-1 и CNE2 культивировали совместно с макрофагами M2, обработанными восстановленными или истощенными miR-18a или TGFBR3, чтобы понять их совместную роль в NPC с участием сигнального пути TGF-β.
Результаты
MiR-18a был высоко экспрессирован, а TGFBR3 слабо экспрессирован в клетках NPC. Восстановление MiR-18a, нокдаун TGFBR3 или совместное культивирование с миметиками miR-18a или макрофагами M2, трансфицированными si-TGFBR3, способствовали развитию клеток SUNE-1, росту опухоли у мышей, снижению p-Smad1 / t-Smad1 и повышению p- Smad3 / t-Smad3. Подавление miR-18a, сверхэкспрессия TGFBR3 или совместное культивирование с ингибиторами miR-18a или макрофагами M2, трансфицированными OE-TGFBR3, подавляли прогрессирование клеток CNE2, рост опухоли у мышей, увеличивали p-Smad1 / t-Smad1 и снижали p-Smad3 / т-Smad3.
Заключение
Наше исследование выясняет, что miR-18a из макрофагов M2 приводит к ускоренному прогрессированию клеток NPC и росту опухоли у мышей nude посредством репрессии TGFBR3, наряду с инактивацией Smad1 и активацией Smad3.
Введение
Рак носоглотки (NPC) - злокачественная эпителиальная опухоль, склонная к локальной инфильтрации и ранним отдаленным метастазам [1]. Пациенты с NPC часто жалуются на паралич шестого нерва и синдром Хорнера [2]. В настоящее время применяемое лечение преимущественно состоит из лучевой терапии, а также комплексной лучевой терапии и химиотерапии [3]. К сожалению, лучевая терапия и химиотерапия неожиданно сопровождаются осложнениями, а приобретенная устойчивость к лучевой терапии затрудняет исходы NPC [3]. Учитывая, что задача выявления потенциальной таргетной терапии является приоритетной.
Доказано, что дисрегулируемые микроРНК (miRNA) участвуют в онкогенезе, метастазировании, инвазии и устойчивости к лучевой и химиотерапии NPC [1]. Как подсемейство miRNAs, miR-18a, как обнаружено, способствует прогрессированию NPC посредством супрессора морфогенеза в ингибировании гениталий 1 и активации пути mTOR [4]. Кроме того, подтверждается, что miR-18a активирует пролиферацию и метастазирование клеток NPC посредством регуляции DICER1 [5]. Кроме того, доказано, что прогрессия клеток NPC управляется miR-18a через нарушение биогенеза miRNA [6]. Более того, miR-18a, как было задокументировано, критически функционирует в метастазах NPC [7]. Альтернативно активированные (M2) макрофаги являются важными компонентами солидных и гематологических злокачественных новообразований и связаны с прогрессированием, метастазированием и резистентностью к терапии [8]. M2-поляризованные опухолевые макрофаги связаны с плохим прогнозом NPC [9]. Интересно зафиксировано различие макрофагов M2 в вирус-отрицательных по Эпштейну-Барр и положительных по вирусу Эпштейна-Барр NPC [10]. Существует исследование, в котором показано, что miR-18a ингибирует метастазирование клеток рака толстой кишки в печень, индуцируя макрофаги M1 [11]. Рецептор трансформирующего фактора роста-бета III (TGFBR3) представляет собой корецептор TGF-β, который обеспечивает лиганд рецептора TGF-β типа II для стимуляции передачи сигналов и баланса клеточной поверхности, а растворимый TGFBR3 является регулятором во время прогрессирования рака [12]. Сообщается, что низкоэкспрессируемый TGFBR3 индуцирует иммунотолерантное микроокружение опухоли [13]. Напротив, временная сверхэкспрессия TGFBR3 индуцирует апоптоз в клетках NPC человека [14]. Насколько нам известно, поляризация макрофагов M2, индуцированная miR-181a, способствует метастазированию опухолевых клеток, опосредованному макрофагами M2, посредством Kruppel-подобного фактора 6 и оси α, связывающей CCAAT / энхансер [15].
В совокупности, хотя многие исследования открыли независимую роль макрофагов miR-18a, TGFBR3 и M2 в NPC, комбинированные взаимодействия между этими тремя факторами все еще неуловимы. Учитывая, что это исследование запущено для расшифровки механизмов и участия этих факторов в NPC.
Материалы и методы
Этика
Эксперимент был одобрен этическим комитетом Третьей больницы Сянъя Центрального Южного университета и соответствовал стандартам медицинской этики. Это исследование было проведено с письменного согласия всех доноров. Эксперименты на животных проводились в соответствии с требованиями национальных правил обращения с лабораторными животными и их использования.
Сбор мононуклеарных клеток периферической крови
Моноциты были получены из периферической крови здоровых доноров адгезивным методом. Образцы периферической крови были получены от здоровых доноров отделения гематологии больницы Третьего Сянъя Центрального Южного университета. Макрофаги, происходящие из моноцитов, получали путем прикрепления к пластику мононуклеарных клеток периферической крови (PBMC), ранее выделенных центрифугированием в градиенте плотности (Ficoll-Paque, GE Healthcare) из препаратов лейкоцитарной пленки крови от здоровых доноров. Затем 2,0 × 10 6 PBMC культивировали в среде Игла, модифицированной Дульбекко (DMEM), с добавлением 10% сыворотки крови человека (Millipore, Bedford, MA, USA) и пенициллина / стрептомицина на 12-луночных планшетах (NalgeNunc, NY, США). Когда PBMC прикреплялись к стенке в течение 2–3 часов, супернатант и суспендированные PBMC удалялись повторно для получения прикрепленных моноцитов.
Поляризация макрофагов
Моноциты были индуцированы к дифференцировке в макрофаги под действием колониестимулирующего фактора макрофагов человека (M-CSF) и поляризованы в макрофаги M2 с помощью интерлейкина (IL) -4.
Индукция макрофагов
Моноциты культивировали в 20% -ной фетальной бычьей сыворотке (FBS) -DMEM и добавляли M-CSF (100 мкг, Peprotech, NJ, USA) до конечной концентрации 100 нг / мл. Среду обновляли наполовину каждые 3 дня, а затем добавляли 100 нг / мл M-CSF. После культивирования к 7–8-му дню часть клеток собирали, и поверхностные маркеры макрофагов CD68 [16], CD163 [17] и CD206 [18] тестировали иммунофлуоресцентным анализом [19] для идентификации макрофагов.
Поляризация макрофагов
Макрофаги были поляризованы до макрофагов M2 путем добавления 20 нг / мл IL-4 (Peprotech) в среду для дифференцировки в течение еще 24 часов. Часть образцов макрофагов M2 использовали для обнаружения проточной цитометрией. Образцы были разделены на 3 пробирки:пробирка 1 представляла собой ту же партию прикрепившихся макрофагов без стимуляции IL-4; пробирки 2 и 3 представляли собой прикрепленные макрофаги, стимулированные IL-4. При загрузке образцы разбавляли примерно до 10 000 клеток и добавляли неспецифический иммуноглобулин G для блокирования рецептора Fc. Затем в образцы добавляли неспецифическое изотипическое антитело, меченное PE антитело CD68 и меченное PE антитело CD163 (оба от Biolegend, Калифорния, США). Инкубировали в течение 30 минут и промывали 0,5% бычьим сывороточным альбумином в фосфатно-солевом буфере (PBS), образцы центрифугировали и превращали в суспензию клеток с 500 мкл PBS для обнаружения.
Количественная полимеразная цепная реакция с обратной транскрипцией
Количественная полимеразная цепная реакция с обратной транскрипцией (RT-qPCR) была использована для обнаружения miR-18a, CCL22, рецептора γ, активируемого пролифератором пероксисом (PPAR-γ), и экспрессии мРНК TGFBR3 в собранных клетках.
Тотальную РНК экстрагировали из клеток с помощью Trizol (Invitrogen, Carlsbad, CA, USA) и обратно транскрибировали в комплементарную РНК с помощью набора для синтеза первой цепи miRNA Mir-X (Clontech, Mountain View, CA, USA) для miR-18a и PrimeScriptTM RT. Master Mix Kit (Такара, Далянь, Китай) для CCL22, PPAR-γ и TGFBR3. U6 и глицеральдегид-3-фосфатдегидрогеназа (GAPDH) были контролями нагрузки для miR-18a, CCL22, PPAR-γ и TGFBR3. SYBR @ Премикс Ex Taq ™ II (Perfect Real Time) (Takara) в системе LightCycler 480 II (Roche Diagnostics, Индиана, США) использовали в ПЦР. Расчет данных оценивался 2 - △△ CT метод. Праймеры для ПЦР представлены в таблице 1.
Вестерн-блоттинг
Вестерн-блоттинг применялся для обнаружения TGFBR3, общего (t) -Smad1, фосфорилированного (p) -Smad1, t-Smad3 и p-Smad3 белков в собранных клетках.
Общий белок клеток экстрагировали, и концентрацию белка определяли на основе набора бицинхониновой кислоты. Образец белка загружали в лунки при электрофорезе в полиакриламидном геле с додецилсульфатом натрия и переносили на мембрану из поливинилиденфторида (PVDF). Мембрану PVDF блокировали обезжиренным молоком и инкубировали с первичными антителами TGFBR3 (1:2000, R&D Systems, Миннеаполис, Миннесота, США), t-Smad1 (1:1000), p-Smad1 (1:1000, Santa Cruz Biotechnology). , t-Smad3 (1:1000), p-Smad3 (1:1000) и GAPDH (1:1000, все из Abcam, Кембридж, Массачусетс, Великобритания) с последующей инкубацией с вторичными антителами, меченными пероксидазой хрена ( 1:500, Jackson ImmunoResearch Laboratories, Пенсильвания, США). Промывали 3 раза трис-буферным физиологическим раствором с Твин 20, мембрана проявляла усиленную хемилюминесценцию. Количественная оценка сигналов была завершена Национальным институтом здравоохранения ImageJ Imaging. Программное обеспечение для анализа обработки с интенсивностью сигналов, нормализованной к GAPDH.
Культура клеток и скрининг
Клеточные линии человеческого NPC CNE2, TW03, C666-1 и SUNE-1 и нормальная линия носоглоточных клеток человека NP96 (Шанхайский институт биологических наук Китайской академии наук, Шанхай, Китай) культивировали в Мемориальном институте парка Розуэлл (RPMI) - 1640 (Gibco, Калифорния, США), содержащая FBS (Gibco), 100 мкг / мл пенициллина и 100 мкг / мл стрептомицина, и пассировали при 80% конфлюэнтности. RT-qPCR использовали для обнаружения экспрессии miR-18a. Среди этих клеточных линий NPC клетки CNE2 и SUNE-1 показали наибольшее и наименьшее различие в экспрессии miR-18a от клеток NP96, поэтому они были отобраны для анализов понижающей или повышающей регуляции miR-18a.
Группировка и обработка ячеек
Среди всех линий клеток NPC были отобраны клетки SUNE-1 с наименьшим отличием от клеток NP96 по экспрессии miR-18a. Руководствуясь спецификациями Lipofectamine 2000 (Invitrogen), клетки SUNE-1 были трансфицированы имитаторами miR-18a, имитаторами miR-18a отрицательного контроля (NC), si-TGFBR3 или si-TGFBR3 NC.
Среди всех клеточных линий NPC клетки CNE2 с наибольшим отличием от клеток NP96 по экспрессии miR-18a были отобраны и трансфицированы ингибиторами miR-18a, ингибиторами miR-18a NC, сверхэкспрессией (OE) -TGFBR3 или OE-TGFBR3 NC липофектамином 2000. (Invitrogen).
Руководствуясь спецификациями Lipofectamine 2000 (Invitrogen), макрофаги M2 были трансфицированы миметиками miR-18a, миметиками miR-18a NC, si-TGFBR3, si-TGFBR3 NC, ингибиторами miR-18a, ингибиторами miR-18a NC, OE-TGFBR3, или OE-TGFBR3 NC.
Совместное культивирование макрофагов M2 и клеток NPC
Совместное культивирование клеток в камере Transwell было адаптировано для изучения эффектов miRNA из макрофага M2 на клетки NPC. Верхняя камера была заполнена макрофагами M2 с размером пор 0,4 мкм, что препятствовало прохождению только клеток верхней камеры, но не мелких молекул, секретируемых клетками, таких как везикулы, факторы роста, питательные вещества и т. Д. был распространен клетками NPC.
SUNE-1 и CNE2 инкубировали в нормальной FBS (Gibco). Клетки в фазе логарифмического роста были приняты для экспериментов.
Клетки SUNE-1 и CNE2 совместно культивировали с макрофагами M2 в 10% среде FBS-RPMI-1640 (оба от Gibco) в чашке для культивирования клеток Transwell Insert (Coring, Corning, NY, USA) с размером пор 0,4 мкм. .
Клетки SUNE-1 не культивировались совместно с макрофагами M2 и не культивировались совместно с макрофагами M2, miR-18a имитировали трансфицированные макрофаги M2, miR-18a имитировали NC-трансфицированные макрофаги M2, si-TGFBR3-трансфицированные макрофаги M2 или si -TGFBR3 NC-трансфицированные макрофаги M2.
Клетки CNE2 не культивировались совместно с макрофагами M2 или совместно с макрофагами M2, макрофагами M2, трансфицированными ингибиторами miR-18a, ингибиторами miR-18a, NC-трансфицированными макрофагами M2, макрофагами M2, трансфицированными OE-TGFBR3, или OE-TGFBR NC-трансфицированные макрофаги M2.
Анализ 3- (4, 5-диметилтиазол-2-ил) -2,5-дифенилтетразолийбромида
Жизнеспособность клеток тестировали с помощью 3- (4,5-диметилтиазол-2-ил) -2,5-дифенилтетразолийбромида (МТТ), который представлял собой колориметрический анализ, применяемый для определения активности митохондриальной дегидрогеназы, которая снижает МТТ до формазана.
Трипсинизировали и высевали в 96-луночные планшеты в количестве 4 × 10 4 . клеток / лунку, клетки лишали культуральной среды на 0, 12, 24, 36 и 48 час соответственно и добавляли раствор МТТ (500 мкл, 0,5 г / л). Инкубировали в течение 4 ч, супернатант отбрасывали и клетки инкубировали с 200 мкл раствора диметилсульфоксида. Значения оптической плотности (OD, 490 нм) измеряли на считывающем устройстве для микропланшетов (ELX808IU, BioTek, VT, USA). В каждой группе было по 6 параллельных лунок.
Анализ образования колоний
Колониеобразующая способность клеток NPC была протестирована с помощью анализа образования колоний, который отразил зависимость клеточной популяции и клеточную клональную пролиферацию.
После культивирования в течение 24 ч и отделения 0,25% трипсина 300 клеток высевали в чашку диаметром 35 мм с 3 параллельными лунками в каждой группе. При обновлении культуральной среды каждые 3 дня клетки культивировали в течение недели и фиксировали 5 мл 4% параформальдегида. После этого клетки окрашивали красящим раствором кристаллического фиолетового и сушили на воздухе. Чашку переворачивали, на которую накладывали прозрачную пленку из сеток и подсчитывали количество колоний (более 50 клеток) под микроскопом (Olympus, Токио, Япония).
Скретч-тест
Миграцию клеток проверяли скретч-тестом. Клетки обрабатывали трипсином, высевали в 6-луночные планшеты с 3 параллельными лунками для каждой группы и культивировали до 90% слияния. Затем клетки инкубировали в среде с 2% FBS и рисовали вертикальные царапины кончиком 100 мкл. Клетки фотографировали в 0 и 24 часа под инвертированным микроскопом для измерения расстояния миграции клеток.
Анализ Transwell
Инвазию и миграцию клеток проверяли с помощью анализа Transwell. Верхнюю камеру камеры Transwell предварительно погружали и добавляли 100 мкл Matrigel (Coring), разбавленного бессывороточной средой RPMI 1640 в соотношении 1:100. В верхнюю и нижнюю камеры по отдельности добавляли 200 мкл и 600 мкл бессывороточной среды RPMI 1640. Затем в нижнюю камеру добавляли 600 мкл среды RPMI 1640, содержащей 10% FBS, а в верхнюю камеру - 200 мкл клеточной суспензии (12,5 × 10 4 клеток / мл). После инкубации в течение 40 часов клетки окрашивали раствором для окрашивания кристаллическим фиолетовым и протирали ватным тампоном для подсчета клеток, прошедших через матригель, под микроскопом.
Проточная цитометрия
Апоптоз клеток и распределение клеточного цикла определяли методом проточной цитометрии.
Распределение клеточного цикла оценивали по окрашиванию йодидом пропидия (PI). Клетки высевали в 6-луночные планшеты в количестве 4 × 10 5 . клеток / лунку и культивировали до 70–80% конфлюэнции. Фиксировали в предварительно охлажденном 70% этаноле в течение ночи, клетки центрифугировали (супернатант отбрасывали), добавляли РНКазу (1 г / л, 200 мкл) и Triton X-100 (2 мкл) и окрашивали окрашивающим раствором PI для 30 минут. После этого распределение клеточного цикла определяли с помощью проточного цитометра (BD Bioscience, NJ, USA) при 488 нм в соответствии с различной интенсивностью флуоресценции клеток на каждой фразе (фраза G0 / G1, фраза S и фраза G2 / M).
Апоптоз клеток измеряли с помощью аннексина V-флуоресцеина изотиоцианата (FITC) и двойного окрашивания PI. Клетки ресуспендировали в 500 мкл связывающего буфера и окрашивали 5 мкл окрашивающего раствора аннексина V-FITC и 10 мкл раствора PI. Апоптоз клеток также тестировали с помощью проточного цитометра (BD Bioscience) в течение 30 минут без воздействия света. На диаграмме рассеяния живые клетки в нижнем левом квадранте (Q4) были FITC - / PI - , апоптотические клетки на ранней стадии в нижнем правом квадранте (Q3) были FITC + / PI - , а некротические и апоптотические клетки на поздней стадии в верхнем правом квадранте (Q2) были FITC + / PI + . Скорость апоптоза =процент раннего апоптоза (Q3) + процент позднего апоптоза (Q2).
Ксенотрансплантаты опухоли у голых мышей
Рост опухоли наблюдали путем создания модели NPC у мышей nude. Отделенные 0,25% трипсином, клетки SUNE-1 и CNE2 в логарифмической фазе были сконфигурированы в суспензию отдельных клеток при 5 × 10 7 клеток / мл. Суспензию клеток (0,2 мл) вводили в правую подмышку мышей с помощью микроинъектора для создания моделей мышей. Моделируемых мышей выращивали в среде, свободной от патогенов. Начиная с 4-го дня, наблюдали рост опухоли, и мышей взвешивали каждые 4 дня. Голых мышей умерщвляли на 20-й день после инъекции, и опухоли резецировали, взвешивали на электронных весах и фотографировали.
Анализ двойного люциферазного репортерного гена
Система двойного репортерного гена люциферазы была принята для подтверждения сайтов связывания miR-18a и 3'-нетранслируемой области (UTR) мРНК TGFBR3. Веб-сайт биологического прогнозирования (http://www.microrna.org/microrna/home.do) был использован для анализа целевого гена miR-18a и обнаружения существования комплементарного сайта связывания miR-18a на 3 ' UTR TGFBR3. Двойной анализ репортерного гена люциферазы использовали для дополнительной проверки того, является ли TGFBR3 прямым мишенью miR-18a. Были сконструированы pmirGLO-TGFBR3-дикого типа (WT) и pmirG-LO-TGFBR3-мутантный тип (MUT) сайта связывания 3'UTR TGFBR3. TGFBR3-WT или TGFBR3-MUT и miR-18a, имитирующие или имитирующие NC, котрансфицировали в клетки SUNE-1 и CNE2 с помощью Lipofectamine 2000 (Invitrogen) и инкубировали в течение 48 часов. Набор для анализа люциферазы (Promega, Мэдисон, Висконсин, США) использовали для анализа клеток.
Статистический анализ
Для анализа использовали статистическое программное обеспечение SPSS21.0 (IBM Corp. Armonk, NY, USA). Данные были выражены как среднее ± стандартное отклонение. Различия между двумя группами были проанализированы t проверить различия между несколькими группами с помощью одностороннего дисперсионного анализа (ANOVA) с последующим апостериорным тестом Тьюки. Существенная разница считалась P <0,05.
Результаты
Идентификация макрофагов M2
Моноциты, обогащенные адгезивным методом, и моноциты, индуцированные M-CSF, собирали из периферической крови здоровых доноров. Иммунофлуоресцентное обнаружение CD68, CD206 и CD163 подтвердило, что PBMC, индуцированные M-CSF in vitro, превратились в макрофаги с типичными молекулярными характеристиками, которые соответствовали нашим требованиям (рис. 1a).
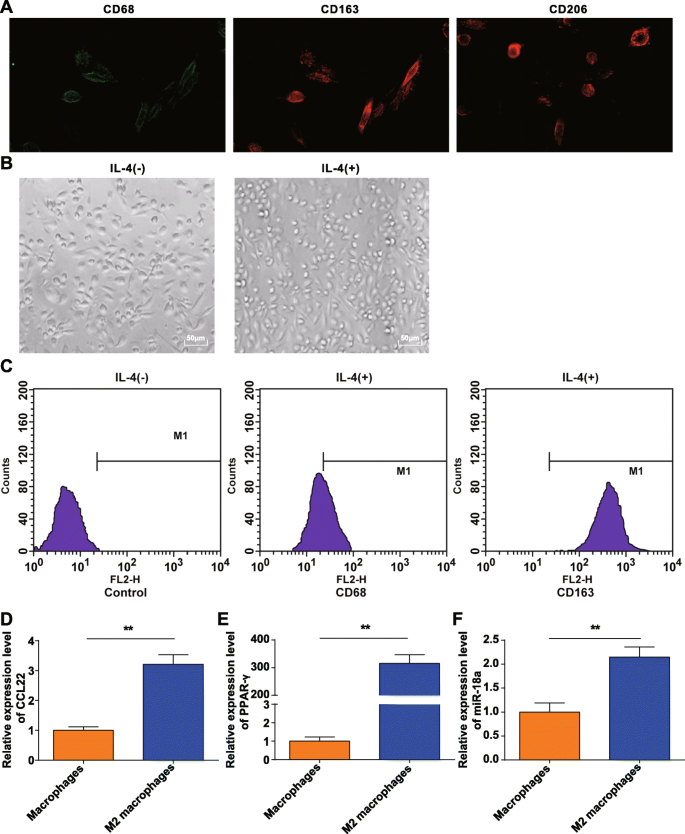
Идентификация макрофагов М2. а . CD68, CD206 и CD163 экспрессировались на поверхности макрофагов, полученных после индукции моноцитов in vitro. б Поляризованные IL-4 макрофаги были макрофагами M2. c CD163 был высоко экспрессирован, а CD68 низко экспрессирован в макрофагах M2, поляризованных IL-4. г CCL22 был высоко экспрессирован в макрофагах M2. е PPAR-γ был высоко экспрессирован в макрофагах M2. е miR-18a высоко экспрессируется в макрофагах M2; * P <0,05; ** P <0,01. Данные измерений были выражены как среднее значение ± стандартное отклонение, N =3. Сравнение двух групп было проанализировано t тест
Полученные макрофаги поляризовали IL-4 и наблюдали под микроскопом на предмет морфологии. Было обнаружено, что макрофаги (M0) без стимуляции IL-4 были разнообразными и нерегулярными, имели круглую, овальную или веретеновую форму. Стимулируемые IL-4, макрофаги M2 становились больше и приобретали преимущественно округлую форму, что соответствовало морфологическим характеристикам макрофагов M2, как описано ранее [20] (рис. 1b).
Проточная цитометрия проверила поверхностные антигены адгезивных клеток, стимулированных 20 нг / мл IL-4 в течение 24 часов, и обнаружила, что CD68 экспрессируется на уровне 21,16%, а CD163 - на уровне 98,69% от общего числа клеток (рис. 1c), что подтверждает адгезивные клетки представляют собой макрофаги М2. RT-qPCR показала, что с клетками M0, напротив, CCL22 и PPAR-γ (типичные поляризующие молекулы) увеличиваются в макрофагах M2 (рис. 1d, e), что указывает на успешную индукцию макрофагов M2.
RT-qPCR также показала, что экспрессия miR-18a увеличилась в макрофагах M2 в отличие от макрофагов M0 ( P ˂ 0,05) (рис. 1f).
MiR-18a сильно экспрессируется, а TGFBR3 плохо экспрессируется в ячейках NPC
Сайт биоинформатики (miRanda) предсказал потенциальные мишени для miR-18a, а TGFBR3 рассматривался как одна мишень для miR-18a (рис. 2a). Двойной анализ репортерного гена люциферазы был осуществлен для проверки того, что miR-18a нацелена на 3'UTR TGFBR3. TGFBR3-WT или TGFBR3-MUT клонировали в вектор pmirGLO и котрансфицировали имитаторами miR-18a или NC в клетки SUNE-1 и CNE2. Миметики miR-18a не влияли на активность люциферазы TGFBR3 3′UTR-MUT, но нарушали активность TGFBR3 3′UTR-WT в клетках SUNE-1 и CNE2, что позволяет предположить, что TGFBR3 был геном-мишенью, регулируемым miR-18a (рис. . 2b).
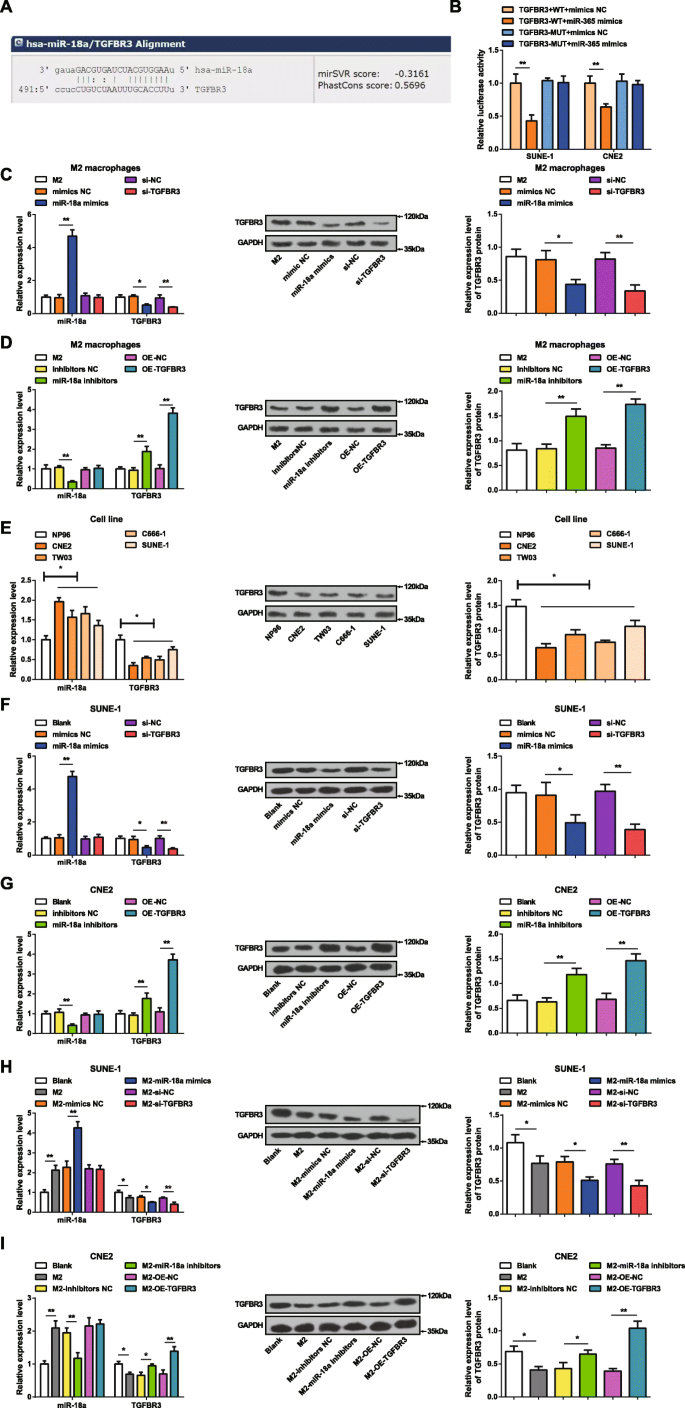
MiR-18a высоко экспрессируется, а TGFBR3 слабо экспрессируется в клетках NPC. а miRanda предсказал, что miR-18a нацелена на TGFBR3. б Двойной анализ репортерного гена люциферазы подтвердил, что miR-18a нацелен на TGFBR3. c Экспрессия miR-18a была повышена, а экспрессия TGFBR3 снижена в макрофагах M2, трансфицированных имитатором miR-18a. г Экспрессия miR-18a снижалась, а экспрессия TGFBR3 повышалась в макрофагах M2, трансфицированных ингибиторами miR-18a. е Экспрессия miR-18a была повышена, а экспрессия TGFBR3 снижена в клеточных линиях NPC по сравнению с клетками NP96. е Экспрессия miR-18a была повышена, а экспрессия TGFBR3 снижена в клетках SUNE-1, трансфицированных имитатором miR-18a. г Экспрессия miR-18a снижалась, а экспрессия TGFBR3 повышалась в клетках CNE2, трансфицированных ингибиторами miR-18a. ч Экспрессия miR-18a была повышена, а экспрессия TGFBR3 снижена в клетках SUNE-1, совместно культивируемых с макрофагами M2, трансфицированными имитаторами miR-18a. я Экспрессия miR-18a снижалась, а экспрессия TGFBR3 повышалась в клетках CNE2, совместно культивируемых с макрофагами M2, трансфицированными ингибиторами miR-18a; * P <0,05; ** P <0,01. Данные измерений были выражены как среднее значение ± стандартное отклонение, N =3. Сравнение между двумя группами было проанализировано t контрольная работа. Сравнения между несколькими группами были проанализированы с помощью однофакторного дисперсионного анализа с последующим апостериорным тестом Тьюки
Эффективность трансфекции имитаторов miR-18a или ингибиторов miR-18a в макрофагах M2 была продемонстрирована путем определения экспрессии miR-18a и TGFBR3 в макрофагах M2 с помощью RT-qPCR и Вестерн-блоттинга. Было очевидно, что (рис. 2c, d) избыточная экспрессия miR-18a повышает экспрессию miR-18a и снижает экспрессию TGFBR3 в макрофагах M2. Напротив, ингибирование miR-18a снижает экспрессию miR-18a и увеличивает экспрессию TGFBR в макрофагах M2. Не было обнаружено различий в экспрессии miR-18a, тогда как экспрессия TGFBR3 снижалась, когда макрофаги M2 трансфицировали si-TGFBR3. Не было обнаружено различий в экспрессии miR-18a, в то время как экспрессия TGFBR3 возрастала после трансфекции OE-TGFBR3 в макрофаги M2.
Экспрессию miR-18a и TGFBR3 в клеточных линиях CNE2, TW03, C666-1, SUNE-1 и NP96 тестировали с помощью RT-qPCR и Вестерн-блоттинга. В клетках NPC по сравнению с клетками NP96 экспрессия miR-18a увеличивалась, а экспрессия TGFBR3 снижалась (фиг. 2e). Поскольку клетки CNE2 и клетки SUNE-1 проявляют наибольшие и наименьшие различия в экспрессии miR-18a от клеток NP96, они были выбраны для продолжающихся анализов подавления и усиления miRNA соответственно.
Чтобы идентифицировать эффекты miR-18a и TGFBR3 на клетки NPC, клетки SUNE-1 трансфицировали имитатором miR-18a или si-TGFBR3, а клетки CNE2 - ингибиторами miR-18a или OE-TGFBR3. RT-qPCR и Вестерн-блоттинг продемонстрировали, что miR-18a имитирует повышенную экспрессию miR-18a и сниженную экспрессию TGFBR3 в клетках SUNE-1. Трансфекция si-TGFBR3 не влияла на экспрессию miR-18a, в то время как снижала экспрессию TGFBR3 в клетках SUNE-1. Ингибиторы miR-18a снижали экспрессию miR-18a и повышали экспрессию TGFBR3 в клетках CNE2. Трансфекция OE-TGFBR3 в клетки CNE2 не влияла на экспрессию miR-18a, но повышала экспрессию TGFBR3 (рис. 2f, g).
Чтобы изучить влияние miR-18a из макрофагов M2 на клетки NPC, макрофаги M2, имитирующие miR-18a или si-TGFBR3, или ингибиторы miR-18a, или макрофаги M2, трансфицированные OE-TGFBR3, культивировали совместно с SUNE-1. или клетки CNE2 в камере Transwell, соответственно. RT-qPCR и Вестерн-блоттинг тестировали экспрессию miR-18a и TGFBR3 в клетках SUNE-1 или CNE2. Клетки SUNE-1, совместно культивируемые с нетрансфицированными или имитирующими miR-18a макрофагами M2, продемонстрировали повышенную экспрессию miR-18a и сниженную экспрессию TGFBR3. Не было замечено различий в экспрессии miR-18a и снижении экспрессии TGFBR3 в клетках SUNE-1, которые культивировались совместно с макрофагами M2, трансфицированными si-TGFBR3 (фиг. 2h). После совместного культивирования с нетрансфицированными макрофагами M2 клетки CNE2 характеризовались повышенной экспрессией miR-18a и пониженной экспрессией TGFBR3. Однако сниженная экспрессия miR-18a и повышенная экспрессия TGFBR3 представлены в клетках CNE2, которые заранее совместно культивировали с макрофагами M2, трансфицированными ингибиторами miR-18a. Не было обнаружено различий в экспрессии miR-18a, а экспрессия TGFBR3 увеличивалась в клетках CNE2, совместно культивируемых с макрофагами M2, трансфицированными OE-TGFBR3 (рис. 2i).
miR-18a из макрофагов M2 повышает жизнеспособность клеток NPC и способность к колониеобразованию
Для определения влияния miR-18a и TGFBR3 на жизнеспособность и колониеобразующую способность клеток SUNE-1 и клеток CNE2 применялись МТТ-анализ и анализ образования колоний. Миметики miR-18a, ингибиторы miR-18a, si-TGFBR3 или OE-TGFBR3 трансфицировали в клетки SUNE-1 или клетки CNE2. Было показано, что в клетках SUNE-1 повышающая регуляция miR-18a или понижающая регуляция TGFBR3 повышали жизнеспособность клеток и увеличивали количество колоний (рис. 3a, e). В клетках CNE2 жизнеспособность клеток снижалась, а количество колоний снижалось из-за ингибирования miR-18a или сверхэкспрессии TGFBR3 (рис. 3b, f).
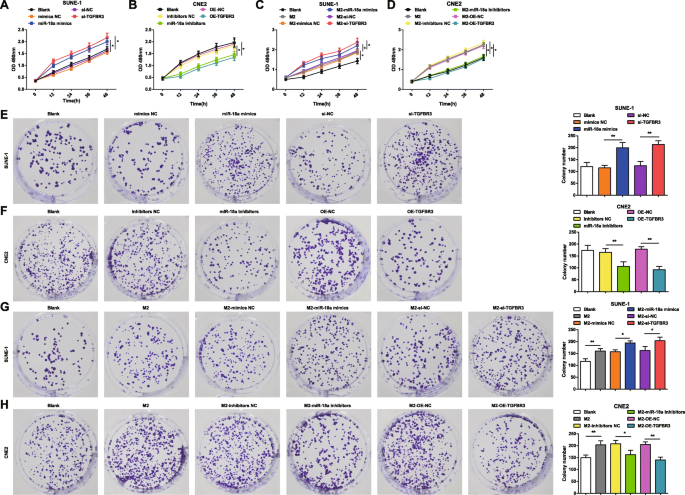
miR-18a из макрофагов M2 индуцирует жизнеспособность клеток NPC и способность к образованию колоний. а miR-18a имитирует или si-TGFBR3 увеличивает жизнеспособность клеток SUNE-1. б Ингибиторы miR-18a или OE-TGFBR3 снижают жизнеспособность клеток CNE2. c Совместное культивирование с макрофагами M2, имитирующими miR-18a или si-TGFBR3, увеличивало жизнеспособность клеток SUNE-1. г Совместное культивирование с ингибиторами miR-18a или макрофагами M2, трансфицированными OE-TGFBR3, снижает жизнеспособность клеток CNE2. е miR-18a имитирует или si-TGFBR3 увеличивает количество колоний клеток SUNE-1. е Ингибиторы miR-18a или OE-TGFBR3 уменьшали количество колоний клеток CNE2. г Совместное культивирование с макрофагами M2, имитирующими miR-18a или si-TGFBR3, увеличивало количество колоний клеток SUNE-1. ч Совместное культивирование с ингибиторами miR-18a или макрофагами M2, трансфицированными OE-TGFBR3, снижает количество колоний клеток CNE2; * P <0,05; ** P <0,01. Данные измерений были выражены как среднее значение ± стандартное отклонение, N =3. Сравнения между несколькими группами были проанализированы с помощью однофакторного дисперсионного анализа с последующим апостериорным тестом Тьюки
Чтобы изучить влияние miR-18a из макрофагов M2 на жизнеспособность и способность к колониеобразованию клеток NPC, совместно культивировали макрофаги M2, имитирующие miR-18a, ингибиторы miR-18a, si-TGFBR3 или OE-TGFBR3. с клетками SUNE-1 или клетками CNE2 в камере Transwell. Анализ МТТ и образования колоний показал, что клетки SUNE-1, совместно культивируемые с нетрансфицированными, или мимикой miR-18a, или si-TGFBR3-трансфицированными макрофагами M2, были представлены с повышенной жизнеспособностью клеток и увеличенным количеством колоний (рис. 3c, g) .
Клетки CNE2, культивированные совместно с нетрансфицированными макрофагами M2, были отмечены повышенной жизнеспособностью клеток и увеличением количества колоний. Однако клетки CNE2, совместно культивируемые с макрофагами M2, трансфицированными либо ингибиторами miR-18a, либо трансфицированными OE-TGFBR, демонстрировали снижение жизнеспособности клеток и уменьшение количества колоний (рис. 3d, h).
miR-18a из макрофагов M2 способствует вторжению и миграции клеток NPC
Для лучшего понимания того, как miR-18a и TGFBR3 влияют на миграцию и инвазию клеток NPC, были реализованы скретч-тест и анализ Transwell. Результаты показали, что клетки SUNE-1, трансфицированные миметиками miR-18a или si-TGFBR3, характеризовались увеличенным расстоянием миграции клеток и инвазией клеток (рис. 4a и 5a).
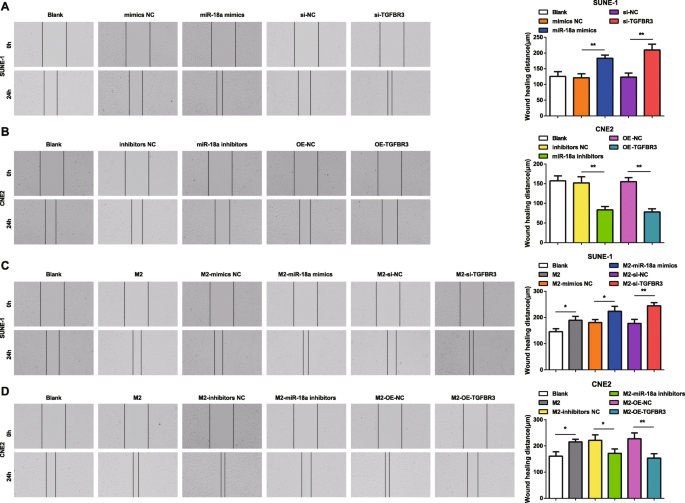
miR-18a из макрофагов M2 способствует миграции клеток NPC. а miR-18a имитирует или si-TGFBR3 увеличивает миграцию клеток SUNE-1. б Ингибиторы miR-18a или OE-TGFBR3 уменьшали миграцию клеток CNE2. c Совместное культивирование с макрофагами M2, имитирующими miR-18a или si-TGFBR3, увеличивало миграцию клеток SUNE-1. г Co-culture with miR-18a inhibitors or OE-TGFBR3-transfected M2 macrophages decreased migration of CNE2 cells; * P <0,05; ** P <0.01. Measurement data were expressed as mean ± standard deviation, N =3. Comparisons among multiple groups were analyzed by one-way ANOVA, followed by Tukey’s post hoc test

miR-18a from M2 macrophages promotes NPC cell invasion ability. а miR-18a mimics or si-TGFBR3 increased invasion of SUNE-1 cells. б miR-18a inhibitors or OE-TGFBR3 decreased invasion of CNE2 cells. c Co-culture with miR-18a mimics- or si-TGFBR3-transfected M2 macrophages increased invasion of SUNE-1 cells. б Co-culture with miR-18a inhibitors or OE-TGFBR3-transfected M2 macrophages decreased invasion of CNE2 cells; * P <0,05; ** P <0.01. Measurement data were expressed as mean ± standard deviation, N =3. Comparisons among multiple groups were analyzed by one-way ANOVA, followed by Tukey’s post hoc test
In CNE2 cells transfected with miR-18a inhibitors or OE-TGFBR3, the reductions appeared in cell migration distance and invasion cells (Figs. 4b and 5b).
How miR-18a from M2 macrophages influenced invasion and migration abilities of NPC cells were deciphered by miR-18a mimics-, miR-18a inhibitors-, si-TGFBR3-, or OE-TGFBR3-transfected M2 macrophages co-culturing with SUNE-1 or CNE2 cells. The results highlighted that SUNE-1 cells co-cultured with untransfected, or miR-18a mimics-transfected or si-TGFBR3-transfected M2 macrophages were manifested with increased cell migration distance and invasion cells (Figs. 4c and 5c).
Both cell migration distance and invasion cells increased in CNE2 cells co-cultured with untransfected M2 macrophages. Upon co-culture with miR-18a inhibitors-transfected or OE-TGFBR3-transfected M2 macrophages, CNE2 cells were showed with reduced migration distance and invasion cells (Figs. 4d and 5d).
miR-18a from M2 Macrophages Arrests Fewer NPC Cell at G0/G1 Phase and Suppresses Apoptosis
Cell cycle distribution and apoptosis were tested by flow cytometry to stratify the effects of miR-18a and TGFBR3 on NPC cells. It was indicated that transfection of miR-18a mimics or si-TGFBR3 reduced SUNE-1 cells arrested in the G0/G1 phase, increased cells in the S and G2/M phases, and reduced cell apoptosis rate (Figs. 6a and 7a).

miR-18a from M2 macrophages arrests fewer NPC cell at G0/G1 phase. а miR-18a mimics or si-TGFBR3 decreased SUNE-1 cells in the G0/G1 phase, and increased SUNE-1 cells in the S and G2/M phases. б miR-18a inhibitors or OE-TGFBR3 increased CNE2 cells in the G0/G1 phase, and decreased CNE2 cells in the S and G2/M phases. c Co-culture with miR-18a mimics- or si-TGFBR3-transfected M2 macrophages decreased SUNE-1 cells in the G0/G1 phase, and increased SUNE-1 cells in the S and G2/M phases. г Co-culture with miR-18a inhibitors or OE-TGFBR3-transfected M2 macrophages increased CNE2 cells in the G0/G1 phase, and decreased CNE2 cells in the S and G2/M phases; * P <0,05; ** P <0.01. Measurement data were expressed as mean ± standard deviation, N =3. Comparisons among multiple groups were analyzed by one-way ANOVA, followed by Tukey’s post hoc test

miR-18a from M2 macrophages inhibits NPC cell apoptosis. а miR-18a mimics or si-TGFBR3 decreased apoptosis of SUNE-1 cells. б miR-18a inhibitors or OE-TGFBR3 increased apoptosis of CNE2 cells. c Co-culture with miR-18a mimics- or si-TGFBR3-transfected M2 macrophages decreased apoptosis of SUNE-1 cells. г Co-culture with miR-18a inhibitors or OE-TGFBR3-transfected M2 macrophages increased apoptosis of CNE2 cells; * P <0,05; ** P <0.01. Measurement data were expressed as mean ± standard deviation, N =3. Comparisons among multiple groups were analyzed by one-way ANOVA, followed by Tukey’s post hoc test
Upon transfection with miR-18a inhibitors or OE-TGFBR3, CNE2 cells in the G0/G1 phase trended toward an elevation while those in the S and G2/M phases toward a reduction, and cell apoptosis rate raised (Figs. 6b and 7b).
With the purpose to decode the mechanism of miR-18a from M2 macrophages in NPC cell cycle distribution and apoptosis, M2 macrophages transfected with miR-18a mimics, miR-18a inhibitors, si-TGFBR3, or OE-TGFBR3 were co-cultured with SUNE-1 cells or CNE2 cells in the Transwell chamber. Co-cultured with untransfected, or miR-18a mimics-transfected or si-TGFBR3-transfected M2 macrophages, reduced SUNE-1 cells were displayed in the G0/G1 phase and increased cells in the S and G2/M phases, and SUNE-1 cell apoptosis rate decreased (Figs. 6c and 7c).
Co-cultured with untransfected M2 macrophages, CNE2 cells in the G0/G1 phase reduced, cells in the S and G2/M phases increased, and apoptosis rate declined. On the contrary, co-cultured with miR-18a inhibitors-transfected or OE-TGFBR3-transfected M2 macrophages, CNE2 cells in the G0/G1 phase elevated while those in the S and G2/M phases decreased, and apoptosis rate elevated (Figs. 6d and 7d).
miR-18a from M2 Macrophages Reduces p-Smad1/t-Smad1 and Elevates p-Smad3/t-Smad3 in NPC Cells
Western blot assay detected TGF signaling pathway-related proteins in NPC cells to further explain the effects of miR-18a and TGFBR3 on TGF signaling pathway.
It was explained that transfection of miR-18a mimics or si-TGFBR3 reduced p-Smad1/t-Smad1 and elevated p-Smad3/t-Smad3 in SUNE-1 cells (Fig. 8a).

miR-18a from M2 macrophages decreases p-Smad1/t-Smad1 and increases p-Smad3/t-Smad3 in NPC cells. а miR-18a mimics or si-TGFBR3 in SUNE-1 cells decreased p-Smad1/t-Smad1 and elevated p-Smad3/t-Smad3. б miR-18a inhibitors or OE-TGFBR3 in CNE2 cells increased p-Smad1/t-Smad1 and decreased p-Smad3/t-Smad3. c Co-culture with miR-18a mimics- or si-TGFBR3-transfected M2 macrophages decreased p-Smad1/t-Smad1 and elevated p-Smad3/t-Smad3. г Co-culture with miR-18a inhibitors or OE-TGFBR3-transfected M2 macrophages increased p-Smad1/t-Smad1 and decreased p-Smad3/t-Smad3; * P <0,05; ** P <0.01. Measurement data were expressed as mean ± standard deviation, N =3. Comparisons among multiple groups were analyzed by one-way ANOVA, followed by Tukey’s post hoc test
Transfection with miR-18a inhibitors or OE-TGFBR3 led to declined p-Smad3/t-Smad3 and increased p-Smad1/t-Smad1 in CNE2 cells (Fig. 8b).
miR-18a from M2 macrophages influencing TGF signaling pathway in NPC cells was determined by Western blot assay through testing TGF signaling pathway-related proteins in SUNE-1 cells and CNE2 cells which had co-cultured with miR-18a mimics-, miR-18a inhibitors-, si-TGFBR3-, or OE-TGFBR3-transfected M2 macrophages in the Transwell chamber.
SUNE-1 cells co-cultured with untransfected, miR-18a mimics-transfected, or si-TGFBR3-transfected M2 macrophages were manifested with reduced p-Smad1/t-Smad1 and incremental p-Smad3/t-Smad3 (Fig. 8c).
Co-cultured with untransfected M2 macrophages, CNE2 cells trended toward declined p-Smad1/t-Smad1 and elevated p-Smad3/t-Smad3. In an opposite way, CNE2 cells were demonstrated with increased p-Smad1/t-Smad1 and decreased p-Smad3/t-Smad3 when co-cultured with miR-18a inhibitors-transfected or OE-TGFBR3-transfected M2 macrophages (Fig. 8d).
miR-18a from M2 Macrophages Induces Tumor Growth in Nude Mice with NPC
Tumor xenografts were conducted on nude mice to further elucidate the impacts of miR-18a and TGFBR3 on tumor growth of NPC.
It was indicated that injected with miR-18a mimics-transfected or si-TGFBR3-transfected SUNE-1 cells, mice were manifested with enlarged tumor volume and heavier tumor weight (Fig. 9a).

miR-18a from M2 macrophages promotes tumor growth in nude mice with NPC. а miR-18a mimics or si-TGFBR3 in SUNE-1 cells increased tumor volume and weight. б miR-18a inhibitors or OE-TGFBR3 in CNE2 cells decreased tumor volume and weight. c Co-culture with miR-18a mimics- or si-TGFBR3-transfected M2 macrophages increased tumor volume and weight. г Co-culture with miR-18a inhibitors or OE-TGFBR3-transfected M2 macrophages decreased tumor volume and weight; * P <0,05; ** P <0.01. Measurement data were expressed as mean ± standard deviation, three nude mice in each group. Comparisons among multiple groups were analyzed by one-way ANOVA, followed by Tukey’s post hoc test
Reduced tumor volume and weight were presented in mice with injection of miR-18a inhibitors-transfected or OE-TGFBR3-transfected CNE2 cells (Fig. 9b).
Tumor growth was observed in mice which had injected with miR-18a mimics-, miR-18a inhibitors-, si-TGFBR3-, or OE-TGFBR3-transfected M2 macrophages to illustrate the mechanism of miR-18a from M2 macrophages in NPC.
After co-culture with untransfected, miR-18a mimics-transfected or si-TGFBR3-transfected M2 macrophages, SUNE-1 cells were injected into mice and mice were observed with larger tumor volume and heavier tumor weight (Fig. 9c).
CNE2 cells were co-cultured with M2 macrophages and injected into mice with the results suggesting growing tumor volume and weight. Both tumor volume and weight were inclined to reduce when mice were injected with CNE2 cells which had co-cultured with miR-18a inhibitors-transfected or OE-TGFBR3-transfected M2 macrophages (Fig. 9d).
Discussion
NPC refers to a polygenic disease threatened by a wide range of factors [21]. MiRNAs are previously implied to participate in the pathogenesis of NPC via regulation of their target genes which are indicators of cellular processes and pathways [22]. Concretely, miR-18a advances NPC progression by miRNA biogenesis impairing [6]. Given that this study goes forward to decipher the combined interactions of miR-18a from M2 macrophages and TGFBR3 in NPC with the conclusion elucidating that miR-18a from M2 macrophages stimulates NPC progression via TGFBR3 inhibition (Fig. 10).

Schematic representation of macrophage-derived exosomal miR-18a in NPC and the involvement of TGFBR3-mediated TGF-β signaling pathway
At the start of this study, macrophages are stimulated by IL-4 to differentiate to M2 macrophages which are found to enrich the expression of miR-18a. As we know, during macrophage polarization, miRNA’s expression was altered [23]. In addition, M2 polarization enriches genes which are involved in the cell cycle and metabolic processes, and the M2 phenotype is conducive to tumor growth and angiogenesis in neoplastic tissues [24]. Based on the M2 macrophages-enriched miR-18a, a series of experiments were successfully conducted.
Initially, our study has uncovered that miR-18a is highly expressed while TGFBR3 is poorly expressed in NPC cells. Drawn from a previous study, it is concluded that miR-18a is overexpressed in NPC tissues with its association with lymph node metastasis and clinical stage [5]. Besides that, miR-17-92 cluster members including miR-18a are documented to be overexpressed in NPC tissues [25]. Furthermore, upregulated miR-18a is reported to demonstrate in NPC tissues which is connected with tumor node metastasis stage and tumor size [4]. Experimentally, except for the downregulated TGFBR3 in tongue squamous cell carcinoma [26], there has been another study depicting reduced TGFBR3 in clear-cell renal cell carcinomas accompanied by unwanted prognosis [27]. Anyhow, the results in this study are consistent with these study findings to some extent.
In order to explore the roles of miR-18a and TGFBR3 in NPC cell progression, we have conducted a series of assays with the results indicating that upregulated miR-18a or downregulated TGFBR3 triggers NPC cell progression while miR-18a repression or TGFBR3 elevation has the opposite effects on NPC cells. Widely, suppression of miR-18a is evidenced to hamper cell progression in malignancies including ovarian cancer, colitis-associated colorectal cancer, and hepatocellular carcinoma [28,29,30]. Narrowly, an existed study has pronounced that upregulated miR-18a promotes NPC cell progression via mediation of DICER1 [6]. In addition, it is noticed that overexpressed miR-18a in NPC is believed to connect with NPC metastasis and repressed miR-18a partially contributes to better prognosis of NPC patients [31]. Lately, it is surveyed that downregulation of miR-18a is capable of discouraging NPC proliferation, invasion, and migration [4]. Additionally, a decrease in TGFBR3 expression is regarded to link with laryngeal squamous cell carcinoma (LSCC) invasion and miR-223/TGFBR3 axis regulation takes part in LSCC progression inhibition [32]. TGFBR3 elevation is documented to restrict NPC cell viability, induce apoptosis, and activate pro-apoptosis signaling pathways [14]. Previously, a study has indicated that upregulation of TGFBR3 promotes apoptosis and cells arrested in the G2/M phase, resulting in impaired cell viability and migration in salivary gland adenoid cystic carcinoma [33]. Intriguingly, it is formerly described that induction of TGFBR3 contributes to disrupt intrahepatic cholangiocarcinoma progression [34].
Despite the protective role of lowly expressed miR-18a and overexpressed TGFBR3 in NPC cell in vitro, we have performed tumor xenografts in nude mince in vivo for further verification with the results explaining that miR-18a knockdown or TGFBR3 elevation restrains tumor growth in nude mice. As demonstrated in a prior study, miR-18a-injected nude mice show with enhanced tumor growth [5] and conversely, the miR-18a antagomir-injected nude mice are displayed with suppressed tumor growth in NPC [4]. In the light of the TGFBR3 reduction in tumor growth, it is suggested that poorly expressed TGFBR3 provokes tumor formation in clear-cell renal cell carcinoma [27]. In the opposite way, an increase in TGFBR3 is recognized to hinder tumor growth in lung cancer with the presence of long non-coding RNA ADAM metallopeptidase with thrombospondin type 1 motif, 9 antisense RNA 2 elevation, and miR-223-3p suppression [35]. This study has also predicted and verified that TGFBR3 is a target gene of miR-18a. But, more studies still need to be conducted for further verification.
Conclusion
Generally speaking, this study elaborates the concrete mechanisms that miR-18a from M2 macrophages inhibits TGFBR3 expression to exacerbate the progression of NPC via TGF-β signaling pathway, the results of which is abrogated by miR-18a knockdown or TGFBR3 elevation. This study updates the therapeutic target for NPC. However, a large cohort researches are still in need for in-depth explorations.
Доступность данных и материалов
Not applicable.
Наноматериалы
- Гибридный композит на основе наноструктурированного диоксида кремния / золота и целлюлозы с амино-POSS, получе…
- Повышение противоопухолевой эффективности и фармакокинетики буфалина с помощью пегилированных липосом
- Получение полимерных наносфер палладия (II) с ионным отпечатком и удаление палладия (II) из водного раствора
- Наносборки 5-аминолевулиновой кислоты и сквалена для фотодетекции и терапии опухолей:исследования in vitro
- Стимуляция роста клеток SH-SY5Y наночастицами золота, модифицированными 6-меркаптопурином и проникающим в нейро…
- Наночастицы маггемита действуют как нанозимы, улучшая рост и устойчивость к абиотическому стрессу у Brassica napus
- Синтез и характеристика модифицированного BiOCl и их применение при адсорбции низкоконцентрированных красите…
- Материал и оптические свойства флуоресцентных углеродных квантовых точек, полученных из лимонного сока поср…
- Самосеянный рост MOCVD и значительно усиленная фотолюминесценция нанопроволок InGaAs / InP Core – Shell
- Доставка модифицированных магнитных наночастиц Fe3O4 для CpG ингибирует рост опухоли и спонтанные метастазы в л…