


Влияние наночастиц золота на метаболизм тестостерона в микросомах печени человека
Аннотация
Комплексы короны золотые наночастицы (AuNP) и белок могут изменять метаболизм тестостерона (TST), опосредованный цитохромом P450 (CYP), изменяя их физико-химические свойства. Мы исследовали влияние размера наночастиц, химии поверхности и белковой короны на метаболизм TST в объединенных микросомах печени человека (pHLM) с использованием AuNP 40 и 80 нм, функционализированных разветвленным полиэтиленимином (BPEI), липоевой кислотой (LA) и полиэтиленгликолем (PEG). ), а также корона белка плазмы человека (ПК). Индивидуальные вариации в AuNP-опосредованном метаболизме TST также были охарактеризованы среди HLM одного донора, который содержал разные уровни активности CYP. Ингибирующие эффекты 40 нМ AuNP и, в меньшей степени, 80 нМ AuNP проявлялись в отношении продукции в общей сложности пяти гидроксилированных метаболитов TST в pHLM, но PC ослабляли их. Между тем, голый AuNP увеличивал продукцию андростендиона. Индивидуальные вариации метаболизма TST наблюдались в пределах HLM одного донора. В большинстве случаев 40- и 80-нм голые и ПК-AuNP по существу подавляли метаболизм TST при неингибиторной концентрации, но ПК-PEG-AuNP повышали уровень андростендиона. Эти исследования способствуют лучшему пониманию роли AuNP как разрушителя TST, изменяя метаболизм TST, и могут быть использованы для проверки других NP как потенциальных эндокринных разрушителей.
Введение
Золотые наночастицы (AuNP) широко используются в доставке лекарств, медицинской диагностике и тераностике рака, а также в потребительских товарах, например, в косметике, упаковке пищевых продуктов, благодаря своим уникальным оптическим и физическим свойствам [1,2,3]. При воздействии смеси белков NP связываются с белками и образуют белковую корону, которая изменяет химический состав поверхности, конформации адсорбированного белка и последующие биологические ответы, например, токсичность NP, поглощение NP клетками, каталитическую активность цитохрома P450 (CYP ) ферменты по отношению к лекарствам [4,5,6,7]. Исследования in vitro с первичными эпителиальными клетками и линиями раковых клеток показывают, что AuNP токсичен для гепатоцитов человека, линии клеток гепатомы C3A и сперматозоидов [6,7,8]. Но образование белковой короны вокруг NP интенсивно ослабляло или усиливало токсичность AuNP в зависимости от химии поверхности [6, 7]. Белковая корона препятствовала клеточному поглощению AuNP в гепатоцитах человека, клетках проксимальных канальцев почек, клетках HepG2, клеточной линии C3A, независимо от их размеров и поверхностных зарядов [6, 7, 9,10,11,12].
Ферменты CYP печени в первую очередь участвуют в синтезе и / или метаболизме эндогенных и экзогенных соединений, но широкий спектр агентов, например, лекарства, пестициды или NP, обратным образом влияют на синтез, метаболизм и / или детоксикацию стероидных гормонов, что приводит к фармакологическим эффектам. и физиологическая функция [13,14,15,16,17]. Тестостерон (TST) является важным андрогеном и CYP3A4-специфическим субстратом (основное преобразование в 6β-OH TST) регио- и стереоселективным образом [18]. Во время фазы I метаболизма TST также гидроксилируется до 2β-OH TST с помощью CYP3A4 и деалкилируется до андростендиона (AD) с помощью CYP2D6 [17, 19]. Исследования in vitro с гепатоцитами человека, клеточной линией C3A, микросомами печени человека (HLM) и рекомбинантными ферментами CYP показали, что AuNP, покрытый белковой короной, модулирует широкий спектр ферментов CYP, включая CYP1A2, 2C9, 2C19, 2D6, 2E1, и 3A4 [6, 7, 20, 21]. Другой металлический NP, голый AgNP, также подавлял опосредованную CYP3A4 продукцию 6β-OH TST в HLM [22]. AuNP, функционализированный разветвленным полиэтиленимином (BPEI) и липоевой кислотой (LA), снижал активность CYP3A4 в линии клеток C3A, но белок короны плазмы крови человека (PC) ослаблял ее [7]. Напротив, голый (без PC) и PC BPEI-AuNP ингибировали CYP2C9 и 3A4 в гепатоцитах человека, независимо от размера NP [6].
Исследования in vivo показали, что небольшой размер AuNP (4 и 13 нм) накапливался в основном в печени и селезенке у самцов мышей BALB / c и индуцировал экспрессию печеночных генов Cyp1a1 и 2b [23]. Другие металлические НЧ, НЧ оксида цинка, ингибировали активность печеночных CYP1A2, 2C11 и 3A2 у самцов крыс Sprague Dawley с увеличением патологических изменений в печени [24].
На сегодняшний день мало что известно, как AuNP связывает опосредованный CYP метаболизм TST (гидроксилирование и деалкилирование TST) в отсутствие и / или в присутствии биологически релевантной белковой короны. Цели данного исследования - изучить влияние ПК на физико-химические свойства катионных BPEI AuNP с длиной волны 40 и 80 нм, анионного LA AuNP и нейтрального полиэтиленгликоля (PEG) AuNP. Влияние AuNP на CYP-опосредованный метаболизм TST с ПК и без него будет охарактеризовано с помощью pHLM. Индивидуальные вариации метаболизма TST будут также изучены в HLM одного донора, который содержал ферменты CYP в разной степени.
Методы / экспериментальные
Химические вещества
2,3,4- 13 С 3 тестостерон (CAS № 327048-83-9) и 6β-гидрокситестостерон (6β-OH TST, CAS № 62-99-7) были получены от MilliporeSigma (Сент-Луис, Миссури). Тестостерон (TST, CAS # 58-22-0), 2α-гидрокситестостерон (2α-OH TST, CAS # 004075-14-3), 2β-гидрокситестостерон (2β-OH TST, CAS # 10390-14-4), 6α -гидрокситестостерон (6α-OH TST, CAS № 2944-87-8), 11β-гидрокситестостерон (11β-OH TST, CAS № 1816-85-9), 15β-гидрокситестостерон (15β-OH TST, CAS № 39605-73- 7), 16α-гидрокситестостерон (16α-OH TST CAS # 63-01-4), 16β-гидрокситестостерон (16β-OH TST, CAS № 17528-90-4), 11-кетотестостерон (CAS № 564-35-2) , андростендион (AD, CAS # 63-05-8), 4-гидроксиандростендион (CAS # 566-48-3) и 11β-гидроксиандростендион (CAS # 382-44-5) были приобретены у Steraloids (Newport, RI ). Ацетонитрил и муравьиная кислота степени чистоты для ЖХ-МС были получены от Fisher Scientific (Fair Lawn, NJ), а сверхчистая вода была произведена на собственном производстве с помощью системы Synergy® UV-R от Merck KGaA (Дармштадт, Германия). Если не указано иное, все остальные реагенты были приобретены у MilliporeSigma (Сент-Луис, Миссури).
Микросомы печени человека
Объединенные микросомы печени человека (pHLM) (200 доноров, 100 мужчин и 100 женщин) и микросомы печени одного донора были получены от Corning Inc. (Шарлотт, Северная Каролина). Поставщик объединяет pHLM, но не пул одного донорского HLM. Характеристики и активность выбранного фермента цитохрома P450 (CYP) одного донорного HLM, использованного в этом исследовании, представлены в дополнительном файле 1:Таблица S1.
Синтез наночастиц золота
Biopure ™ 40- и 80-нм сферический AuNP, функционализированный катионным разветвленным полиэтиленимином (BPEI), анионной липоевой кислотой (LA) и нейтральным полиэтиленгликолем (PEG), были приобретены у nanoComposix (Сан-Диего, Калифорния). Основные материалы были синтезированы путем восстановления гидрата тетрахлораурата (III) водорода (HAuCl 4 3H 2 O) в водном растворе карбоната калия и подверглись процессу старения и фильтрации в тангенциальном потоке (TFF). Поверхность AuNP функционализировали с помощью LA или PEG путем добавления дигидролипоевой кислоты (0,2:1, w / w ) или ПЭГ с концевыми тиолметоксигруппами (Laysan Bio Inc., Arab, AL) (0,5:1, w / w ) соответственно с промывкой TFF и стерильной фильтрацией. Поверхности, функционализированные BPEI, были синтезированы с помощью химии EDC / NHS путем связывания карбоновой кислоты LA с аминами BPEI. Несвязанный BPEI удаляли промывкой TFF и последующим центрифугированием.
Приготовление коронок из белка плазмы крови человека
Объединенная человеческая плазма (HP, n =5) был получен от Biological Specialty Corp. (Кольмар, Пенсильвания). AuNP 40 и 80 нм инкубировали с человеческой плазмой при физиологическом объеме плазмы от общего объема крови, 55% ( v / v ) в орбитальном инкубаторе с встряхиванием / вращением при 37 ° C и 250 об / мин в течение 1 часа. В конце инкубации белки короны (PC) плазмы крови человека, окружающие NP, собирали центрифугированием при 20000 × g при 20 ° C в течение 20 минут с последующими тремя промывками фосфатно-солевым буфером (PBS). Несвязанные и неплотно связанные белки отбрасывали центрифугированием. Полученный PC AuNP диспергировали в PBS и использовали для характеристики физико-химических свойств и его взаимодействия с TST.
Физическая характеристика AuNP
Размер частиц и свойства поверхности измеряли с помощью динамического рассеяния света (DLS) и просвечивающей электронной микроскопии (TEM). Гидродинамические диаметры (D H ) и дзета-потенциал 40- и 80-нм голого (без PC) BPEI-, LA- и PEG-AuNP в деионизированной (DI) воде и PC AuNP в PBS измеряли с помощью Zetasizer Nano-Zs (Malvern Instruments, Вустершир, Великобритания) в 0 ч при 25 ° C. D H , индекс полидисперсности (PDI) и дзета-потенциал были также получены для голого и PC AuNP в микросомальном инкубационном буфере (pH 7,4) при 0 мин и 45 мин при 37 ° C. Образцы были измерены пять раз с 11 частями по 10 с каждый. ПЭМ охарактеризовала морфологию голых и ПК AuNP. Все AuNP помещали на покрытые формваром медные сетки и просматривали на Tecnai G2 Spirit BioTWIN с детектором Oxford (FEI Company, Hillsboro, OR) при ускоряющем напряжении 120 кВ. Комплект для микроскопии GATAN (GATAN Inc., Плезантон, Калифорния) измерял диаметры AuNP. Спектры оптического поглощения измеряли с помощью многорежимного микропланшетного ридера Spectra Max i3 (Molecular Devices, Саннивейл, Калифорния).
Метаболизм тестостерона in vitro в отсутствие и в присутствии голого и ПК AuNP
Предварительные исследования были проведены для определения времени инкубации и концентрации микросомального белка, чтобы обеспечить линейную скорость метаболизма для TST (конечная концентрация 10 мкМ). Продукция метаболитов TST была линейной от 1,3 до 9,3 мг микросомального белка / мл -1 до 60 мин. Метаболические анализы были выполнены, как полностью описано [25]. Вкратце, pHLM в микросомальном инкубационном буфере обрабатывали 10 мкМ TST, а затем добавляли AuNP без покрытия 40 и 80 нм (без PC) при 0, 7, 32, 63, 143, 250, 400 и 571 мкг / мл -1 ; для ПК AuNP pHLM 0, 7, 32, 63 и 143 мкг мл -1 . Буфер для микросомальной инкубации содержал 100 мМ фосфатный буфер, 3,3 мМ MgCl 2 . и 1 мМ EDTA (pH 7,4). Метаболическая реакция была инициирована с регенерирующей системой НАДФН и без нее, которая содержала 0,25 мМ НАДФ, 2,5 мМ глюкозо-6-фосфат и 2 ед. Мл -1 глюкозо-6-фосфатдегидрогеназа. После 45-минутной инкубации при 37 ° C реакцию останавливали добавлением 4% ( v / v ) водный раствор фосфорной кислоты (1:1, v / v ). После центрифуги при 3500 об / мин в течение 20 мин образец супернатанта собирали и хранили при -20 ° C до дальнейшего использования. Кроме того, HLM одного донора в микросомальном инкубационном буфере обрабатывали 10 мкМ TST с последующей инкубацией с 63 мкг мл -1 всех голых и ПК AuNP в течение 45 мин при 37 ° C. По окончании инкубации образец обрабатывали и хранили при -20 ° C, как указано выше.
Стандарты и подготовка образцов
Основные стандартные исходные растворы TST, его метаболитов и 13 С 3 Меченые TST в качестве внутреннего стандарта (ISTD) получали в метаноле в концентрации 1 мМ и хранили при -20 ° C до дальнейшего использования. Концентрации рабочих стандартных растворов TST и его метаболитов составляли 0,01, 0,05, 0,1, 0,5, 1, 5, 10, 50, 100 и 200 мкМ с серийными разведениями основного исходного раствора. Для стандартных калибраторов аликвоту 50 мкл каждого рабочего стандартного раствора добавляли к 450 мкл реакционного буфера, что приводило к разведению 1:10, в то время как 0,1 мкМ раствора ISTD также готовили с использованием 4% водного раствора фосфорной кислоты. Образцы для контроля качества (QC) были приготовлены в концентрациях 0,01, 0,05 и 0,1 мкМ.
После оттаивания образцы подвергали центрифугированию при 3500 об / мин в течение 20 мин при комнатной температуре. В супернатант добавляли 50 мкл 0,1 мкМ ISTD и подвергали его обработке 96-луночным планшетом для микроэлюирования Oasis PRIME HLB и планшетом для сбора в процессоре положительного давления Waters-96 при 80 фунт / кв. Дюйм в течение 1-2 мин. (Waters Corp., Милфорд, Массачусетс). После промывания 300 мкл воды 5% метанолом и элюирования 50 мкл смеси ацетонитрил / метанол (90/10, v / v ) полученный элюент разбавляли 50 мкл воды (конечный объем 100 мкл) и подвергали жидкостной хроматографии-масс-спектрометрии (LC-MS / MS).
Жидкостная хроматография-масс-спектрометрия
Все образцы разделяли на колонке Waters UPLC HSS T3 (2,1 × 50 мм, 1,8 мкм) с помощью системы жидкостной хроматографии Waters Acquity Ultra Performance с трехквадрупольным детектором (UPLC TQD) (Waters Corp., Милфорд, Массачусетс). Подвижные фазы A и B представляли собой 0,1% муравьиной кислоты в воде и 0,1% муравьиной кислоты в метаноле, соответственно. Метод градиентной ЖХ использовался при скорости потока 600 мкл мин -1 . в течение 0–8,4 мин. Градиент составлял 0–1 мин (30% B), 1–3 мин (до 50% B), 3–3,5 мин (50% B), 3,5–7 мин (до 80% B), 7–7,01 мин ( до 98% B), 7,01–7,5 мин (98% B) и 7,51–8,4 мин (30% B). Условия МС кратко описаны ниже. Источник ионизации работал с положительным электрораспылением (ESI + ) режим с капиллярным напряжением 4000 В; для температуры источника 150 ° C; и для температур десольватации 450 ° C. Расход газа десольватации (N 2 ), газовый конус (N 2 ) и газа столкновений (аргон) составляли 900 л · ч −1 , 100 л ч −1 , и 0,1 мл мин -1 , соответственно. Тип сканирования - мониторинг множественных реакций (MRM), время выполнения MS составляет 8,4 мин. Переходы MRM, использованные для анализа, суммированы в дополнительном файле 1:Таблица S2 и рис. 2. Объем впрыска составлял 2 мкл, и колонка поддерживалась при 50 ° C на протяжении всего анализа. Все методы количественного определения основывались на калибровочной кривой по семи точкам в диапазоне концентраций от 0,001 до 20 мкМ. Предел обнаружения (LOD) и предел количественного определения (LOQ) 0,001 мкМ и 0,005 мкМ были установлены для TST и целевых метаболитов.
Статистический анализ
Влияние диспергентов на D H и PDI голого и ПК на AuNP оценивались с помощью t Стьюдента тест с двусторонним распределением. Половина максимальной ингибирующей концентрации (IC 50 ) и половина максимальной концентрации активации (ЕС 50 ) AuNP в сторону продукции CYP-зависимых метаболитов TST в pHLM определяли путем подбора уравнения Хилла с переменным наклоном к наблюдаемым данным с использованием GraphPad Prism®. Односторонний дисперсионный анализ (ANOVA) был проведен с использованием GraphPad Prism® для оценки эффектов лечения AuNP на метаболизм TST в HLM одного донора. Когда эффекты были значительными, проводили множественное сравнение с помощью теста честной значимой разницы Тьюки (HSD) с 5% уровнем значимости. Коэффициент корреляции Пирсона ( r ) между CYP-активностью HLM одного донора и продукцией CYP-зависимых метаболитов TST определяли с помощью GraphPad Prism® версии 6.07 (Ла-Холья, Калифорния).
Результаты и обсуждение
Физико-химическая характеристика AuNP короны голого и человеческого белка плазмы
Влияние короны белков плазмы (ПК) плазмы человека на размер НЧ, поверхностный заряд и морфологию, а также на спектральные свойства было охарактеризовано с помощью DLS, ПЭМ и УФ-видимой спектроскопии (рис. 1). ПЭМ-изображения продемонстрировали, что все чистые (без ПК) и ПК AuNP, за исключением 40 и 80 нм ПК BPEI-AuNP, были монодисперсными с постоянным распределением по размерам и уникальными спектральными диапазонами в УФ-видимой области (520–557 нм) (рис. 1a, b). . Отчетливые PC вокруг AuNP в PBS также были обнаружены с помощью ПЭМ. Агрегация разветвленного полиэтиленимина (BPEI) -AuNP, покрытого 40 и 80 нм PC, в PBS при 0 мин при 25 ° C коррелировала множественные пики в распределении размеров и красные смещения спектров поглощения относительно голого BPEI-AuNP (рис. 1b). Гидродинамический диаметр (D H ) значения 40 и 80 нм PC BPEI-AuNP, растворенного в PBS при 0 мин при 25 ° C и в микросомальном буфере для инкубации при 0 и 45 мин при 37 ° C, не определялись с помощью DLS, наряду с множественными пиками в распределении по размерам. D H значения 40 нм голого BPEI- и LA-AuNP и ПК PEG-AuNP и 80 нм голого BPEI-AuNP в микросомальном инкубационном буфере существенно увеличились до 45 мин при 37 ° C, тогда как его значение уменьшилось для 80 нм ПК LA. -AuNP (таблица 1). Индекс полидисперсности (PDI) 40 нм без покрытия и ПК PEG-AuNP и 80 нм ПК PEG-AuNP увеличивался через 45 мин при 37 ° C. Кроме того, значения дзета (z) потенциала голого BPEI-AuNP с длиной волны 40 и 80 нм и голого PEG-AuNP с длиной волны 40 нм существенно снизились с течением времени. В предыдущем исследовании сообщалось, что AuNP (7 и 70 нм), связанный с микросомальными белками печени человека, изменяет характеристический максимум поглощения в УФ-видимом диапазоне [21]. Эти результаты были подтверждены недавними исследованиями в нашей лаборатории, показавшими, что ПК и корона человеческого сывороточного альбумина изменяли размер НЧ, красное смещение максимальной абсорбции и морфологию независимо от растворяющей среды и времени инкубации [6, 7, 10, 26]. Основываясь на изменениях физико-химических свойств PC-опосредованных NP и ферментативной функции активности CYP [6, 7], потенциальное влияние AuNP 40 и 80 нм на CYP-опосредованный метаболизм микросомальных TST печени человека было исследовано в присутствии более биологически соответствующий ПК.

Просвечивающие электронные микрофотографии ( a ) голый AuNP 40 и 80 нм в деионизированной воде и b ПК AuNP при 40 и 80 нм в PBS при 0 ч при 25 ° C, УФ-спектр поглощения (верхняя вставка) и динамическое распределение светорассеяния (нижняя вставка). Стрелки указывают на формирование ПК. BPEI разветвленный полиэтиленимин, LA липоевая кислота, ПЭГ полиэтиленгликоль, ND не определено, ПК корона белка плазмы человека, голый нет ПК
AuNP-опосредованный метаболизм тестостерона в объединенных микросомах печени человека
Всего было проверено 11 метаболитов TST, и шесть метаболитов были обнаружены в объединенных микросомах печени человека (pHLM) при 10 мкМ TST. Направленный ЖХ-МС / МС анализ TST и шести выбранных метаболитов показан на рис. 2. Список выбранных метаболитов включал пять гидроксилированных метаболитов TST (2β-OH TST, 6β-OH TST, 15β-OH TST, 16α -OH TST, 16β-OH TST) и деалкилированный метаболит (андростендион, AD). Это коррелирует с предыдущими исследованиями с использованием гепатоцитов человека и HLM, согласно которым TST в основном гидроксилировался до 6β-OH TST и, в меньшей степени, 2β-OH TST, 15β-OH TST, 16α-OH TST и 16β-OH TST, а также деалкилированный метаболит, AD [17, 19, 27].
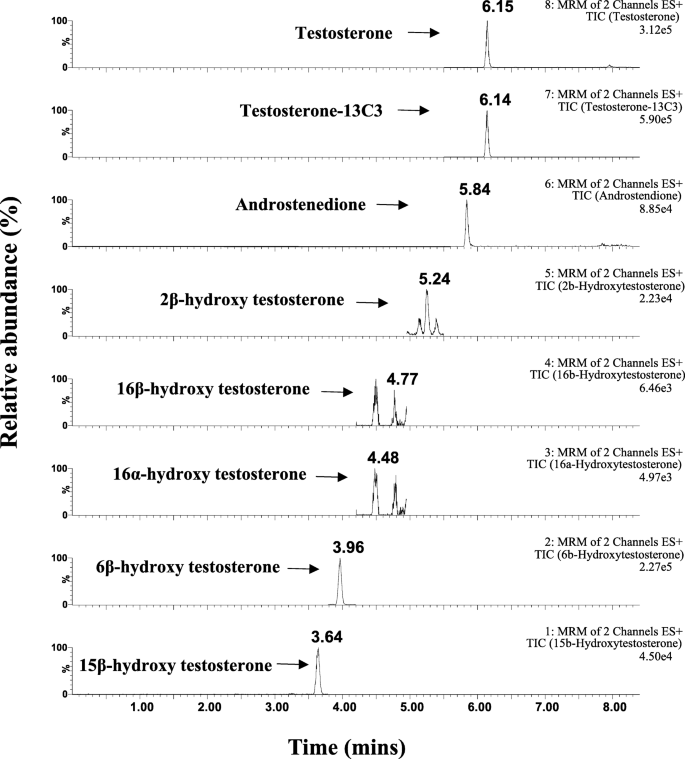
Хроматограмма экстрагированных ионов (XIC) для тестостерона (TST), 13 C3-меченный TST, андростендион, 2β-гидрокситестостерон (2β-OH TST), 16α-гидрокситестостерон (16α-OH TST), 16β-гидрокситестостерон (16β-OH TST), 6β-гидрокситестостерон (6β-OH TST) и 15β -гидрокситестостерон (15β-OH TST), продуцируемый в объединенном HLM при конечной концентрации 10 мкМ тестостерона в присутствии НАДФН в течение 45 мин при 37 ° C. HLM микросомы печени человека, НАДФН восстановленный никотинамидадениндинуклеотидфосфат
Результат совмещения TST с голым AuNP 40 и 80 нм в pHLM показан на рис. 3 и 4. Все 40- и 80-нм голые AuNP изменяли продукцию 2β-OH TST, 6β-OH TST и 15β-OH TST в pHLM с различной степенью ингибирования (рис. 3a – f). Половина максимальной ингибирующей концентрации (IC 50 ) значения 40 нм BPEI-AuNP для продукции 6β-OH TST составляли 416 мкг · мл -1 ; для 80 нм BPEI-AuNP 438 мкг мл -1 ; и для 80 нм ПЭГ-AuNP 387 мкг мл -1 (Рис. 3в, г). Для продукции 15β-OH TST, IC 50 значения 40 нм BPEI-AuNP составляли 1113 мкг мл -1 ; для 80 нм BPEI-AuNP 415 мкг мл -1 ; и для 80 нм ПЭГ-AuNP 480 мкг мл -1 (Рис. 3д, е). Эти результаты были подтверждены исследованиями in vitro с линиями раковых клеток человека и тканями печени, в которых металлические NP, AgNP и пористые кремниевые NP препятствовали продукции 6β-OH TST в клетках эпителиальной колоректальной аденокарциномы человека Caco2, клетках гепатоцеллюлярной карциномы HepG2 и микросомах печени человека. [22, 28]. Голый AuNP 40 и 80 нм не ингибировал продукцию 16α-OH TST и 16β-OH TST, за исключением 40 нм BPEI-AuNP, который подавлял продукцию 16β-OH TST в самой высокой концентрации (517 мкг / мл - 1 ) (Рис. 4a – d). 40 и 80 нм BPEI-AuNP увеличивали продукцию андростендиона (AD) в наивысшей концентрации с соответствующими скоростями метаболизма 4,3 и 2,1 пмоль мг белка -1 мин −1 соответственно по сравнению с контролем (0,7 пмоль мг белка -1 мин −1 ) (Рис. 4д, е). Но 80 нм LA-AuNP ингибировал продукцию AD с IC 50 значение 233 мкг мл -1 . Эти результаты показали, что голый AuNP опосредует продукцию выбранного метаболита TST в поверхностном покрытии и в зависимости от размера. Кроме того, все 40- и 80-нм PC AuNP не подавляли продукцию шести выбранных метаболитов из TST в pHLM при концентрации до 143 мкг / мл -1 , независимо от покрытия поверхности (рис. 5 и 6). В частности, ПК ослаблял опосредованное 40 нм голым BPEI-AuNP ингибирование продукции 6β-OH TST и 15β-OH TST при более высоких концентрациях (32 мкг мл -1 до 143 мкг мл -1 ) (Рис. 3в, д и 5в, д). Эти результаты показали, что 40- и 80-нм голые BPEI-, LA- и PEG-AuNP снижали гидроксилирование TST (2β-OH TST, 6β-OH TST и 15β-OH TST) дозозависимым образом (рис. 7). ). Кроме того, голый BPEI-AuNP с длиной волны 40 и 80 нм увеличивал продукцию AD, но первый уменьшал TST 16β-OH. Исследование in vitro показало, что продукция 6β-OH TST, в основном опосредованная CYP3A4, ингибируется однослойными углеродными нанотрубками (SWCNT) дозозависимым образом, но корона бычьего сывороточного альбумина смягчает это [17, 29]. Наша лаборатория недавно сообщила, что 40- и 80-нм голый и PC BPEI-AuNP служат ингибитором CYP1A2, 2C9 и 3A4 на клеточном и транскрипционном уровнях [6, 7]. Исследование in vivo показало, что PEG-AuNP (4 и 13 нм) в основном накапливается в печени самцов мышей BALB / c и изменяет уровни транскрипции печеночных генов Cyp1a1 и 2b [23]. Самцы мышей ICR с внутривенным введением инъекция ПЭГ-NH 2 -AuNP демонстрирует повышенный уровень ТКП в плазме NP без учета морфологии и фертильности сперматозоидов [30]. Другие металлические НП, диоксид титана (TiO 2 ) накапливалась в семенниках мышей-самцов CD1 и снижала экспрессию cyp1b1 и 2e1 [31]. Эпидемиологическое исследование показало, что взрослый мужчина в клинике бесплодия Массачусетса показал низкие уровни TST в плазме, сопоставимые с высоким уровнем 3,5,6-трихлор-2-пиридинола (TCP), вызванного сильным воздействием хлорпирифоса (CFS), известного эндокринного разрушителя и ингибитор CYP-опосредованного метаболизма TST [32]. В предыдущем исследовании сообщалось, что известные инсектициды, нарушающие работу эндокринной системы, CFS, CFS oxon, fonofos, форат, диэтилтолуамид (DEET) и перметрин существенно ингибируют и / или активируют продукцию гидроксилированных и / или деалкилированных метаболитов TST, т. Е. 2β-OH TST, 6β-OH TST, 15β-OH TST и AD и 4-гидрокси AD в печени человека [17]. С учетом вышесказанного, разумно предположить, что AuNP может быть потенциальным эндокринным разрушителем, опосредуя ингибирующие и / или активирующие способности против CYP-опосредованного метаболизма TST.
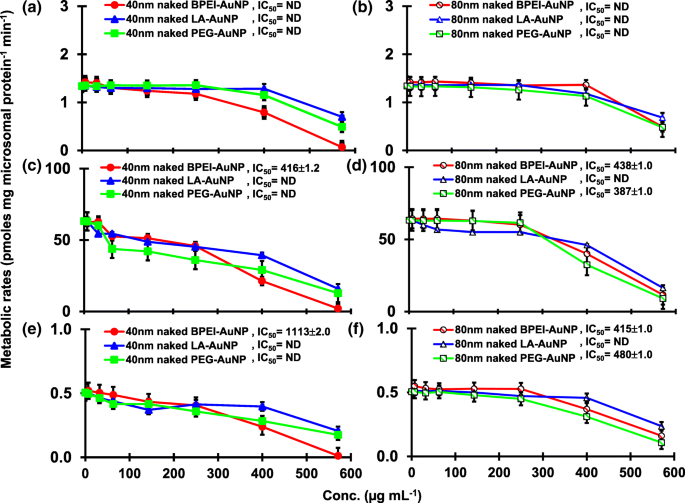
Ингибирующий эффект голого (без PC) AuNP на продукцию 2β-OH TST ( a , b ), 6β-OH TST ( c , d ) и 15β-OH TST ( e , f ) в pHLM. Данные представляют собой среднее значение ± стандартное отклонение. ( нет =3). IC 50 , половина максимальной ингибирующей концентрации; pHLM , объединенные микросомы печени человека; ND , не определяется путем подбора уравнения Хилла с переменным наклоном к наблюдаемым данным с использованием GraphPad Prism®; ПК , белок плазмы человека; BPEI , разветвленный полиэтиленимин; Лос-Анджелес , липоевая кислота; ПЭГ, полиэтиленгликоль; Conc, концентрация; 2β-OH TST, 2β -гидрокситестостерон; 6β-OH TST, 6β -гидрокситестостерон; 15β-OH TST, 15β -гидрокситестостерон
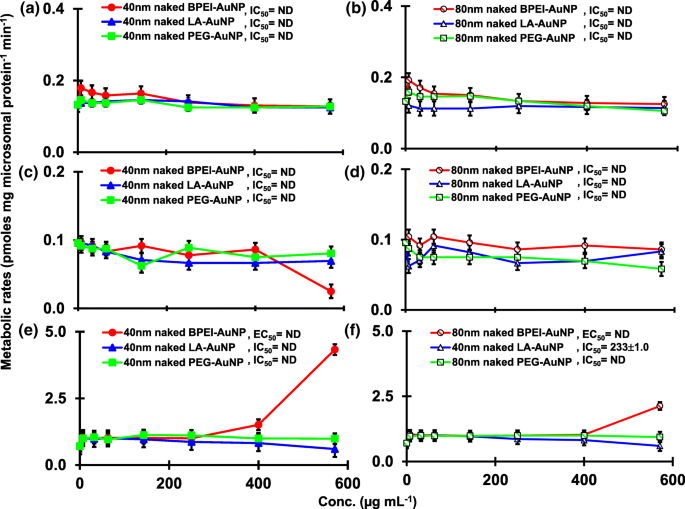
Ингибирующий и стимулирующий эффект голого (без PC) AuNP на продукцию 16β-OH TST ( a , b ), 16β-OH TST ( c , d ) и AD ( e , f ) в pHLM. Данные представляют собой среднее значение ± стандартное отклонение. ( нет =3). IC 50 , половина максимальной ингибирующей концентрации; EC 50 , половина максимальной активации конц .; pHLM , объединенные микросомы печени человека; ND , не определяется путем подбора уравнения Хилла с переменным наклоном к наблюдаемым данным с использованием GraphPad Prism®; ПК , белок плазмы человека; BPEI , разветвленный полиэтиленимин; Лос-Анджелес , липоевая кислота; ПЭГ , полиэтиленгликоль; Conc, концентрация; 16α-OH TST, 16α -гидрокситестостерон; 16β-OH TST, 16β -гидрокситестостерон; AD , андростендион.
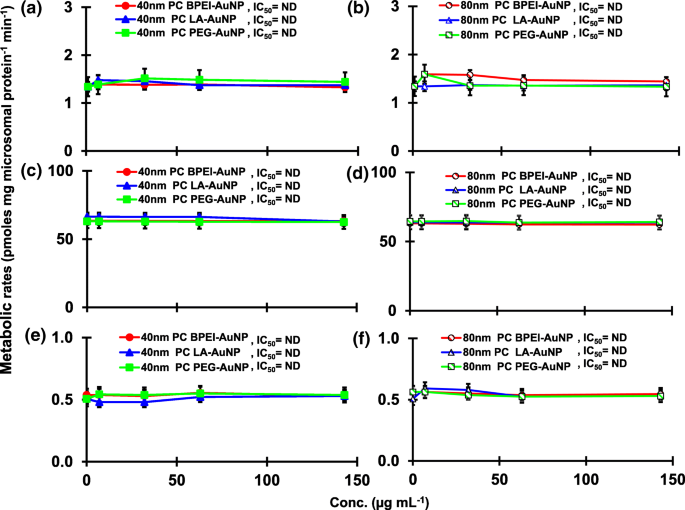
Влияние PC AuNP на продукцию 2β-OH TST ( a , b ), 6β-OH TST ( c , d ) и 15β-OH TST ( e , f ) в pHLM. Данные представляют собой среднее значение ± стандартное отклонение. ( нет =3). IC 50 , половина максимальной ингибирующей концентрации; pHLM , объединенные микросомы печени человека; ND , не определяется путем подбора уравнения Хилла с переменным наклоном к наблюдаемым данным с использованием GraphPad Prism®; ПК , корона белка плазмы человека; BPEI , разветвленный полиэтиленимин; Лос-Анджелес , липоевая кислота; ПЭГ , полиэтиленгликоль; Conc, концентрация; 2β-OH TST, 2β -гидрокситестостерон; 6β-OH TST, 6β -гидрокситестостерон; 15β-OH TST, 15β-гидрокситестостерон.
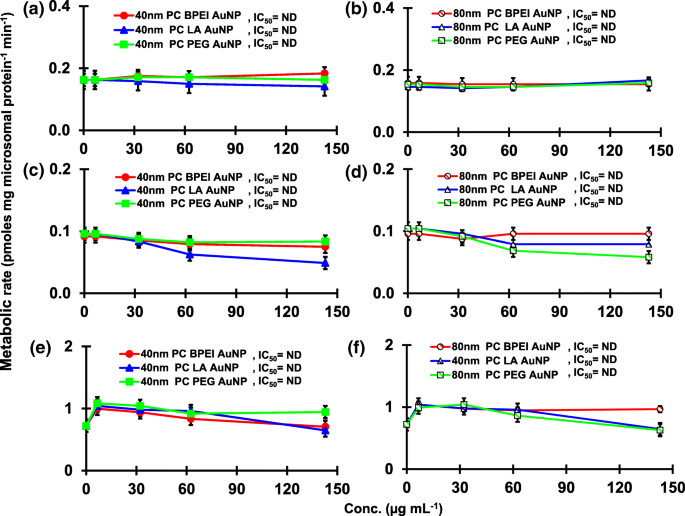
Влияние PC AuNP на продукцию 16α-OH TST ( a , b ), 16β-OH TST ( c , d ) и AD ( e , f ) в pHLM. Данные представляют собой среднее значение ± стандартное отклонение. ( нет =3). IC 50 , половина максимальной ингибирующей концентрации; pHLM , объединенные микросомы печени человека; ND , не определяется путем подбора уравнения Хилла с переменным наклоном к наблюдаемым данным с использованием GraphPad Prism®; ПК , корона белка плазмы человека; BPEI , разветвленный полиэтиленимин; Лос-Анджелес , липоевая кислота; ПЭГ , полиэтиленгликоль; Conc, концентрация; 16α-OH TST, 16α -гидрокситестостерон; 16β-OH TST, 16β -гидрокситестостерон; AD , андростендион.
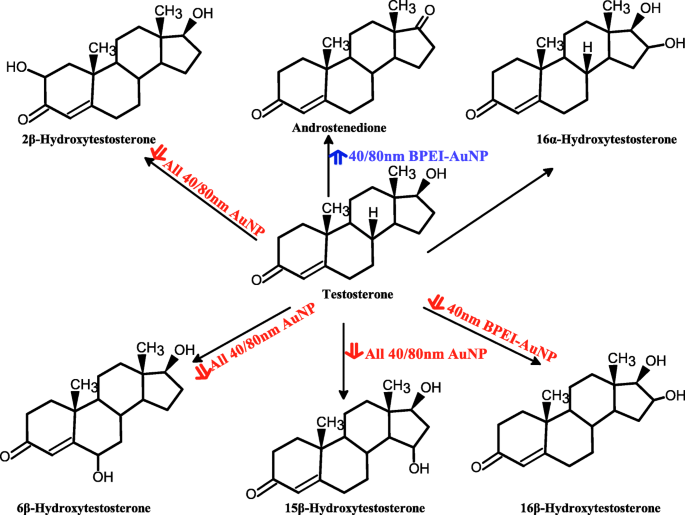
Предлагаемая схема метаболизма тестостерона в объединенных микросомах печени человека и продукции его метаболитов, опосредованной AuNP. AuNP наночастицы золота, BPEI разветвленный полиэтиленимин, LA липоевая кислота, ПЭГ полиэтиленгликоль, ПК протеин плазмы человека корона. Красная стрелка - тормозящее действие; синяя стрелка, стимулирующий эффект
Метаболизм TST в микросомах печени человека с одним донором и его модуляция с помощью AuNP
Продукция шести выбранных метаболитов была охарактеризована с помощью HLM одного донора, выделенного от доноров с различной степенью активности CYP при неингибиторной концентрации на основе pHLM 40 и 80 нм голого и PC AuNP (10 мкг мл -1 ). Индивидуальные вариации метаболизма TST наблюдались среди трех разных HLM одного донора (дополнительный файл 1:Рисунок S1). Взаимосвязь между каталитической активностью каждого фермента CYP и продуцированием метаболитов, полученных из TST, была охарактеризована в трех различных одиночных донорных HLM, которые содержали низкую, среднюю и высокую каталитическую активность CYP (дополнительный файл 1:таблица S1). Продукция 6β-OH TST положительно коррелировала с активностью CYP2C19 ( r =0,99 и p =0,01) и CYP3A4 ( r =0,99 и p =0,03) в пределах отдельных лиц (дополнительный файл 1:рисунок S2). Производство AD отрицательно коррелировало с CYP4A11 ( r =- 0,98 и p =0,04) (Дополнительный файл 1:Рисунок S3). Эти результаты согласуются с предыдущим исследованием, в котором сообщается, что CYP3A4 и CYP2D6 играют ключевую роль в производстве основного метаболита TST, 6β-OH TST и AD, соответственно [17]. Это исследование также показало, что индивидуальные вариации CYP-зависимого метаболизма TST зависят от генотипов ферментов CYP и их фенотипической активности. В предыдущем исследовании сообщалось, что полиморфизмы и фенотипы CYP являются ключевыми характеристиками функции CYP и приводят к таким категориальным фенотипам фармакогенетики, как плохие, промежуточные, экстенсивные и сверхбыстрые метаболизаторы, способствующие индивидуальной восприимчивости к побочным реакциям на лекарства и / или эффективности и дозировке препарата. что слабый метаболизатор фермента CYP, то есть CYP3A4, может быть восприимчив к метаболизму TST в результате воздействия ингибитора CYP, AuNP [33].
Как показано на фиг. 8 и 9, совпадение TST с AuNP в неингибирующей концентрации вызывало увеличение и / или снижение CYP-опосредованного метаболизма TST среди HLM одного донора в зависимости от модификации размера и изменения поверхности. ANOVA показал, что значительные изменения размера AuNP ( p <0,0001), поверхностные покрытия ( p <0,0001), и формирование ПК ( p <0,0001) наблюдались в отношении продукции шести выбранных метаболитов TST в HLM одного донора (HDA1, HDB2 и HDC3).

Effects of 40 nm naked and PC AuNP on the production of 2β-OH TST (A1–A3), 6β-OH TST (B1–B3), 15β-OH TST (C1–C3), 16α-OH TST (D1–D3), 16β-OH TST (E1–E3), and AD (F1–F3); in three different single donor HLM (HDA1, HDB2, and HDC3). Means followed by the same letter were not significantly different for each nanoparticle (Tukey’s honest significant difference =5%). Naked no PC, PC human plasma protein corona, HLM human liver microsomes, BPEI разветвленный полиэтиленимин, LA липоевая кислота, ПЭГ polyethylene glycol, 2β -OH TST 2β-hydroxytestosterone, 6β -OH TST 6β-hydroxytestosterone, 15β -OH TST 15β-hydroxytestosterone, 16α -OH TST 16α-hydroxytestosterone, 16β -OH TST 16β-hydroxytestosterone, AD androstenedione

Effects of 80 nm naked and PC AuNP on the production of 2β-OH TST (A1–A3), 6β-OH TST (B1–B3), 15β-OH TST (C1–C3), 16α-OH TST (D1–D3), 16β-OH TST (E1–E3), and AD (F1–F3); in three different single donor HLM (HDA1, HDB2, and HDC3). Means followed by the same letter were not significantly different for each nanoparticle (Tukey’s honest significant difference =5%). Naked no PC, PC human plasma protein corona, HLM human liver microsomes, BPEI разветвленный полиэтиленимин, LA липоевая кислота, ПЭГ polyethylene glycol, 2β -OH TST 2β-hydroxytestosterone, 6β -OH TST 6β-hydroxytestosterone, 15β -OH TST 15β-hydroxytestosterone, 16α -OH TST 16α-hydroxytestosterone, 16β -OH TST 16β-hydroxytestosterone, AD androstenedione
All 40 nm naked and PC AuNP decreased both 2β-OH TST and 6β-OH TST production in HDA1 and HDC3 except for PC PEG-AuNP in the former and naked LC-AuNP in the latter, whereas in HDB2, only PC AuNP potentiated inhibition of their productions, irrespective of surface coatings (Fig. 8(A1–B3)). The 40 nm naked LA-AuNP was an inhibitor for 15β-OH TST production in HDA1; for HDB2 PC PEG-AuNP; and for HDC3 naked BPEI- and PEG-AuNP and PC LA- and PC PEG-AuNP (Fig. 8(C1–C3)). All 40 nm naked AuNP were an activator for 16α-OH TST production in HDA1 but PC attenuated it except for PC BPEI-AuNP which potentiated its inhibition (Fig. 8(D1)). All 40 nm naked and PC AuNP did not influence the production of 16β-OH production within individuals (Fig. 8(E1–E3)). AD production was modulated by the 40 nm naked and PC AuNP with varying degrees of inhibition except for PC PEG-AuNP which served an activator in HDC3 (Fig. 8(F1–F3)).
The 80 nm naked and PC AuNP-mediated inhibition for 2β-OH TST production was observed within individuals except for 80 nm naked and PC PEG-AuNP which served as the activators in HDB2 and HDC3, respectively (Fig. 9(A1–A3)). These results were not consistent with all naked and PC 40 nm AuNP-mediated inhibition for 2β-OH TST, irrespective of surface coatings (Fig. 8(A1–A3)). For 6β-OH TST, the 80 nm naked AuNP were the activators except for LA-AuNP but PC potentiated its inhibition in HDB2 (Fig. 9(B2)), which was similar to the 40 nm naked and PC AuNP-mediated inhibition and/or activation, respectively (Fig. 8(B2)). For 15β-OH TST, 80 nm naked BPEI- and PEG-AuNP and PC PEG-AuNP were the activators in HDB2 and in HDC3, respectively (Fig. 9(C2 and C3)). The 80 nm naked AuNP increased 16α-OH TST production in HDA1, irrespective of surface coatings but PC attenuated it except for PC LA-AuNP which was an inhibitor (Fig. 9(D1)). This is similar to the 40 nm naked and PC AuNP-mediated activation and attenuation for 16α-OH TST production in HDA1 (Fig. 8(D1)). All 80 nm naked and PC were not inhibitory to 16β-OH TST production within all individuals, irrespective of surface coatings (Fig. 9(E1–E3)). These results were consistent with the 40 nm naked and PC-mediated its production within individuals (Fig. 8(E1–E3)). For AD production, the 80 nm naked BPEI- and PEG-AuNP were the inhibitors but PC attenuated and vice versa with naked and PC LA-AuNP in HDA1 (Fig. 9(F1)). The 80 nm naked and PC AuNP decreased its production in HDC3 except for PC PEG-AuNP, which was an activator (Fig. 9(F3)). This study strongly suggests that AuNP interaction with CYP enzymes in HLM cause a decrease and/or increase in TST conversion to hydroxylated and dealkylated metabolites within individuals and the presence of PC played the inhibitive or protective role. In vivo study reported that the male CD-1 mice orally administrated with ketoconazole, a noncompetitive CYP3A4 inhibitor showed that a decrease in serum TST level, gonadal TST secretion, and hepatic TST hydroxylation activity that included 6β-OH TST, 15α-OH TST, 15β-OH TST, and 16β-OH TST [34]. In vitro studies with human hepatocyte, C3A cell line, HepG2 cell line, HLM, and recombinant CYP enzymes suggested that AuNP modulated the activity of various CYP enzymes that included CYP1A2, 2C9, 2C19, 2D6, 2E1, and 3A4 [6, 7, 20, 21]. PC and human serum albumin corona mitigated an inhibitory effect of BPEI- and LA-AuNP on CYP1A2, 2C9, and 3A4 enzyme activity in human hepatocytes and C3A cell line [6, 7]. That being said, it may be rational to propose that AuNP interference with CYP enzymes relates individual susceptibility to unexpected toxicological effects that may result in an altered circulating TST level tied to endocrine disrupting substance and/or drug-drug interaction sharing the same CYP enzymes [35].
Conclusions
These studies exhibit that AuNP interaction with PC definitely modulate CYP-dependent metabolism of TST in HLM derived from a large donor pool that better represents the average American population. The 40 nm naked (no PC) AuNP and to a lesser degree 80 nm naked AuNP inhibited TST hydroxylation but activated TST dealkylation at high concentration. Cationic BPEI-AuNP withheld the production of 6β-OH TST and 15β-OH TST in pooled HLM but the presence of a more biologically relevant PC alleviated their adverse effects as function of size and surface charge modification. In most cases, the 40 and 80 nm naked and PC AuNP are essentially inhibitory to TST metabolism in single donor HLM in a surface chemistry-dependent manner at the noninhibitory concentration. In addition, PC PEG-AuNP caused an activation of AD production in HDC3, irrespective of size. These results may indicate that individual variations in AuNP-mediated TST metabolism could be a factor for their toxicity and could be utilized to identify vulnerable subgroup to TST-disrupting NP.
Доступность данных и материалов
All data generated or analyzed during this study are included in this article and its supplementary information file.
Сокращения
- 11β-OH TST:
-
11β-hydroxytestosterone
- 15β-OH TST1:
-
5β-hydroxytestosterone
- 16α-OH TST:
-
16α-hydroxytestosterone
- 16β-OH TST:
-
16β-hydroxytestosterone.
- 2α-OH TST:
-
2α-hydroxytestosterone
- 2β-OH TST:
-
2β-hydroxytestosterone
- 6α-OH TST:
-
6α-hydroxytestosterone
- 6β-OH TST:
-
6β-hydroxytestosterone
- AD:
-
Androstenedione
- AgNP:
-
Silver nanoparticles
- ANOVA:
-
One-way analysis of variance
- AuNP:
-
Наночастицы золота
- BPEI:
-
Branched polyethylenimine
- CFS:
-
Chlorpyrifos
- CYP:
-
Cytochrome P450
- DEET:
-
Diethyltoluamide
- DH :
-
Hydrodynamic diameters
- DI:
-
Деионизированная вода
- DLS:
-
Dynamic light scattering
- EC50 :
-
Half maximal activation concentration
- EDC/NHS:
-
1-Ethyl-3-(3-dimethylaminopropyl) carbodiimide/N-hydroxysuccinimide
- ESI + :
-
Electrospray positive
- HLM:
-
Human liver microsomes
- HSD:
-
Tukey’s honest significant difference
- IC50 :
-
Half maximal inhibitory concentration
- ISTD:
-
Internal standard
- LA:
-
Lipoic acid
- LC-MS/MS:
-
Liquid chromatography-mass spectrometry
- LOD:
-
Limit of detection
- LOQ:
-
Limit of quantitation
- MRM:
-
Multiple reaction monitoring
- NADP:
-
Nicotinamide adenine dinucleotide phosphate
- NADPH:
-
Reduced NADP
- naked:
-
No PC
- NP:
-
Наночастицы
- PBS:
-
Физиологический раствор с фосфатным буфером
- PC:
-
Human plasma protein corona
- PDI:
-
Polydispersity index
- PEG:
-
Полиэтиленгликоль
- pHLM:
-
Pooled human liver microsomes
- QC:
-
Quality control
- SWCNT:
-
Single-walled carbon nanotube
- ТЕМ:
-
Просвечивающая электронная микроскопия
- TFF:
-
Tangential flow filtration
- TiO2 :
-
Titanium dioxide
- TST:
-
Testosterone
- UPLC TQD:
-
Ultra performance liquid chromatography system with Triple quadrupole Detector
Наноматериалы
- Полупроводниковые наночастицы
- Плазмонные наночастицы
- Наночастицы золота для химиотерапевтических датчиков
- Многофункциональные наночастицы золота для улучшенных диагностических и терапевтических применений:обзор
- Золотой нанобиосенсор на основе локализованного поверхностного плазмонного резонанса способен диагностиро…
- Потенциальная токсичность наночастиц диоксида титана для печени, мозга и эмбрионов у мышей
- Повышенная стабильность магнитных наночастиц золота с помощью сополимера поли (4-стиролсульфоновой кислоты …
- Модифицированный гиперразветвленный полиглицерин как диспергатор для контроля размера и стабилизации нано…
- Стимуляция роста клеток SH-SY5Y наночастицами золота, модифицированными 6-меркаптопурином и проникающим в нейро…
- Татуировка из наночастиц золота произвела революцию в медицинской диагностике