


Наноскопия хондрогенно дифференцирующихся стволовых клеток человека, полученных из жировой ткани:наноструктура и экспрессия интегрина β1
на основе атомно-силовой микроскопии
Аннотация
Интегрин β1, как известно, участвует в дифференцировке, миграции, пролиферации, заживлении ран, развитии тканей и органогенезе. Чтобы проанализировать вероятность связывания между лигандом интегрина β1 и кластером дифференцированных рецепторов 29 (CD29), была использована атомно-силовая микроскопия (AFM) для обнаружения нативных интегриновых β1-связанных рецепторов на поверхности стволовых клеток, полученных из жировой ткани человека (hADSc). . Вероятность связывания взаимодействия интегрина β1-лиганд-рецептор исследовали с помощью функционализированных интегрином β1 наконечников на hADSc во время ранней хондрогенной дифференцировки на уровне двумерной культуры клеток. Морфология клеток и ультраструктура hADSc были измерены с помощью AFM, который продемонстрировал, что клетки с длинным веретеном превратились в полигональные клетки с уменьшенным соотношением длины / ширины и повышенной шероховатостью во время хондрогенной индукции. Связывание лиганда интегрина β1 и рецепторов CD29 детектировали с помощью β1-функционализированных наконечников для живых hADSc. Всего было записано 1200 кривых на 0, 6 и 12 день хондрогенной индукции. Средние силы разрыва составили соответственно 61,8 ± 22,2 пН, 60 ± 20,2 пН и 67,2 ± 22,0 пН. События разрыва составили 19,58 ± 1,74%, 28,03 ± 2,05% и 33,4 ± 1,89% соответственно, что продемонстрировало увеличение вероятности связывания между лигандом интегрина β1 и рецепторами на поверхности hADSc во время хондрогенной индукции. Интегрин β1 и сигнальный путь β-катенин / SOX коррелировали во время хондрогенной дифференцировки. Результаты этого исследования предполагают, что AFM предлагает кинетическое и визуальное понимание изменений в связывании интегрина β1-лиганда с рецептором CD29 на hADSc во время хондрогенеза. Было продемонстрировано, что изменения клеточной морфологии, ультраструктуры мембран и вероятность связывания лиганд-трансмембранный рецептор являются полезными маркерами для оценки процесса хондрогенной дифференцировки.
Фон
Остеоартрит (ОА) - распространенное дегенеративное заболевание суставов пожилых людей [1], при дегенеративном ОА, характеризующемся прогрессирующим разрушением суставного хряща. Хрящ высокоорганизован и не содержит кровеносных сосудов, нервов или лимфатической ткани [2]. Внеклеточный матрикс (ЕСМ) в основном состоит из коллагена II и гликопротеина и очень важен для гомеостаза хряща. Поскольку хрящ не имеет сосудов, его способность к самообновлению ограничена. Хотя лечение ОА (как хирургическое, так и нехирургическое) может быстро облегчить симптомы пациента с ОА, особенно боль, оно не может восстановить нормальную структуру и функцию суставного хряща [3]. В будущем лечение, вероятно, будет включать тканевую инженерию со стволовыми клетками и каркасами для восстановления дефектов и дегенеративного суставного хряща [4]. Мезенхимальные стволовые клетки - это мультипотентные стромальные клетки, которые обладают остеогенным, адипогенным, хондрогенным и миогенным потенциалом в зависимости от комбинации факторов роста [5]. Анализ дифференцировки мезенхимальных стволовых клеток показал, что Wnt / β-катенин, мишень рапамицина у млекопитающих (mTOR), фосфоинозитид-3-киназа (PI3K) и другие пути играют важную роль в дифференцировке [6,7,8]. Однако основной механизм, с помощью которого индуцируется хондрогенная дифференцировка, остается неуловимым. Это особенно верно для механизма, с помощью которого внеклеточные сигналы активируют внутриклеточные сигнальные пути. Мы обнаружили, что интегрин β1 претерпевает изменения во время хондрогенной дифференцировки. Таким образом, мы предположили, что интегрин β1 может играть важную роль в хондрогенной дифференцировке человеческих жировых стволовых клеток (hADSc) из-за его участия в различных сигнальных путях тканевой дифференцировки. В этом исследовании основное внимание уделялось сигнальному пути Wnt / β-катенин.
Многочисленные исследования показали, что взаимодействия между клетками и внеклеточной средой регулируются трансмембранными белками, в частности членами семейства интегринов [9]. Интегрины состоят из гетеродимерных трансмембранных гликопротеинов нековалентно связанных α- и β-цепей [10]. Теоретически известно 64 интегрина, из которых только 24 были обнаружены. Интегрины играют жизненно важную роль в межклеточной адгезии, межклеточной адгезии, передаче сигналов между клетками и организации актинового цитоскелета [11]. ЕСМ играет важную роль в гомеостазе тканей, и что ЕСМ регулирует интегрины. Интегрины опосредуют множество фундаментальных процессов, включая клеточную адгезию, миграцию, пролиферацию, дифференциацию, гибель клеток, заживление ран, развитие тканей и органогенез. Во время хондрогенной дифференцировки мезенхимальных стволовых клеток экспрессия интегрина β1 связана с сигнальным путем SOX и с коллагеном II. Основное внимание в этом исследовании уделялось димеру интегрина β1, поскольку он является наиболее заметным β-димером среди гетеродимеров хряща и, как известно, взаимодействует со многими различными α-димерами [12]. Кластер дифференцировки 29 (CD29) представляет собой субъединицу интегрина β1, ассоциированную с очень поздними антигенными рецепторами, экспрессирующуюся почти на всех клетках и типах тканей.
Здесь использовалась атомно-силовая микроскопия (АСМ), чтобы помочь нам измерить изменения во время хондрогенной дифференцировки hADSc. Как тип сканирующей зондовой микроскопии с очень высоким разрешением, АСМ предоставила новую возможность обнаруживать морфологию и клеточную мембрану отдельных клеток в жидкости на наноуровне. Между тем, для измерения связывания лиганд-рецептор на живых клетках использовалась система силовой спектроскопии одиночных молекул (SMFS) в сочетании с атомно-силовой микроскопией (AFM). Система SMFS была более чувствительна к изменениям рецепторов в клеточной мембране, и изображения силы связывания были визуализированы. В этой работе связывание интегрина β1-лиганд-рецептор исследовали с помощью АСМ-наконечников, функционализированных интегрином β1. При применении АСМ было обнаружено, что хондрогенная дифференцировка изменяет форму клеток hADSc и увеличивает шероховатость клеток. Это приложение предоставило метод для оценки хондрогенной дифференцировки путем прямого измерения взаимодействий интегрина β1-лиганд-рецептор и изменения ультраструктуры клеточной поверхности, улучшая исследование и скрининг клеточной поверхности визуализированным способом. Хондрогенная дифференцировка изменяет состав и структуру мембран, а также внутриклеточные цитоскелетные взаимодействия. Эти изменения клеточной морфологии, ультраструктуры и связывания лиганд-трансмембранный рецептор служат полезными маркерами для оценки механизмов хондрогенной дифференцировки.
Методы
Культура клеток и реагенты
Для этого исследования клетки были выделены у трех хирургических пациентов (средний возраст 20 лет), как описано ранее [13]. Информированное согласие было получено от всех пациентов. Утверждение этики для этого исследования было получено в Первой аффилированной больнице Цзинаньского университета (форма приложения). Клетки поддерживали в базовой среде, которая включала среду Игла, модифицированную Дульбекко с низким содержанием глюкозы (DMEM, Life Technologies, Калифорния, США), с добавлением 10% инактивированной нагреванием фетальной телячьей сыворотки (FBS, Life Technologies, Калифорния, США), 100 единиц / мл пенициллина (Life Technologies, Калифорния, США), 100 мкг / мл стрептомицина (Life Technologies, Калифорния, США), 0,11 мг / мл пирувата натрия (Life Technologies, Калифорния, США) и L-глутамина (Life Technologies, CA, США). Клетки поддерживали при 37 ° C в увлажненном инкубаторе, содержащем 5% CO 2 . со средой меняют каждые 3 дня.
Дифференциация in vitro
Для хондрогенной индукции hADSc с четвертого по восьмой пассажи высевали с высокой плотностью клеток (2 × 105/10 мл) и культивировали в хондрогенной среде, содержащей DMEM / F12 с добавлением 1% FBS, 1% инсулина-трансферина-селена ( ITS) + добавка (Cyagen, Гуанчжоу, Китай), 10 нг / мл трансформирующего фактора роста-бета1 (TGF-β1) (Peprotech, Rocky Hill, Нью-Джерси, США), 100 нг / мл инсулиноподобных факторов роста-1 ( IGF-1) (Peprotech, Peprotech, Rocky Hill, Нью-Джерси, США), 10-7 M дексаметазон (Sigma, Сент-Луис, Миссури, США) и 50 мкг / мл аскорбиновой кислоты (Sigma, Сент-Луис, Миссури). , США). Среду меняли каждые 2 дня, добавляя только что добавленные TGF-β1 и IGF-1. Хондрогенез оценивали по окрашиванию альциановым синим и толуидиновым синим.
Чтобы вызвать остеогенную и адипогенную дифференцировку, клетки четвертого-восьмого пассажа обрабатывали остеогенной и адипогенной средой в течение 2 недель соответственно. Остеогенная среда состояла из DMEM с добавлением 10-7 М дексаметазона (Sigma, Сент-Луис, Миссури, США), 50 мкг / мл аскорбиновой кислоты (Sigma, Сент-Луис, Миссури, США) и 10 ммоль / л β-глицерина. фосфат (Sigma, Сент-Луис, Миссури, США). Остеогенез оценивали по окрашиванию ализарином красным.
Адипогенная среда состояла из DMEM с добавлением 0,5 ммоль / л 3-изобутил-1-метилксантина (IBMX) (Sigma, Сент-Луис, Миссури, США), 1 мкмоль / л гидрокортизона (Sigma, Сент-Луис, Миссури, США), Индометацин 0,1 ммоль / л (Sigma, Сент-Луис, Миссури, США). Адипогенную дифференциацию оценивали по окрашиванию Oil Red O.
Идентификация поверхностных антигенов hADSc с помощью проточной цитометрии
HADSC были переварены трипсином, а затем дважды промыты DMEM перед повторным суспендированием при плотности клеток 2 × 10 7 клеток / мл. Клеточная суспензия (50 мкл; 1 × 10 6 клеток) был добавлен в пробирки с эпоксидным эпоксидом объемом 1,5 мл и затем инкубирован с антителами против CD34, анти-CD44, анти-CD45, анти-CD73, анти-CD90, анти-CD106, анти-HLA-DR и анти-CD105. 20 мин при 37 ° C в темноте. Анти-CD34, анти-CD44 и анти-CD45 были получены от CST (Беверли, Массачусетс, США); другие антитела были получены от Abcam (Кембридж, Массачусетс, США). Затем суспензию клеток центрифугировали при × 500 g в течение 5 мин с последующим удалением супернатанта и ресуспендированием клеток в 200 мкл буфера для окрашивания. Все шаги были повторены дважды перед анализом методом проточной цитометрии.
Анализ иммуноблоттинга (IB)
Клетки собирали для иммуноблоттинга, как описано ранее [14]. В качестве первичных используемых антител были антитела против β-катенина (ab32572), против интегрина β1 (ab30394) и против коллагена II (ab34712), полученные от Abcam (Кембридж, Массачусетс, США). Анти-β-актин (8H10D10, 1:2000), анти-GSK-3β (27C10, 1:1000) и анти-SOX (92G2, 1:1000) были получены от Cell Signaling Technology (CST, Beverly, MA, США). Вторичные антитела, конъюгированные с HRP (1:1000–1:3000), были приобретены у CST.
Иммунофлуоресценция
Для хондрогенной дифференцировки клетки обрабатывали в течение 0, 6 и 12 дней, переваривали и культивировали на стекле в 24-луночных планшетах (Costar353047, Corning, Нью-Йорк, США) в течение 24 часов. Клетки дважды промывали ледяным фосфатным буферным раствором (PBS), фиксировали 4% параформальдегидом в течение 15 мин при комнатной температуре. После блокирования клетки инкубировали с первичным антителом, реактивным с интегрином β1, в течение 1 часа с последующей инкубацией в течение 1 часа в темноте с меченным Alexa Fluor 488 антимышиным IgG (H + L) (CST # 4408, MA, USA. ), 4 ', 6-диамидино-2-фенилиндол (DAPI, Sigma, Миссури, США). Для окрашивания фаллоидином после блокирования клетки пермеабилизировали 0,2% Triton X-100 в течение 30 мин, затем клетки инкубировали с DAPI и фаллоидином-Alexa Flour 573 (Life Technologies, Калифорния, США) в течение 1 часа. После трехкратной промывки субклеточную локализацию интегрина β1 и изменение нитчатого актина (F-актина) оценивали во время дифференцировки хряща с помощью лазерного сканирующего конфокального микроскопа (ZEISS, LSM 700, Оберкохен, Германия).
Подготовка советов AFM
Наконечники Si3N4 (DNP-10, Bruker Corp) с константой пружины (0,06 Н / м) были химически модифицированы антителом к CD29 следующим образом [15]. Наконечники очищали ацетоном, ультрафиолетом и раствором пираньи (H 2 SO 4 :H 2 О 2 =3:1, v / v ) в разное время (5 мин, 30 мин и 10 мин). После тщательной промывки очищенной водой кончики формировали путем инкубации с раствором 1% 3-APTES (Sigma, Сент-Луис, Миссури, США) в этаноле в течение 30 мин. Наконечники трижды промывали ультрачистой водой и обрабатывали 2,5% раствором глутаральдегида (Sigma, Сент-Луис, Миссури, США) в течение 1 часа. Лишний глутаральдегид трижды промывали водой. Наконец, наконечники помещали в раствор анти-интегрина β1 (1 мг / мл) и инкубировали в течение ночи при 4 ° C. Перед экспериментами модифицированные зонды промывали PBS.
Измерения AFM
AFM (Bioscope Catalyst, Bruker, США) использовали для исследования морфологии hADSc и ультраструктурных изменений во время хондрогенной дифференцировки. Точная постоянная силы наконечников AFM измерялась в PBS. Для оценки морфологии и ультраструктуры клетки несколько раз промывали PBS. Затем добавили 4% раствор параформальдегида в 3,5-см 2 блюдо для культивирования на 15 мин. После промывания клеток PBS клетки хранили в PBS при 4 ° C до использования. Жесткость пружины наконечников составляла от 4,2 до 5,8 Н / м в контактном режиме. Морфологические и ультраструктурные изображения hADSc были получены в PBS при комнатной температуре с помощью АСМ. Изображения ультраструктуры, окружающие ядра hADSc, были получены в контактном режиме. Программное обеспечение для анализа наноскопа использовалось для оценки ультраструктуры клеточной поверхности для более чем 15 различных изображений 10 × 10 мкм, по крайней мере, для 15 различных клеток в группах (день 0, 6, 12). Силу связывания между интегрином β1-модифицированными наконечниками AFM и рецепторами CD29 живых hADSc анализировали в течение различных хондрогенных периодов (0, 6 и 12 дней). Сила связывания измерялась в режиме приближения-втягивания системы AFM (Bioscope Catalyst, Bruker, США). Для изучения событий разделения интегрина β1 и живых клеток использовали модифицированные антителом интегрина β1 наконечники при скоростях приближения-отвода 500 нм / с. Силовая постоянная функционализированных наконечников составляла 0,058 ± 0,006 Н / м. Пороговое усилие на клетки составляло 800 пН. Антитело против интегрина β1 (100 мкг / мл) добавляли к клеткам на 30 мин перед экспериментами по измерению силы. Блокирующие интегрин β1 и незащищенные зонды также использовали в качестве контроля для обнаружения неспецифической силы разрыва между кончиками, модифицированными интегрином β1 антителом, и клетками. Для количественной оценки вероятности связывания лиганда интегрина β1 с рецептором кривые силы специфического взаимодействия измеряли с помощью зондов, функционализированных антителом интегрина β1. В одном эксперименте было измерено более 400 силовых кривых, а результаты суммированы по крайней мере из трех независимых экспериментов. Таким образом, приблизительно 1200 исходных кривых сила-расстояние в каждом сравнительном эксперименте были получены из 30-40 различных ячеек с использованием программного обеспечения для анализа Nanoscope. Путем усреднения значений силы по крайней мере для трех независимых экспериментов было определено влияние хондрогенной индукции на силу взаимодействия между лигандом интегрина β1 и рецепторами CD29 на поверхности клетки.
Обратная транскрипция и ПЦР в реальном времени
Использовали наборы для очистки РНК TRIzol® Plus (Life Technologies, Калифорния, США), и 1 мкг РНК подвергали обратной транскрипции в кДНК с использованием набора для обратной транскрипции кДНК высокой емкости (Invitrogen) в соответствии с протоколом производителя с небольшими модификациями. Интегрин β1 и GAPDH были количественно определены с помощью qRT-PCR со специфическими для генов праймерами:5'-TGGAGGAAATGGTGTTTGC-3 '(интегрин β1-смысл) и 5'-CGTTGCTGGCTTCACAAGTA-3' (интегрин β1-антисмысловой); 5'-CTGACTTCAACAGCGACACC-3 '(GAPDH-смысл) и 5'-CCCTGTTGCTGTAGCCAAAT-3' (GAPDH-антисмысловой). Для ПЦР в реальном времени выполняли пошаговую ПЦР в реальном времени (Applied Biosystems) с использованием Fast SYBR @ GREEN Master Mix (Life Technologies, Калифорния, США). Экспрессию целевого гена нормализовали к GAPDH в качестве внутреннего стандарта и рассчитывали с использованием сравнительного метода 2-ΔΔCT. Каждый анализ проводился в трех экземплярах.
Статистический анализ
Все эксперименты проводили не менее трех раз, данные выражали как среднее ± стандартное отклонение (SD). Сравнение двух групп было проведено t контрольная работа. Существенные различия между средними значениями групп определяли с помощью однофакторного анализа ANOVA, за которым следовали тесты Бонферрони и Тамхана T2 (одинаковые дисперсии не предполагались). Значения p <0,05 считались статистически значимыми.
Результаты и обсуждение
Оценка hADSc
Мезенхимальные стволовые клетки - это мультипотентные стромальные клетки, которые обладают остеогенным, адипогенным, хондрогенным и миогенным потенциалом. Существует два основных способа идентификации hADSc:маркеры CD клеточной поверхности и способность к дифференцировке [16]. Как показано в Дополнительном файле 1:Рисунок S1 и Дополнительном файле 2:Рисунок S2, производные клетки были hADSc. Затем с помощью МТТ-анализа определяли пролиферацию клеток hADSc пассажа 3 (дополнительный файл 3:Рисунок S3).
Индуцированные изменения морфологии и ультраструктуры поверхности во время хондрогенеза hADSc
АСМ всегда используется для определения морфологии и ультраструктуры клеток на наномасштабе [17]. Форма клетки связана с ее специализированной функцией клетки и организацией ткани. В некоторых исследованиях рака АСМ можно использовать в качестве метода визуализации для анализа морфологических изменений с целью оценки эффектов лекарств. Кроме того, форма мезенхимальных стволовых клеток изменяется во время хондрогенной индукции [18]. Хотя изменения формы клеток, по-видимому, необходимы для дифференцировки, мало что известно о том, влияет ли морфология клеток на ранние стадии развития дифференцировки мезенхимальных стволовых клеток. Следовательно, морфология и изменения ультраструктуры мембраны во время хондрогенеза hADSc были оценены с помощью АСМ, поскольку эти изменения важны [19] и могут напрямую влиять на функцию клеток [20]. Морфология поверхности и ультратонкая структура hADSc были исследованы во время хондрогенной дифференцировки в разные периоды времени (рис. 1 и 2). Очевидно, что морфология и ультраструктура поверхности различались в каждой группе сравнения. На 0 сутки клетки имели форму удлиненного веретена с относительно гладкой поверхностью. Архитектура клеточной мембраны была однородной. После хондрогенной индукции на 6 и 12 дни наблюдали значительные изменения морфологии клеток. Большинство клеток постепенно сжимались до многоугольной формы (рис. 1a) с уменьшением среднего отношения длины / ширины клеток во время хондрогенной дифференцировки (рис. 1b). Многочисленные исследования показывают, что изменения морфологии клеток соответствуют цитоскелету клеток [21]. Мы также обнаружили изменения цитоскелета во время хондрогенной дифференцировки, что было объяснено в последних результатах.
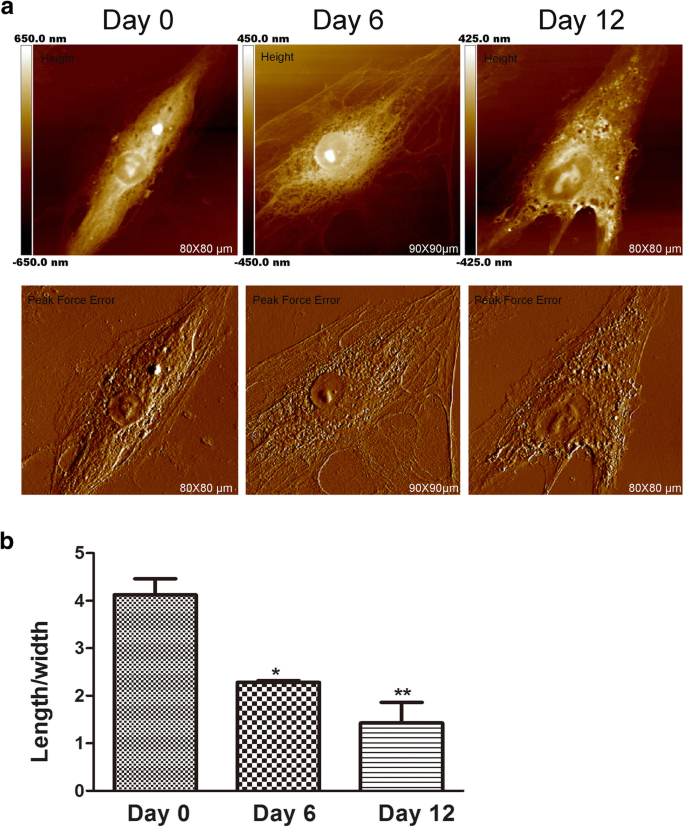
Особенности морфологии hADSc при хондрогенезе. а Морфологические изображения всего hADSc были получены на 0, 6 и 12 день хондрогенной дифференцировки. Изображения были проанализированы с помощью модели изображения ошибки высоты и пиковой силы с помощью наноскопа. б Среднее отношение длины / ширины клеток измеряли после обработки хондрогенной дифференцировкой через 0, 6 и 12 дней. * p <0,05, ** p <0,01
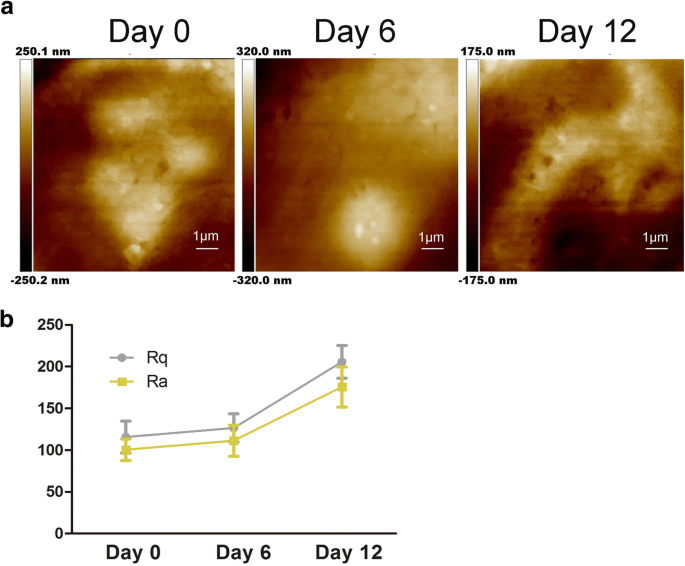
Характеристики ультраструктуры мембраны hADSc при хондрогенной дифференцировке. а Изменения ультраструктуры клеточной мембраны оценивали после хондрогенной дифференцировки в течение 0, 6 и 12 дней. б Параметры шероховатости поверхности Ra и Rq клеток измеряли во время хондрогенной индукции hADSc в течение 0, 6 и 12 дней
Как показано на рис. 2а, ультраструктура клеточной мембраны также изменилась; частицы стали увеличенными и неоднородными. Предыдущие исследования показали, что Ra и Rq были определяющими величинами шероховатости для оценки изменений в клеточных мембранах, обработанных по-разному [22]. Rq - это среднеквадратичная шероховатость, \ (\ mathrm {Rq} =\ sqrt {\ frac {\ sum_ {t-1} ^ N {\ left (Zn- \ overline {Z} \ right)} ^ 2 } {N-1}} \); \ (\ mathrm {Rq} =\ sqrt {\ frac {\ sum _ {\ mathrm {t} -1} ^ {\ mathrm {N}} {\ left (\ mathrm {Zn} - \ overline {\ mathrm {Z) }} \ right)} ^ 2} {\ mathrm {N} -1}}; \) Ra означает среднюю шероховатость, \ (\ mathrm {Ra} =\ frac {1} {N} {\ sum} _ { t-1} ^ N1 \ mid Zi- \ overline {Z} \ mid \). Чтобы получить шероховатость, размер сканирования составляет 10 мкм × 10 мкм. Как показано на рис. 2b, как Ra, так и Rq двух разных областей увеличиваются во время хондрогенеза hADSc. Значения Ra и Rq клеток в день 0 были низкими, что указывает на гладкую поверхность (рис. 2b). Значения Ra и Rq одновременно увеличивались с хондрогенной дифференцировкой, показывая большую гетерогенность и шероховатость на поверхности клеток (рис. 2а). На основании наблюдаемых изменений хондрогенная дифференцировка привела к изменению морфологии клеток и соотношению высоты / ширины клеток (рис. 1a, b). Есть исследования, показывающие, что ECM может регулировать клеточную адгезию путем регулирования интегринов [11]. Следовательно, увеличенные значения шероховатости предполагают изменения ECM и ультраструктуры клеточной мембраны во время хондрогенеза. Эти данные демонстрируют, что хондрогенная дифференцировка влияет на морфологию клеток, ECM и структуру клеточной мембраны.
Изменения цитоскелета во время хондрогенной индукции hADSc
Во время дифференцировки стволовых клеток морфология клетки и структурные изменения мембраны связаны с цитоскелетом клетки после развития клон-специфичных клеточных характеристик [21]. Как показано на фиг. 3a, красный и синий сигналы флуоресценции соответственно указывают на F-актин и DAPI. Цитоскелет клетки сильно изменился во время хондрогенной индукции (рис. 3а). С одной стороны, микрофиламенты цитоскелета шли вдоль длинной оси клетки в день 0 группы, в то время как микрофиламенты цитоскелета распространялись радиальным массивом, когда hADSc обрабатывали хондрогенной дифференцировкой в течение 12 дней. С другой стороны, распределение клеточных микрофиламентов было однородным в группе 0 дня, но микрофиламенты в основном были распределены по периферии hADSc, обработанной хондрогенной дифференцировкой в течение 12 дней.
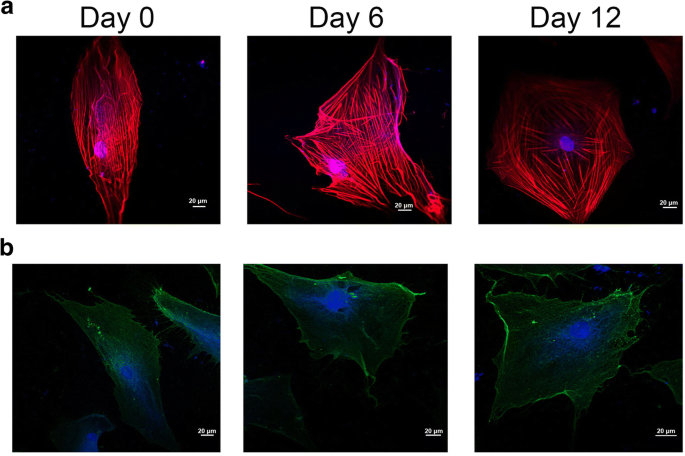
Организация цитоскелета и расположение интегрина β1 на хондрогенно дифференцирующихся hADSc. а Изменения цитоскелета выявляли при хондрогенезе hADSc с помощью конфокальной микроскопии. б Местоположение интегрина β1 измеряли во время хондрогенной дифференцировки с помощью конфокальной микроскопии. Цитоскелет и ядро окрашивали F-актином и DAPI соответственно. Красный и синий сигналы флуоресценции соответственно указывают на F-актин и DAPI
Хондрогенная дифференцировка изменила вероятность связывания интегрина β1 с рецепторами на hADSc
АСМ также является полезным инструментом для изучения силы связывания между лигандами и их рецепторами, делая четкую передачу сигнала мембрана-рецептор на поверхности клеток [23]. С помощью AFM изменения между интегрином β1 и его рецепторами измеряются визуально, простым и специфическим способом. Взаимодействие лиганда-рецептора интегрина β1 с живыми клетками - это способ исследовать процесс связывания на клеточной мембране. Процедура функционализации наконечника AFM заключается в связывании интегрина β1 с наконечниками AFM посредством связывания APTES и глутаральдегида. Эти наконечники использовали для обнаружения связывания интегрина β1 с рецепторами CD29 на поверхности клеток (рис. 4а). Одномолекулярная силовая спектроскопия (SMFS) была использована для оценки распределения силы разделения живых клеток против интегрина β1 в локализованных областях отдельных живых hADSc (рис. 4b). Типичные силовые кривые показаны на фиг. 4c, d, которые изображают кривую для одной молекулы (фиг. 4c) и две пары кривых пиков разрыва (фиг. 4d). Для проверки специфичности полученных силовых кривых были проведены эксперименты по блокировке и эксперименты с голыми наконечниками АСМ. Голые наконечники АСМ не обнаружили пика удельной силы (рис. 4e). Эксперименты с открытой АСМ показали, что вероятность неспецифического связывания взаимодействия лиганда интегрина β1 с рецептором на поверхности hADSc составляет менее 1%. Для экспериментов по блокированию антитело против интегрина β1 инкубировали с клетками в течение 30 минут, а затем записывали силовые кривые с использованием наконечников, функционализированных интегрином β1. Блокирующее антитело снижает силовые кривые на 90% (рис. 4f). Не было различий в вероятности связывания лиганда-рецептора интегрина β1 на поверхности клеток среди трех групп после обработки антителом против интегрина β1 (рис. 4g). Эти результаты демонстрируют, что наконечники AFM, модифицированные антителами, были очень полезны для определения силы, и что наконечники AFM, функционализированные интегрином β1, были специфичными.
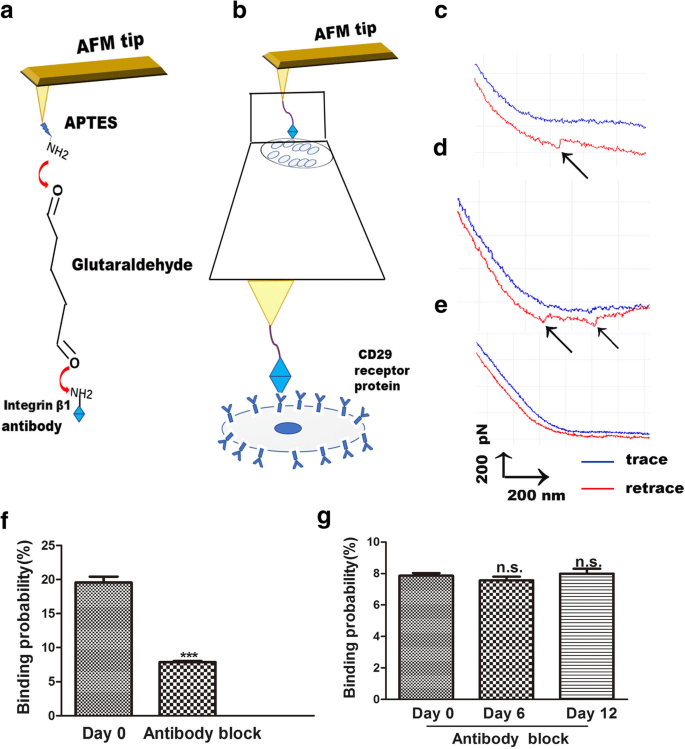
Измерения силы АСМ с помощью наконечника АСМ, функционализированного интегрином β1, на живых hADSc. а Схематическое изображение стратегии, используемой для иммобилизации интегрина β1 на наконечнике AFM. б Схематическое изображение силы одной молекулы, измеренной между кончиками АСМ, функционализированными интегрином β1, и живыми hADSc. c, d Типичные силовые кривые, полученные с помощью наконечников АСМ, модифицированных интегрином β1, на hADSc и e после блокирования системы раствором моноклонального антитела к интегрину β1. е Вероятность связывания кончиков, функционализированных интегрином β1, на hADSc до и после блокирования антителом к интегрину β1 в день 0. g Вероятность связывания CD29-функционализированных наконечников на hADSc после блокирования антителом к интегрину β1 через 0, 6 и 12 дней. *** p <0,001, н.с .. нет существенной разницы
Сила связывания (сила разрыва) - это сила взаимодействия между лигандами и их рецепторами [24]. Изменения морфологии и ультраструктуры поверхности плазматических мембран связаны со многими процессами клеточной биологии, такими как дифференцировка, апоптоз и миграция клеток. Считается, что во время дифференцировки изменения цитоскелета связаны с изменениями интегрина, особенно интегрина β1. Интегрин β1 (CD29) очень важен для клеточной адгезии к ECM и межклеточной адгезии. Он также может взаимодействовать с внутриклеточными белками, стимулируя сигнальные молекулы, связанные с актиновым цитоскелетом [25]. В этом исследовании изменения цитоскелета и морфологии клеток наблюдались во время хондрогенеза hADSc с помощью конфокальной лазерной сканирующей микроскопии (CLSM) и AFM. Во время хондрогенной дифференцировки изменения цитоскелета, морфологии и ультраструктуры поверхности могут быть новым и надежным индикатором состояния клеток. Интегрин β1, рецептор CD29, распределяется по поверхности клетки, судя по иммунофлуоресценции (рис. 3b). Сила связывания и стабильность комплексов лиганд интегрина β1-рецептор во время хондрогенеза hADSc оценивали на 0, 6 и 12 день дифференцировки. Всего за каждый день было записано 1200 кривых со средними усилиями разрыва 61,8 ± 22,2 пН, 60 ± 20,2 пН и 67,2 ± 22,0 пН, соответственно (рис. 5a – c). Распределение величины силы было проанализировано как среднее значение силы + SD (рис. 5г). Значительной разницы в средней силе между 0 и 6 днями не было. Разница в средней силе между 0 и 12 днями. Величина силы связывания увеличивалась на 12 день. Между тем, события разрыва в 0, 6 и 12 дни. составляли соответственно 19,58 ± 1,74%, 28,03 ± 2,05% и 33,4 ± 1,89% (рис. 5д). Повышенная вероятность связывания также указывает на то, что интегрин β1 (CD29) играет важную роль в хондрогенной дифференцировке и может предоставлять информацию для хондрогенной дифференцировки через сигнальные пути. Следовательно, увеличение нанодоменов β1 интегрина во время хондрогенной дифференцировки может фундаментально влиять на силу связывания лиганд-рецептор CD29 на живых hADSc. Изменения морфологии и ультраструктуры поверхности плазматических мембран сопровождают изменения структуры белка интегрина β1, конформации, силы связывания и стабильности комплексов лиганд интегрина β1 – рецептор на клетках. Таким образом, интегрин β1 играет необходимую роль в хондрогенной дифференцировке hADSc.
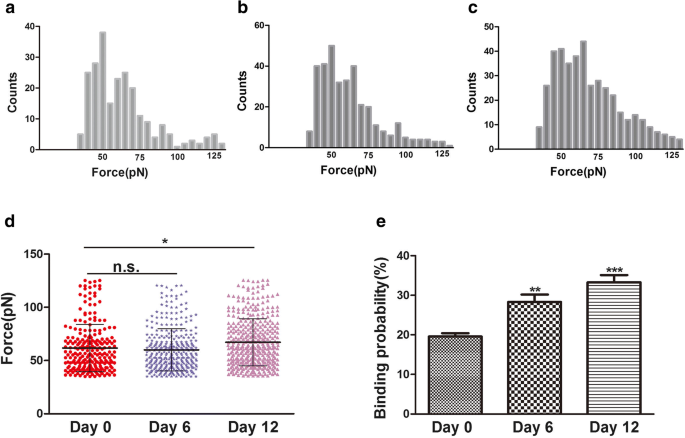
Сила связывания и вероятность связывания измеряются на поверхности живых hADSc с помощью наконечников АСМ, функционализированных интегрином β1. а – в Гистограммы силы связывания интегрина β1 антитело-рецептор, полученные в ходе хондрогенной дифференцировки hADSc в течение 0, 6 и 12 дней. г Силы связывания для интегриновых β1-рецепторов были получены на 0, 6 и 12 день хондрогенной дифференцировки hADSc. е Вероятность связывания интегринового β1-рецептора определяли при хондрогенной дифференцировке hADSc в течение 0, 6 и 12 дней. * p <0,05, ** p <0,01, *** p <0,001, н.у. нет существенной разницы
Повышающая регуляция интегрина β1 во время хондрогенной дифференцировки hADSc
Многочисленные исследования показали, что члены семейства интегринов играют важную роль в дифференцировке клеток. Кроме того, интегрины могут регулировать взаимодействие между внеклеточной средой и клетками, контролируя пути передачи сигналов через связанные белки [26]. Предыдущие исследования показали, что вероятность связывания может зависеть от плотности и конформации трансмембранного белка (рецепторов) на поверхности клетки [27]. Конформация интегрина может представлять собой закрытый головной убор, имеющий низкое сродство к лиганду, или открытый головной убор, который имеет высокое сродство к лиганду [28, 29]. Экспрессия интегрина β1 повышалась как на уровне транскрипции, так и на уровне трансляции с повышенной экспрессией коллагена II, характерной для хондроцитов (рис. 6a, b). Таким образом, повышенная экспрессия интегрина β1 согласовывалась с повышенной вероятностью связывания безотносительно к конформации.

The role of integrin β1 and β-catenin/SOX pathway in regulating hADSc chondrogenic differentiation. а Protein integrin β1 was up-regulated during chondrogenesis of hADSc as assessed by western blotting. Cartilage differentiation up-regulated collagen II expression at different days. б The mRNA of integrin β1 was up-regulated during chondrogenic differentiation of hADSc. c Measurement of proteins associated with the β-catenin/SOX pathway during chondrogenic differentiation of hADSc for 0, 6, and 12 days. * p <0,05, ** p < 0.01
The Role of Integrin β1 in Chondrogenic Differentiation Regulated by the β-catenin/SOX Signaling Pathway
Previous studies have shown Wnt/β-catenin, PI3K, and mTOR signaling pathways to be related to integrin β1 [30,31,32]. Each is important in mesenchymal stem cell differentiation. Likewise, studies have demonstrated SOX and collagen II to be regulated by integrin β1 during chondrogenesis of hADSc. SOX is a hallmark component of the Wnt/β-catenin signaling pathway. Hence, we hypothesized that chondrogenic differentiation was regulated by the β-catenin/SOX pathway via integrin β1. SOX, GSK-3β, β-catenin, and integrin β1 were all increased during chondrogenesis of hADSc (Fig. 6c), with integrin β1 inducing cell signaling. These data demonstrate chondrogenic differentiation to be regulated by the β-catenin/SOX pathway via integrin β1.
Prospective and Limitations
In this work, changes in cellular morphology, the structure of the membrane, and the binding probability of integrin β1 ligand–receptors were demonstrated to be useful image markers to evaluate the chondrogenic differentiation process. This is a new method for evaluation of morphology, membrane ultrastructure, and changes in transmembrane proteins during chondrogenic differentiation. There are limitations to this study. Although increased binding probability was related to the high expression of integrin β1, the conformation of integrin β1 during chondrogenesis was not investigated. Further work is necessary to determine the conformation of integrin β1 during chondrogenic differentiation. Integrin β1 was demonstrated to participate in the β-catenin/SOX signaling pathway during chondrogenesis of hADSc. However, the relationship between integrin β1 and β-catenin/SOX signaling pathway is still not fully established. Further work is necessary to identify the exact role of integrin β1 in this pathway.
Выводы
In the present work, a novel method (AFM) was employed to evaluate chondrogenic induction in hADSc. Cell surface ultrastructural changes were assessed by AFM imaging. AFM was used to investigate the binding force and binding probability between integrin β1 ligand and its receptors on the surface of hADSc by integrin β1-functionalized AFM tips. Based on AFM data, during chondrogenesis, cell morphology was changed from an elongated spindle shape to a polygonal shape with increased cell roughness. By use of integrin β1-functionalized AFM tips, the binding probability and force magnitude of integrin β1 ligand–receptor on the surface of hADSc were found to increase during chondrogenic induction. By immunoblot, integrin β1 was demonstrated to participate in the β-catenin/SOX signaling pathway, which regulated the chondrogenesis of hADSc. Taken together, these results and the established methodology contribute to a better understanding of cell morphology and roughness. Further, the data provide thermodynamic and kinetic insight into the integrin β1 ligand-binding process, at the single-molecule level. This AFM method will be useful for investigation of signaling pathways in living hADSc during chondrogenesis. Changes in the cellular nanostructure, as well as structure of the membrane, and the binding probability of transmembrane proteins are useful markers to evaluate chondrogenic differentiation mechanisms. This AFM method can be used to understand the mechanism of mesenchymal stem cell differentiation in tissue engineering and will be useful for an enhanced understanding of mesenchymal stem cell chondrogenic differentiation.
Сокращения
- AFM:
-
Атомно-силовая микроскопия
- CD:
-
Cluster of differentiation
- DAPI:
-
4′,6-diamidino-2-phenylindole
- DMEM:
-
Dulbecco’s Modified Eagle’s Medium
- ECM:
-
Extracellular matrix
- F-actin:
-
Нитчатый актин
- FBS:
-
Фетальная бычья сыворотка
- hADSc:
-
Human adipose-deprived stem cells
- IB:
-
Immunoblotting analysis
- IBMX:
-
3-isobutyl-1-methylxanthine
- IGF-1:
-
Insulin-like growth factors-1
- ITS:
-
Insulin transferrin selenium
- mTOR:
-
Mammalian target of rapamycin
- OA:
-
Osteoarthritis
- PBS:
-
Фосфатный буферный раствор
- PI3K:
-
Phosphoinositide 3-kinase
- SD:
-
Standard deviation
- SMFS:
-
Single-molecule force spectroscopy
- TGF-β1:
-
Transforming growth factor-beta1
Наноматериалы
- Демонстрация гибкого биосенсора на основе графена для чувствительного и быстрого обнаружения клеток рака яи…
- Усовершенствованные технологии осаждения атомного слоя для микро-светодиодов и VCSEL
- Наночастицы черного фосфора способствуют остеогенной дифференцировке EMSC посредством усиленной экспрессии …
- Раскрытие атомной и электронной структуры углеродных нановолокон с набором чашек
- Анализ организации актина и фокальной адгезии в клетках U2OS на полимерных наноструктурах
- Нанотехнологии инактивируют раковые стволовые клетки
- Сравнение металлических электродов с рисунком типа Nanohole и Nanopillar, используемых в органических солнечных элем…
- Синтез нанокристаллов ZnO и применение в инвертированных полимерных солнечных элементах
- Изучение силы адгезии и стеклования тонких пленок полистирола с помощью атомно-силовой микроскопии
- Возобновляемый флуоресцентный зонд BHN-Fe3O4 @ SiO2 Гибридная наноструктура для Fe3 + и ее применение в биоимиджинге