


Экзосомная микроРНК-18b-3p, полученная из мезенхимальных стволовых клеток пуповины человека, подавляет возникновение преэклампсии, воздействуя на LEP
Аннотация
Экзосомы, полученные из мезенхимальных стволовых клеток пуповины человека (hucMSC), экспрессирующие микроРНК, были выявлены при заболеваниях человека. Однако подробный молекулярный механизм происходящих из hucMSC экзосомальных miR-18b-3p на преэклампсию (PE) остается дальнейшим исследованием. Мы стремились изучить влияние экзосом и miR-18b-3p / лептина (LEP) на возникновение ПЭ. Выявлена морфология экзосом hucMSC и hucMSC (Exos). Экзосомы инфицировали различными лентивирусами, экспрессирующими miR-18b-3p, чтобы исследовать роль miR-18b-3p в PE. Модель PE на крысах была создана путем внутрибрюшинной инъекции N -нитро-1-аргинин метиловый эфир. Экспрессию LEP и miR-18b-3p тестировали в тканях плаценты PE крысы. Также было обнаружено влияние экзосом на экспрессию LEP и miR-18b-3p. Определяли систолическое артериальное давление (САД), протеинурию, воспалительные факторы, массу плода крысы и плаценты, а также апоптоз клеток у крыс PE. Наконец, взаимосвязь между miR-18b-3p и LEP была подтверждена с использованием анализа двойного люциферазного репортерного гена и анализа РНК с понижением. Экзосомы, восстанавливающие miR-18b-3p или ингибирующие LEP, снижали САД и протеинурию у крыс с PE, а также увеличивали вес плода крысы и плаценты, снижали уровни воспалительных факторов в сыворотке крови, а также подавляли апоптотические клетки крыс PE, оказывая подавляющее действие по прогрессированию ПЭ. miR-18b-3p был снижен, а LEP увеличен в тканях плаценты PE крыс. LEP был прямым геном-мишенью miR-18b-3p. Повышающая регуляция miR-18b-3p или обработка экзосом подавляла экспрессию LEP и снижала возникновение PE, тогда как подавление miR-18b-3p приводило к противоположным эффектам. Подавление LEP обращало эффект восстановления miR-18b-3p на PE крыс. Произведенный из HucMSCs экзосомальный miR-18b-3p нацелен на участие LEP в возникновении и развитии PE. Это исследование может предоставить новую теоретическую основу для механизма и исследования ПЭ.
Введение
Преэклампсия (ПЭ), характеризующаяся протеинурией и гипертонией [1], является основной причиной материнской и материнской смертности и заболеваемости при беременности человека [2]. Этиология и патогенез ПЭ не ясны [3], которая, как сообщается, связана с аномальной инвазией трофобластов, приводящей к дисфункции эндотелия у матери, хронической недостаточности перфузии плаценты и гипертонии с неблагоприятными исходами [4]. За исключением родов плода и плаценты, специфической терапии ПЭ не существует [5]. Поэтому необходимо срочно изучить терапевтические цели, чтобы улучшить прогноз этого заболевания.
Пуповина человека (huc) является подходящим источником мезенхимальных стволовых клеток (МСК), которые секретируют множество трофических факторов и цитокинов, а также обладают сильными противовоспалительными и иммуномодулирующими свойствами [6]. Исследование подтвердило, что PE ускоряет экспрессию нейроглиальных маркеров в MSCs Wharton пуповинного желе [7]. Сообщалось также о защитном эффекте экзосом hucMSCs (Exos) на морфологию плаценты и ангиогенез у PE крыс [8]. Экзосомы представляют собой небольшие (50–100 нм) секреторные везикулы, которые обеспечивают связь между клетками в микроокружении опухоли посредством инкапсуляции и передачи канцерогенных факторов в отдаленные участки или окружающие клетки посредством циркуляции [9]. Исследование выявило повреждение функций сосудов и осложнения, вызванные эффективным переносом sFlt-1 и sEng в эндотелиальные клетки у пациентов с ПЭ с помощью экзосом [10]. МикроРНК (миРНК) представляют собой эндогенные некодирующие РНК длиной 18–25 нуклеотидов и регулируют экспрессию гена на посттранскрипционном уровне [11]. Данные исследования показали, что экспрессия miR-18b влияет на клеточную инвазию, жизнеспособность и миграцию клеток трофобласта при PE [12]. Кроме того, Wu et al. предположили, что miR-18b ослабляет пролиферацию эндотелиальных клеток сетчатки человека, индуцированную высоким содержанием глюкозы, что может предложить новое понимание механизма патогенеза диабетической ретинопатии [13]. Однако роль происходящей из hucMSC экзосомальной miR-18b-3p в PE остается неизвестной. Лептин (LEP) оказывает плейотропное действие на дифференцировку / пролиферацию клеток и иммунитет к физиологическим состояниям и в основном выделяется из адипоцитов в дополнение к другим тканям, включая плаценту [14]. Исследование подтвердило, что аномальное метилирование промотора LEP участвует в прогрессии PE [15]. Другое исследование показало, что плацента является основным местом экспрессии LEP во время беременности [16]. Тем не менее, связь связывания между miR-18b-3p и LEP все еще неуловима. Таким образом, мы стремились изучить роль происходящей из hucMSC экзосомальной miR-18b-3p в ПЭ с участием LEP, и мы пришли к выводу, что происходящая из hucMSC экзосомная miR-18b-3p может ингибировать прогрессию PE посредством нацеливания на LEP.
Материалы и методы
Этическое одобрение
Исследование было одобрено институциональным наблюдательным советом Народной больницы Уханьского университета. Все участники подписали документ об информированном согласии. Все эксперименты на животных проводились в соответствии с Руководством по уходу и использованию лабораторных животных Международного комитета Народной больницы Уханьского университета.
Изолированность, культура и идентификация HucMSC
Пуповину плода, доставленную здоровым родильницей, собирали, нарезали на фарш и фильтровали через сито, затем смешивали с фосфатно-солевым буфером (PBS). Ткани пуповины центрифугировали при 1500 об / мин в течение 5 мин с радиусом центрифуги 10 см. Ткани суспендировали в среде Игла, модифицированной Дульбекко (DMEM) / F12, содержащей 10% фетальной телячьей сыворотки (FBS), и переносили в колбу для культивирования. Жидкости меняли через 4 дня, а затем меняли каждые 3 дня. Клетки субкультивировали, когда конфлюэнтность достигала около 90%. Адгезивный рост и морфологию hucMSCs наблюдали под световым микроскопом. Клетки окрашивали окрашивающим раствором масляным красным О (Beyotime Institute of Biotechnology, Шанхай, Китай) для выявления остеогенной дифференцировки hucMSC и окрашивали окрашивающим раствором щелочной фосфатазы (ALP) (Beyotime) для выявления адипогенной дифференцировки hucMSC. Проточный цитометр (Beckman Coulter Life Sciences, Бреа, Калифорния, США) был адаптирован для тестирования CD73, CD166 (оба 1:10, BD Biosciences, Франклин Лейкс, Нью-Джерси, США) и CD105 (1:20, AbD Serotec, Оксфорд, США). Великобритания).
Извлечение и идентификация HucMSC-Exos
Выращивали хорошо растущие hucMSC. Супернатант собирали и центрифугировали при 28 500 об / мин в течение 1 ч с радиусом центрифуги 10 см. Супернатант отбрасывали, клетки фиксировали 2% глутаровым альдегидом и 1% осмиевой кислоты, дегидратировали этанолом, погружали в оксид пропилена, сушили в течение 2 часов, заливали Epon812 и нарезали ломтиками. Срезы окрашивали ураном и свинцом соответственно. Наконец, экзосомы наблюдали под электронным микроскопом. Детектор Nanosight (Malvern Instruments, Малверн, Великобритания) использовался для обнаружения броуновского движения наночастиц экзосом и их размера. Поверхностные маркеры hucMSC-Exos были идентифицированы с помощью вестерн-блоттинга, и результаты показали, что hucMSC-Exos экспрессирует CD9, CD81 и CD63.
Метод заражения лентивирусом
HucMSC инфицировали лентивирусом, содержащим низкую экспрессию вектора miR-18b-3p и низкую экспрессию отрицательного контроля (NC) вектора miR-18b-3p (Shanghai GenePharma Co, Ltd, Шанхай, Китай). Наконец, были получены стабильно экспрессируемые NC hucMSC-антагомир и антагомир hucMSC-miR-18b-3p. Клетки культивировали в течение 48 часов, супернатант собирали и центрифугировали с ультрацентрифугированием, чтобы получить соответствующий экзос-антагомир NC и экзос-miR-18b-3p антагомир.
Экспериментальные животные
Были отобраны крысы Wistar (массой 200–250 г, возрастом 8 недель, независимо от пола) по здоровому уровню и половой зрелости (Центр экспериментальных животных Уханьского университета, Ухань, Китай). Крыс кормили в барьерной системе с температурой 18–28 ° C, относительной влажностью 40–70% и адекватным питанием и водой.
Создание моделей Rat PE
Модель ПЭ у крыс была создана путем внутрибрюшинной инъекции 50 мг / кг ингибитора синтетазы оксида азота, метилового эфира N (G) -нитро-1-аргинина (L-NAME, Beyotime) со ссылкой на статью [17]. Успешное создание модели ПЭ было основано на повышении артериального давления с 20 мм рт.ст. и выше 115 мм рт.ст., а также усилении протеинурии.
Группировка животных
Самка крысы и самец случайным образом проживали вместе в соотношении 1:1, и две крысы содержались в отдельной специальной клетке в 17–18 часов. предыдущий день. Сперму в вагинальном секрете самок крыс наблюдали с помощью вагинальной пробки и микроскопа на следующий день. Если результат был положительным в то же время, этот день записывали как 0-й день беременности. С 13-го rd В день беременности крысы были разделены на 6 групп (по 10 крыс в каждой группе):нормальная группа (такое же количество физиологического раствора вводили внутрибрюшинно с 13 по 20 день беременности), группа PE (L-NAME [50 мг / кг в день] вводили внутрибрюшинно с 13 по 20 день беременности, а 20 мкл физиологического раствора вводили в плаценту с 16 по 19 день беременности), группа PE + miR-NC (L-NAME [50 мг / кг в день] вводили внутрибрюшинно с 13 по 20 день беременности, и 20 мкл 4 нмоль miR-NC вводили в плаценту с 16 по 19 день беременности), PE + miR-18b-3p группа агомира (L-NAME [50 мг / кг в день] вводили внутрибрюшинно с 13 по 20 день беременности, а 20 мкл агомира miR-18b-3p 4 нмоль вводили в плаценту с 16 по 19 день беременности) , Группа антагомира PE + miR-18b-3p (L-NAME [50 мг / кг в день] вводили внутрибрюшинно с 13 по 20 день беременности, а 20 мкл 4 нмоль антагомира miR-18b-3p вводили в п Lacenta с 16 по 19 день беременности), PE + miR-18b-3p антагомир + небольшая интерферирующая РНК (si) -LEP группе (L-NAME [50 мг / кг в день] вводили внутрибрюшинно с 13 по 20 день. беременности, и 20 мкл 4 нмоль антагомира miR-18b-3p и si-LEP вводили в плаценту на 16-19 день беременности) и группе PE + si-LEP (L-NAME [50 мг / кг на день] вводили внутрибрюшинно с 13 по 20 день беременности, а 20 мкл 4 нмоль si-LEP вводили в плаценту с 16 по 19 день беременности). Крыс лечили экзосомами и экзосомами, несущими лентивирусы. Крысы были разделены на 5 групп (по 10 крыс в каждой группе):нормальная группа (такое же количество физиологического раствора вводили внутрибрюшинно с 13 по 20 день беременности), группа PE (L-NAME (50 мг / кг в день). ) вводили внутрибрюшинно с 13 по 20 день беременности, а 20 мкл физиологического раствора вводили в плаценту с 16 по 19 день беременности), группа PE + Exos (L-NAME (50 мг / кг в день) вводили внутрибрюшинно с 13 по 20 день беременности, и 20 мкл Exos (80 мкг экзосом суспендировали в 20 мкл физиологического раствора) вводили в плаценту с 16 по 19 день беременности), PE + Exos-antagomir NC группе (L-NAME (50 мг / кг в день) вводили внутрибрюшинно с 13 по 20 день беременности, и 20 мкл экзос-антагомира NC (80 мкг экзосом суспендировали в 20 мкл физиологического раствора) вводили в плаценту. с 16 по 19 день беременности) и группе антагомира PE + Exos-miR-18b-3p (L-NAME (50 мг / кг в день) вводили внутрибрюшинно с 13 дня на 20-й день гестации и 20 мкл антагомира Exos-miR-18b-3p (80 мкг экзосом суспендировали в 20 мкл физиологического раствора) вводили в плаценту на 16-19-й день беременности).
Определение систолического артериального давления (САД) и определение 24-часовой протеинурии
Давление у крыс измеряли путем измерения артериального давления в хвостовой артерии крыс. САД в хвостовой манжете всех беременных крыс измеряли на 10, 13, 16 и 19 день беременности с использованием детектора давления в хвостовой артерии крысы (Tensys® Medical Inc., Сан-Диего, Калифорния, США). Давление было измерено 3 раза за короткое время; затем за артериальное давление принималось среднее значение.
В случае бесплатного питания и воды суточная моча беременных крыс собиралась на 10, 13, 16 и 19 день беременности, а содержание белка определялось в нефрологическом отделении Народной больницы Уханьского университета.
Образец коллекции
Беременных крыс анестезировали 3% пентобарбиталом натрия на 21-й день беременности. Периферическую кровь крыс консервировали, центрифугировали для взятия сыворотки и хранили в холодильнике при -20 ° C для ожидания. Затем плод крысы и плаценту извлекали путем кесарева сечения, плодную оболочку и соединенную пуповину удаляли, и пуповину, соединенную с плодом крысы, отрезали. Плаценту и плод крысы помещали на асептическую марлю для сушки крови и околоплодных вод соответственно, а затем помещали на аналитические весы для взвешивания веса. Одна часть тканей плаценты была зафиксирована 4% параформальдегидом, дегидратирована этанолом, очищена ксилолом, залита парафином и подвергнута непрерывному поперечному срезу (5 мкм) для окрашивания гематоксилин-эозином (HE) и терминального дезоксинуклеотидилтрансферазо-опосредованного дезоксиуридинтрифосфат-биотина. окрашивание по метке конца никеля (TUNEL). Остальные хранили при -80 ° C для определения количественной полимеразной цепной реакции с обратной транскрипцией (RT-qPCR), вестерн-блоттинга и иммуноферментного анализа (ELISA).
ELISA
Содержание фактора некроза опухоли-α (TNF-α), интерлейкина (IL) -1β и IL-6 в сыворотке крови исследовали с помощью ELISA. Концентрации TNF-α, IL-1 и IL-6 определяли в соответствии с инструкциями набора (R&D Systems, Миннеаполис, Миннесота, США). Значения оптической плотности (OD) (490 нм) тестировали с помощью считывающего устройства для микропланшетов (Thermo Fisher Scientific, MA, США). Соответствующая стандартная кривая была получена с использованием значения OD в качестве абсциссы и концентрации соответствующего стандартного образца в качестве ординаты. Концентрации TNF-α, IL-1β и IL-6 рассчитывались по стандартной кривой.
Окрашивание HE
Образцы парафина тканей плаценты очищали в ксилоле, обезвоживали обычным градиентным спиртом, окрашивали гематоксилином, дифференцировали 1% -ным спиртом соляной кислоты и возвращали в голубой цвет 1% -ным раствором аммиачной воды. Затем ткани контрастировали 1% раствором эозина, обезвоживали (75%, 90%, 95% этанол, соответственно, абсолютный этиловый спирт) и очищали ксилолом, сушили, блокировали и наблюдали под электронным микроскопом.
Окрашивание TUNEL
Залитые парафином срезы обычно депарафинизировали и обезвоживали в соответствии с инструкциями, а затем апоптоз выявляли с помощью набора TUNEL (Nanjing Kejin Biotechnology Co., Ltd., Цзянсу, Китай). 4,6-Диамино-2-фенилиндол (Shanghai Baitai Biotechnology Co., Ltd., Шанхай, Китай) использовали для наблюдения TUNEL-положительных клеток с помощью флуоресцентного микроскопа (Nikon, Токио, Япония) [18].
RT-qPCR
Ткани плаценты взвешивали. На 50–100 мг тканей плаценты добавляли 1 мл TRIzol (Invitrogen, Carlsbad, California, USA) и полностью растворяли. К тканям добавляли 200 мкл хлороформа и центрифугировали при 4 ° C, 12000 об / мин для экстракции общей РНК. Концентрацию и чистоту РНК определяли на спектрофотометре нуклеиновых кислот белка DU-800 (Beckman). U6 и β-актин использовали в качестве контроля загрузки. Праймеры для ПЦР были разработаны и составлены компанией Shanghai Sangon Biotechnology Co. Ltd. (Шанхай, Китай). Последовательности праймеров перечислены в таблице 1. РНК была преобразована в кДНК на основе инструкций набора для обратной транскрипции РНК (Sangon). ПЦР амплифицировали, и продукты проверяли электрофорезом в агарозном геле. Данные были рассчитаны с помощью 2 −ΔΔCt метод.
Вестерн-блоттинг
Общий белок тканей плаценты экстрагировали с помощью буфера для лизиса клеток для анализа радиоиммунопреципитации (Beyotime). HucMSC-Exo использовали для извлечения буфера, который центрифугировали при 14000 об / мин. Супернатант сохраняли для тестирования экспрессии белка экзосомального маркерного белка (CD81, CD63 и CD9) в сыворотке. Концентрацию белка определяли с помощью набора бицинхониновой кислоты (Beyotime, P0010). Образец загружали в соответствии с количественными результатами белка, обрабатывали электрофорезом в 10% -ном додецилсульфат-полиакриламидном геле и переносили на мембрану. Мембрану блокировали 5% обезжиренным молоком, зондировали первичными антителами LEP, CD63, CD81, CD9 и β-актином (4 мл, 1:1000, Santa Cruz Biotechnology, Inc, Санта-Крус, Калифорния, США), повторно зондировали с 4 мл вторичных антител козы против кроличьего IgG / пероксидазы хрена, экспонирование и проявление. β-Актин использовался в качестве внутреннего стандарта. Значение серого анализировали с помощью программного обеспечения для гель-графического анализа Image Lab.
Анализ репортерного гена с двойной люциферазой
Программное обеспечение для онлайн-прогнозирования https://cm.jefferson.edu/ было адаптировано для прогнозирования целевого отношения между miR-18b-3p и LEP, а также сайта связывания miR-18b-3p и 3'-нетранслируемой области LEP (UTR ). Была составлена последовательность промоторной области LEP 3'UTR, содержащая сайт связывания miR-18b-3p. Были сконструированы плазмида LEP 3'UTR дикого типа (WT) и мутантный тип LEP 3'UTR (MUT). Рекомбинантные плазмиды были названы LEP 3'UTR-WT и LEP 3'UTR-MUT соответственно. Культивированные клетки 293T котрансфицировали миметиком miR-18b-3p и миметиком LEP 3′UTR-WT, миметиком miR-18b-3p и миметиком LEP 3′UTR-MUT, миметиком NC и LEP 3′UTR-WT, миметиком NC и ЛЭП 3'УТР-МУТ на 30 ч. Затем были собраны клетки 293T. Активность люциферазы светлячков и рениллы в клетках измеряли с помощью измерений люминесценции в соответствии с набором для обнаружения репортерного гена двойной люциферазы (Promega, Мэдисон, Висконсин, США).
Анализ РНК с понижением давления
Биотинилированные РНК-зонды (Bio-miR-NC, Bio-miR-18b-3p и Bio-miR-18b-3p-Mut) инкубировали с лизатом клеток 293T и экстрагировали с использованием магнитных шариков, конъюгированных с антибиотиком стрептомицином. Эксперимент проводился в соответствии с инструкциями наборов для раскрытия магнитной РНК Pierce (Pierce, IL, USA). РНК элюировали и очищали с использованием TRIzol (Pierce). Обогащение LEP комплексом РНК количественно оценивали с помощью RT-qPCR, как описано ранее [19].
Статистический анализ
Все данные были эксплицированы с помощью программного обеспечения SPSS 21.0 (IBM Corp. Armonk, NY, США). Данные измерений были указаны как среднее ± стандартное отклонение. Данные были получены по независимой выборке t тест для сравнения двух групп, тогда как сравнения между несколькими группами оценивали с помощью одностороннего дисперсионного анализа (ANOVA) с последующим апостериорным тестом Тьюки. Критерий статистической значимости был установлен на уровне p <0,05.
Результаты
Морфология и идентификация HucMSC и HucMSC-Exos
Массы ткани пуповины наблюдали под инвертированным микроскопом. Видно, что клетки выползали из тканевой массы на 3-е сутки; клетки проявляли форму веретена и нитевидность, а также росли как колония примерно через 5 дней. При культивировании до пассажа 3 морфология клеток была однородной, длинной веретенообразной и сходной с морфологией фибробластов, а расположение было правильным (рис. 1а). После 2 недель адипогенной дифференцировки hucMSC в цитоплазме образовались липидные капли, и липидные капли показали структуру Кранца под инвертированным микроскопом (рис. 1b), что позволяет предположить, что изолированный культивированный hucMSC обладал способностью адипогенной дифференцировки. Через 2 недели после остеогенной дифференцировки большое количество коричневых кальциевых узелков можно было увидеть под инвертированным микроскопом (рис. 1c), что указывает на то, что изолированные культивируемые hucMSC обладают способностью к остеогенной дифференцировке. Проточный цитометр был адаптирован для проверки иммунофенотипа клеток, и результаты показали, что клетки сверхэкспрессируют поверхностный маркер CD73, CD105 и CD166 МСК (рис. 1d).
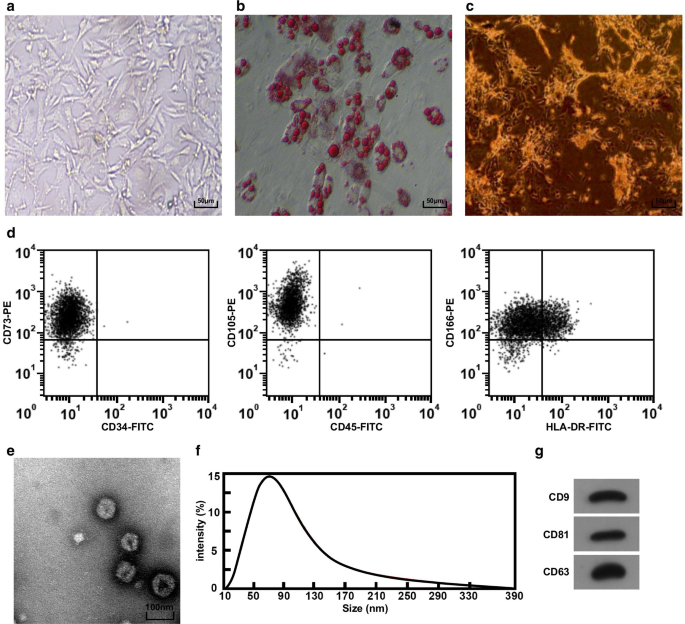
Морфология и идентификация hucMSC и hucMSC-Exos. а Морфология hucMSC под инвертированным микроскопом, b hucMSC тестировали путем окрашивания масляным красным О. c hucMSC тестировали окрашиванием ALP. г Для определения иммунофенотипа использовали проточную цитометрию. е Форма и размер hucMSC-Exos, наблюдаемые с помощью ПЭМ. е Обнаружение гранулометрического состава экзосом с помощью анализа Nanosight. г Экспрессия белков CD9, CD81 и CD63 в hucMSC-Exos была обнаружена с помощью вестерн-блоттинга
Морфологию hucMSC-Exos наблюдали с помощью ПЭМ, и результаты показали, что экзосомы были круглыми или овальными с низкой центральной плотностью и густым окрашиванием с обеих сторон (рис. 1e). Для анализа размера частиц экзосом использовался анализ с помощью нанозрения, и результаты показали, что размер частиц в основном распределялся между 40 и 100 нм, более концентрированным около 80 нм (рис. 1f). Вестерн-блоттинг показал, что все поверхностные маркеры CD81, CD63 и CD9 экспрессируются в hucMSC-Exos (рис. 1g).
Восстановление miR-18b-3p снижает патологические характеристики крыс с PE
Результаты САД и 24-часовой протеинурии показали, что:не было существенной разницы в САД и 24-часовой протеинурии в 6 группах до введения (10-й день беременности). САД и 24-часовая протеинурия на 19-й день беременности не показали очевидных различий у нормальных крыс. У PE крыс или PE крыс, получавших miR-NC, miR-18b-3p агомир, miR-18b-3p антагомир, miR-18b-3p антагомир + si-LEP, si-NC или si-LEP, SBP и 24 ч. протеинурия стала нарастать на 13-й день беременности. Не было четкой разницы в уровне САД и 24-часовой протеинурии на 16-й и 19-й день беременности у крыс с PE, получавших агомир miR-18b-3p и si-LEP. Крысы PE имели повышенное САД и 24-часовую протеинурию на 19-й день беременности; это увеличение снижалось за счет повышения miR-18b-3p, но дополнительно усиливалось ингибированием miR-18b-3p; Снижение LEP отменяет роль подавления miR-18b-3p в SBP и 24-часовой протеинурии на 19 день беременности у крыс с PE (рис. 2a, b).
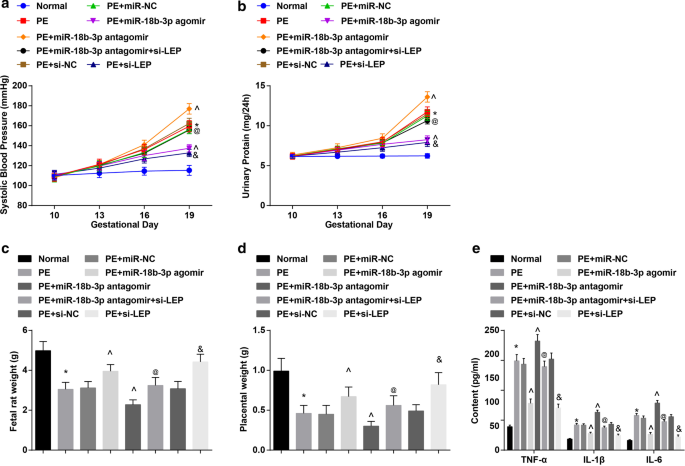
Восстановление miR-18b-3p снижает патологические характеристики крыс с PE. а Результаты САД у крыс. б Результаты суточной протеинурии у крыс. c Изменение веса плодов крысы. г Изменение веса плаценты у крыс. е Изменения воспалительных факторов в сыворотке крови выявляли с помощью ИФА. нет =10, * p <0,05 по сравнению с нормальной группой. ^ p <0,05 по сравнению с группой PE + miR-NC. @ p <0,05 по сравнению с группой антагомира PE + miR-18b-3p. & p <0,05 по сравнению с группой PE + si-NC. Данные измерений были представлены как среднее ± стандартное отклонение, а сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа с последующим тестом Тьюки
У крыс PE снизилась масса плода крысы и плаценты; повышенная регуляция miR-18b-3p или пониженная регуляция LEP увеличивалась, тогда как пониженная регуляция miR-18b-3p снижала вес плода крысы и плаценты у крыс PE; Подавление LEP обратило вспять эффект ингибирования miR-18b-3p на вес плода крысы и плаценты у крыс PE (рис. 2c, d).
Воспалительных факторов в сыворотке крови крыс PE не обнаружено. Было обнаружено, что содержание TNF-α, IL-1β и IL-6 увеличивалось у крыс PE; Повышение miR-18b-3p или ингибирование LEP подавлялось, тогда как уменьшение miR-18b-3p способствовало содержанию TNF-α, IL-1β и IL-6; Эффект ингибированной miR-18b-3p на содержание TNF-α, IL-1β и IL-6 был отменен истощением LEP (рис. 2e).
Избыточная экспрессия miR-18b-3p улучшает гистопатологические изменения тканей плаценты у крыс с PE
У нормальных крыс ворсинки плаценты были богаты кровеносными сосудами и имели четкую структуру, синцитиотрофобласты были основными трофобластами в ворсинах плаценты, а цитотрофобластов было меньше. У крыс PE или крыс PE, получавших miR-NC, miR-18b-3p антагомир, si-NC или miR-18b-3p антагомир + si-LEP, количество ворсин плаценты уменьшилось, структура была размытой и атрофированной, некоторые ворсинки был проведен фибриноидный некроз, и количество узелков синцитиотрофобластов в ворсинах плаценты увеличилось, и большинство ворсинок были незрелыми. Количество трофоцитов было уменьшено, и патологические изменения были облегчены у крыс с PE, получавших агомир miR-18b-3p и si-LEP (рис. 3a).
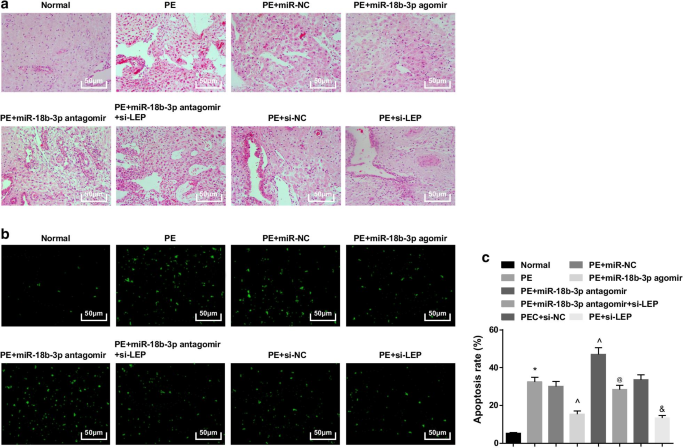
Сверхэкспрессия miR-18b-3p улучшает патологические изменения и подавляет апоптотические клетки тканей плаценты у PE крыс. а Окрашивание HE использовали для проверки патологических свойств тканей плаценты. б Окрашивание TUNEL применяли для определения апоптотических клеток тканей плаценты у крыс PE. c Скорость апоптоза клеток определяли окрашиванием TUNEL. нет =10, * p <0,05 по сравнению с нормальной группой. ^ p <0,05 по сравнению с группой miR-NC. @ p <0,05 по сравнению с группой антагомира miR-18b-3p. & p <0,05 по сравнению с группой si-NC. Данные измерений были представлены как среднее ± стандартное отклонение, а сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа с последующим тестом Тьюки
Окрашивание TUNEL показало, что можно было увидеть небольшое количество апоптотических клеток. Крысы PE имели повышенное количество апоптозных клеток, которое снижалось за счет повышения miR-18b-3p и подавления LEP, и дополнительно усиливалось ингибированием miR-18b-3p; Подавление LEP также обращает вспять эффект ингибирования miR-18b-3p на количество апоптотических клеток у PE крыс (рис. 3b, c).
Взятые вместе, крысы с повышенной регуляцией miR-18b-3p или ингибированным LEP имели пониженную степень прогрессирования PE в гистологии, а замалчивание LEP могло отменить терапевтический эффект ингибированного miR-18b-3p.
miR-18b-3p подавляется, в то время как LEP активируется в тканях плаценты PE крысы, а miR-18b-3p нацеливается на LEP
Основываясь на приведенных выше результатах, подавление LEP обращало терапевтический эффект подавления miR-18b-3p на крыс с PE в патологии и гистологии; Таким образом, мы предположили, что miR-18b-3p может быть связана с LEP.
Вестерн-блоттинг и RT-qPCR показали, что у крыс PE снизились уровни экспрессии miR-18b-3p и увеличились уровни экспрессии LEP; обработка агомиром miR-18b-3p активировала miR-18b-3p и подавляла LEP у крыс PE, тогда как обработка антагомира miR-18b-3p увеличивала экспрессию LEP; Подавление LEP обращает вспять промоторный эффект снижения miR-18b-3p на экспрессию LEP у крыс PE (рис. 4a – c).
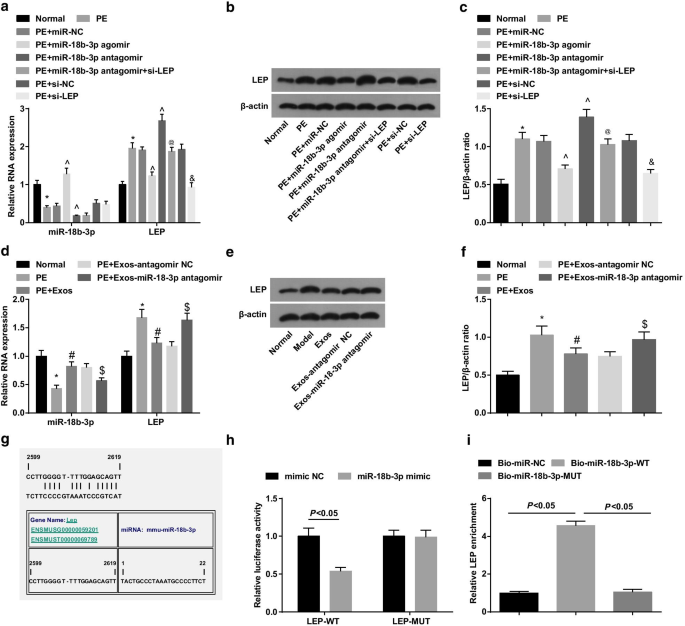
miR-18b-3p подавляется, а LEP активируется в тканях плаценты крыс PE. а Экспрессию мРНК miR-18b-3p и LEP в тканях плаценты определяли с помощью RT-qPCR. б Белковая полоса LEP в тканях плаценты. c Был проведен вестерн-блоттинг для определения экспрессии белка LEP в тканях плаценты. г Экспрессию мРНК miR-18b-3p и LEP в тканях плаценты после обработки экзосом выявляли с помощью RT-qPCR. е Белковая полоса LEP в тканях плаценты после обработки экзосомами. е Проводили вестерн-блоттинг для обнаружения экспрессии белка LEP после обработки экзосом. г Сайты связывания miR-18b-3p и LEP рассчитаны с помощью онлайн-программного обеспечения. ч Целевые отношения между miR-18b-3p и LEP подтверждены анализом двойного люциферазного репортерного гена. я Направляющая взаимосвязь между miR-18b-3p и LEP подтверждена анализом РНК с понижением. нет =10, * p <0,05 по сравнению с нормальной группой. ^ p <0,05 по сравнению с группой miR-NC. @ p <0,05 по сравнению с группой антагомира miR-18b-3p. & p <0,05 по сравнению с группой si-NC. # p <0,05 по сравнению с группой PE. $ p <0,05 по сравнению с группой ПЭ + экзос-антагомир NC. Данные измерений были представлены как среднее ± стандартное отклонение, а сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа с последующим тестом Тьюки
Вестерн-блоттинг и RT-qPCR использовали для изучения роли экзосом у PE крыс. Результаты показали, что экзосомы активировали miR-18b-3p и подавляли LEP у PE крыс, что указывает на подавляющий эффект экзосом на развитие PE. Более того, экзосомы, передающие антагомир miR-18b-3p, индуцировали подавление miR-18b-3p и активацию LEP у PE крыс (рис. 4d-f).
Целевое соотношение между miR-18b-3p и LEP было предсказано программой онлайн-прогнозирования биоинформатики https://cm.jefferson.edu/ (рис. 4g). Анализ двойного люциферазного репортерного гена показал, что миметик miR-18b-3p снижает люциферазную активность LEP 3'UTR-WT, но не влияет на активность LEP 3'UTR-MUT (фиг. 4h). Кроме того, анализ РНК pull-down показал, что обогащение LEP увеличивалось WT-биотинилированным miR-18b-3p (фиг. 4i). Эти данные показали, что LEP является целевым геном miR-18b-3p.
hucMSC-Exos ослабляет патологические характеристики крыс с PE
Результаты САД и 24 ч показали, что не было значительной разницы в САД и 24-часовой протеинурии в 5 группах до введения (10 день беременности). САД и 24-часовая протеинурия на 19-й день беременности не показали четких различий у нормальных крыс. У крыс с PE САД и суточная протеинурия начали повышаться на 13-й день беременности. Не было четкой разницы в уровне САД и 24-часовой протеинурии на 16-й и 19-й день беременности у крыс с PE, получавших hucMSC-Exos и hucMSC-Exos, передающих антагомир NC. САД и 24-часовая протеинурия усилились на 19 день беременности у крыс PE, тогда как это повышение было снижено путем инъекции hucMSC-Exos. Ингибирование miR-18b-3p обращало эффект hucMSC-Exos на САД и 24-часовую протеинурию на 19 день беременности у крыс с PE (рис. 5a, b).
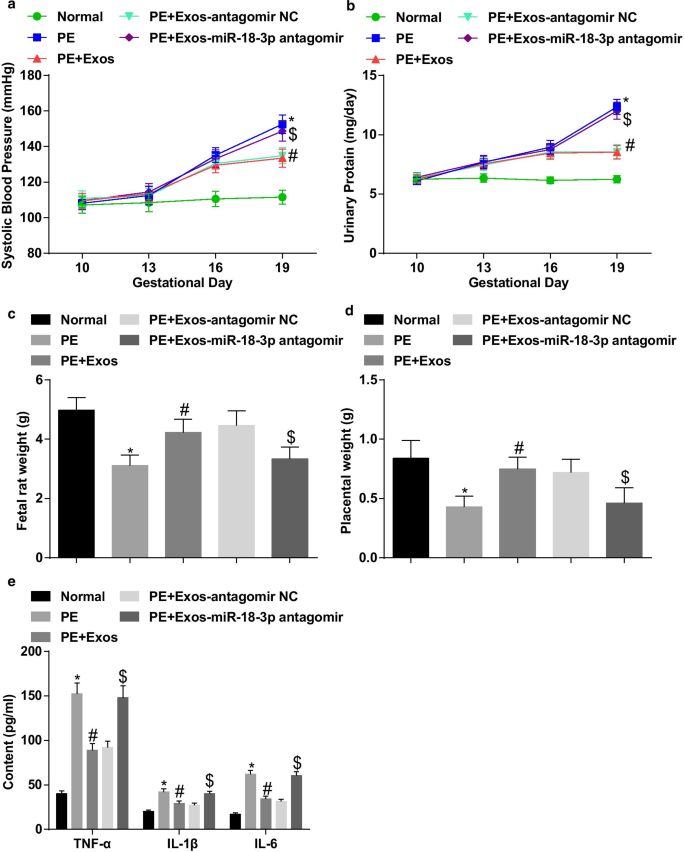
hucMSC-Exos ослабляет патологические характеристики крыс с PE. а Результаты САД у крыс после обработки экзосомами. б Результаты суточной протеинурии у крыс после обработки экзосомами. c Изменение веса плодов крысы после обработки экзосомами. г Изменение веса плаценты у крыс после обработки экзосомами. е Изменения факторов воспаления в сыворотке крови после обработки экзосом определяли с помощью ИФА. нет =10, * p <0,05 по сравнению с нормальной группой. # p <0,05 по сравнению с группой PE. $ p <0,05 по сравнению с группой ПЭ + экзос-антагомир NC. Данные измерений были представлены как среднее ± стандартное отклонение, а сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа с последующим тестом Тьюки
Был измерен вес плода крысы и плаценты, и мы обнаружили, что у крыс PE уменьшился вес плода крысы и плаценты; Подавление miR-18b-3p устраняет роль hucMSC-Exos в массе плода крысы и плаценты у крыс с PE (рис. 5c, d).
Воспалительные факторы в сыворотке крови выявляли с помощью ИФА. Содержание TNF-α, IL-1β и IL-6 заметно увеличивалось у крыс PE. Обработка экзосом уменьшала содержание TNF-α, IL-1β и IL-6 в сыворотке PE крыс, которое было увеличено путем инъекции экзосом, ингибирующих miR-18b-3p (рис. 5e).
Экзосомы облегчают патологические изменения и подавляют апоптоз тканей плаценты у крыс с PE
У нормальных крыс ворсинки плаценты были обильны кровеносными сосудами с четкой структурой, синцитиотрофобласты были основным трофобластом ворсин плаценты, а цитотрофобластов было меньше. У крыс с PE и крыс с PE, получавших hucMSC-Exos-miR-18b-3p-антагомир, количество ворсин плаценты уменьшилось, структура была нечеткой и атрофированной, на некоторых ворсинках был представлен фибриноидный некроз, а количество узелков синцитиотрофобластов в плаценте ворсинки увеличены, и большинство ворсинок были незрелыми. Патологические изменения улучшились у PE крыс, получавших hucMSC-Exos или hucMSC-Exos-antagomir NC, по сравнению с PE крысами и PE крысами, получавшими антагомир hucMSC-Exos-miR-18b-3p (рис. 6a).
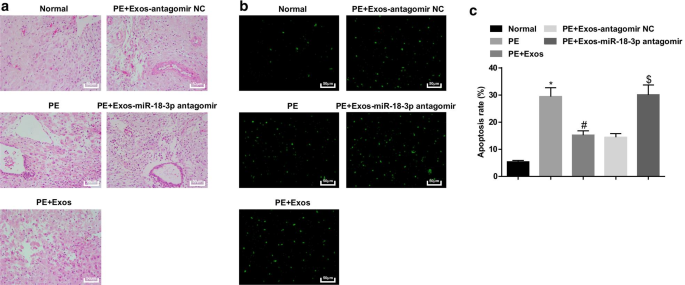
Экзосомы облегчают патологические изменения и уменьшают апоптоз клеток тканей плаценты у крыс PE. а Окрашивание HE использовали для тестирования патологических свойств тканей плаценты у крыс PE после обработки экзосомами. б Окрашивание TUNEL применяли для определения апоптотических клеток тканей плаценты у крыс PE после обработки экзосомами. c Скорость апоптоза клеток определяли окрашиванием TUNEL. нет =10, * p <0,05 по сравнению с нормальной группой. # p <0,05 по сравнению с группой PE. $ p <0,05 по сравнению с группой ПЭ + экзос-антагомир NC. Данные измерений были представлены как среднее ± стандартное отклонение, а сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа с последующим тестом Тьюки
Окрашивание TUNEL показало, что у нормальных крыс можно было увидеть небольшое количество апоптотических клеток. Крысы PE имели повышенное количество апоптотических клеток, а снижение miR-18b-3p обращало воздействие hucMSC-Exos на количество апоптотических клеток в тканях плаценты крыс PE (рис. 6b, c).
Обсуждение
ПЭ представляет собой мультисистемное расстройство беременности, характеризующееся протеинурией и повышенным артериальным давлением или другими неблагоприятными состояниями, и связано с широким спектром материнской эндотелиальной дисфункции [20]. Сообщалось, что hucMSC-Exo улучшал морфологию плацентарной ткани у крыс PE за счет подавления апоптоза клеток и облегчения ангиогенеза в плацентарной ткани дозозависимым образом [8]. В исследовании сообщается, что экспрессия miR-18b влияет на миграцию, жизнеспособность и инвазию клеток при PE [12]. Более того, было подтверждено повышение материнской концентрации LEP и гипометилирование LEP в плаценте при раннем начале PE [21]. Настоящее исследование было разработано для изучения влияния экзосом и нацеленных на miR-18b-3p LEP на возникновение PE. Результаты этого исследования показали, что экзосомная miR-18b-3p, происходящая из hucMSC, ингибирует прогрессию PE за счет снижения LEP.
Основываясь на наших выводах, miR-18b-3p снижал, а LEP повышал в тканях плаценты крыс PE. Подобно нашему исследованию, экспрессия мРНК miR-18b была заметно подавлена в тканях плаценты PE по сравнению с таковой в нормальных тканях плаценты [12]. Кроме того, исследование показало, что содержание miR-18b резко снижено в тканях злокачественной меланомы по сравнению с соответствующими соседними неопухолевыми тканями [22]. Другое исследование подтвердило, что экспрессия LEP в плаценте была повышена у недоношенных PE по сравнению с контролем [23]. Более того, исследование показало, что экспрессия LEP явно повышена в преэклампсической плаценте [15]. Эта литература предоставила нам теоретическую основу для изучения аномальной экспрессии miR-18b-3p и LEP при PE. Более того, с помощью биоинформатического программного обеспечения было предсказано, что LEP нацелен на miR-18b-3p, и эта связь нацеливания была дополнительно подтверждена анализом двойного люциферазного репортерного гена в нашем исследовании. Исследование показало, что LEP является мишенью для всех трех miRNAs (miR-1301, miR-223 и miR-224) при раннем начале PE [16]. Другое исследование показало, что LEP снижает экспрессию miR-93 при остеоартрите и ревматоидном артрите [24]. Однако связывание miR-18b-3p и LEP при заболеваниях человека, особенно при PE, остается малоизученным, что является новизной данного исследования. Более того, результаты нашего исследования показали, что экзосомы увеличивают miR-18b-3p и снижают LEP в тканях плаценты PE. Ранее было задокументировано, что экспрессия miR-18b-5p заметно повышается в экзосомах плазмы колоректального рака [25], в то время как взаимосвязь между hucMSC-Exos и miR-18b-3p / LEP при ПЭ требует дальнейшего изучения.
Кроме того, результаты нашего исследования показали, что восстановление miR-18b-3p снижает САД и 24-часовую протеинурию у крыс PE, увеличивает вес плаценты, снижает содержание TNF-α, IL-1β и IL-6 в сыворотке и тканях плаценты. а также подавление апоптоза клеток. Эти данные показали, что повышение уровня miR-18b-3p способствует облегчению симптомов и патологических изменений при ПЭ. Было продемонстрировано, что стабильная повышающая регуляция miR-18b вызывает эффективную активность ингибитора опухоли, такую как подавление жизнеспособности клеток меланомы, индукция апоптоза и уменьшение роста опухоли in vivo [26]. Другой результат нашего исследования заключался в том, что hucMSC-Exos снижал САД и 24-часовую протеинурию у крыс с PE, увеличивал вес плаценты, снижал содержание TNF-α, IL-1β и IL-6 в сыворотке и тканях плаценты, а также подавлял клеточную активность. апоптоз. Результаты настоящего исследования показали, что модели крыс с PE, обработанные экзосомами, показали увеличение количества и качества плодов, качества плаценты, но апоптоз клеток был значительно снижен [8]. Интересно, что предыдущее исследование продемонстрировало, что добавление экзосом плода крупного рогатого скота снижает содержание макрофагов TNF-α и IL-6 [27]. Исследование показало, что очищенные экзосомы подавляют продукцию IL-1β в макрофагах, стимулированных липополисахаридом / нигерицином [28]. Кроме того, Нонг и др. предположили, что воспалительные маркеры, такие как TNF-α и IL-6, резко снизились после введения экзосом, продуцируемых индуцированными человеком плюрипотентными стволовыми клетками МСК [29]. В статье говорится, что САД было заметно повышено в группе женщин, у которых позже развилась ПЭ [30, 31]. Было показано, что пациенты с ПЭ были положительно связаны с САД, диастолическим артериальным давлением и протеинурией [32]. Кроме того, недавнее исследование предоставило доказательство того, что протеинурия усиливается с приближением срока беременности у женщин с ПЭ [33]. Важным открытием было то, что крысы из группы PE имели повышенный TNF-α по сравнению с группой здоровых беременных [34]. Другое исследование подтвердило, что уровни IL-6 и IL-1β в сыворотке крови у женщин с ПЭ были явно повышены по сравнению с контрольной группой [35]. Вышеприведенные данные свидетельствуют о том, что у пациентов с ТЭЛА обычно наблюдается высокое САД, протеинурия и уровни воспалительных факторов. Таким образом, из наших результатов можно сделать вывод, что происходящий из hucMSC экзосомальный miR-18b-3p оказывает терапевтическое действие на ПЭ.
Заключение
В заключение, наше исследование предоставляет доказательства того, что происходящие из hucMSC экзосомы активируют miR-18b-3p, который нацелен на LEP, подавляя содержание воспалительных факторов и снижая скорость апоптоза клеток в тканях плаценты PE крыс, тем самым подавляя возникновение PE. Таким образом, экзосомальная miR-18b-3p может быть потенциальным кандидатом для лечения PE посредством нацеливания на LEP. Это исследование впервые выявило роль происходящей из hucMSC экзосомальной miR-18b-3p в нацеливании на LEP во время развития PE, что дало новое понимание лечения PE. Тем не менее, из-за ограниченности известных исследований, исследование необходимо строго контролировать и надлежащим образом сообщать в будущих клинических испытаниях.
Доступность данных и материалов
Не применимо.
Наноматериалы
- Как кадмий вредит организму человека?
- Искусственные нейроны могут быть столь же эффективны, как и человеческий мозг
- Появление «самоуправляемой» цепочки поставок
- Критический человеческий элемент в стратегии кибербезопасности
- В условиях пандемии приверженность заводов правам человека падает
- Будущее автоматизации в авиационной отрасли
- Биометрическая система безопасности на основе сердцебиения человека
- Делавэр:идеальное сочетание человека и ИИ
- Человеческий элемент автоматизации
- Человеческий фактор в контроле металлов