


HDAC1-опосредованная MicroRNA-124-5p регулирует NPY, чтобы влиять на способности к обучению и памяти у крыс с депрессией
Аннотация
Исследования гистоновых деацетилаз (HDAC) при депрессии проводились чрезмерно, но не так много на HDAC1. В этой связи настоящее исследование запускается для раскрытия механизма оси HDAC1 / микроРНК (miR) -124-5p / нейропептид Y (NPY) при депрессии. Крыс Sprague Dawley стимулировали хроническим непредсказуемым умеренным стрессом для создания моделей депрессии. Подавленным крысам вводили ингибированный HDAC1 или подавленный miR-124-5p для изучения их роли в массе тела, способности к обучению и памяти, окислительному стрессу и воспалению в сыворотке и экспрессии нейромедиаторов в тканях гиппокампа. Экспрессия MiR-124-5p, HDAC1 и NPY в гиппокампе была протестирована. Также были подтверждены взаимодействия экспрессии miR-124-5p, HDAC1 и NPY. Более высокие уровни экспрессии miR-124-5p и HDAC1 и более низкие уровни NPY были обнаружены в гиппокампе депрессивных крыс. Ингибирование miR-124-5p или подавление HDAC1 ослабляло способности к обучению и памяти и увеличивало массу тела крыс с депрессией. Нокдаун miR-124-5p или ингибирование HDAC1 подавлял окислительный стресс и воспаление и способствовал экспрессии нейротрансмиттеров у депрессивных крыс. HDAC1 опосредует miR-124-5p для регуляции NPY. Нокдаун NPY отменял защитные эффекты ингибированного miR-124-5p на депрессивных крысах. Наше исследование показывает, что подавление miR-124-5p или HDAC1 активирует NPY, улучшая память и способности к обучению у мышей с депрессией, что может обновить существующие знания о депрессии и предоставить новый справочный материал для лечения депрессии.
Введение
Депрессия относится к тяжелому инвалидизирующему психозу, который вызывает тяжелое экономическое бремя и социальные последствия во всем мире [1]. Депрессия характеризуется физиологическими, когнитивными и поведенческими изменениями, которые угрожают общему здоровью пациентов [2]. Лечение, доступное для депрессии, было разработано с применением кетамина в качестве быстродействующего антидепрессанта и усовершенствованием оборудования, способного выборочно обслуживать активность популяций нейрональных подтипов [3]. Более того, универсальные наноархитектуры, такие как углеродные точки, применимы при неврологических расстройствах [4]. Однако неадекватное лечение депрессии может привести к плохой успеваемости, поведенческим дисфункциям, физическим заболеваниям, злоупотреблению психоактивными веществами и даже к самоубийству [5]. Таким образом, существует острая необходимость в поиске инновационных средств лечения депрессии.
Ранее документально подтверждено, что уровень микроРНК (miR) -124 повышается в плазме крови при депрессии и антидепрессантах [6]. Во время хронической непредсказуемой депрессии, вызванной умеренным стрессом (CUMS), экспрессия miR-124 в гиппокампе обнаруживает динамические вариации, указывая на различные патологические изменения на разных стадиях депрессии [7]. Кроме того, miR-124 является кандидатом на улучшение депрессивно-подобного поведения при большом депрессивном расстройстве [8]. Кроме того, подавление miR-124 служит антидепрессантом в префронтальной коре, что отражается в ослабленном депрессивноподобном поведении мышей [9]. Гистондеацетилаза 1 (HDAC1) является медиатором miR-124 [10], который формирует мультибелковый комплекс для регуляции транскрипции и эпигенетической модификации [11]. Насколько нам известно, нарушение активности HDAC1 способствует устойчивости при большом депрессивном расстройстве [12]. Более того, подавление HDAC1 в головном мозге может улучшить расстройства настроения и другие нейропластически измененные заболевания мозга [13]. Ингибиторы HDAC могут ослаблять тревожное поведение и поведение, связанное с употреблением алкоголя, и могут повышать экспрессию нейропептида Y (NPY) [14]. Также нарушение активности HDAC может приводить к увеличению экспрессии NPY в центральном ядре миндалины и медиальном ядре миндалины [15]. NPY - гипоталамический орексигенный нейропептид, которого достаточно для предотвращения тревожности, социальных расстройств и депрессивных симптомов [16]. Кроме того, было доказано, что NPY и его рецепторы оказывают противовоспалительное и антидепрессивное действие на моделях индуцированной липополисахаридами депрессии на крысах [17]. Взятые вместе, комбинированное взаимодействие оси HDAC1 / miR-124-5p / NPY при депрессии неоднозначно. Таким образом, это исследование направлено на раскрытие механизма этой оси для изучения кандидата на лечение депрессии.
Материалы и методы
Этика
Эксперименты на животных, участвовавшие в этом исследовании, соответствовали требованиям этики экспериментальных животных Первой дочерней больницы Харбинского медицинского университета. Эксперименты были оптимизированы для улучшения условий кормления экспериментальных животных, уменьшения количества используемых животных и облегчения страданий животных.
Экспериментальные животные
Крысы-самцы Sprague – Dawley (SD), не содержащие специфических патогенов (SPF), весом 200–220 г были предоставлены Исследовательским центром животных Первой дочерней больницы Харбинского медицинского университета (Харбин, Китай). Крыс содержали в среде класса SPF (22–24 ° C, влажность 60–64%, 12-часовой световой и темный циклы). За исключением периода моделирования и других конкретных моментов времени, крысы могли свободно получать воду и пищу.
Группировка крыс и создание модели
Смоделированные крысы были случайным образом разделены на 8 групп ( n =10):нормальная группа (крысы без какого-либо лечения), группа CUMS (крысы с CUMS-индуцированной депрессией), группа sh-отрицательного контроля (NC) (крысы с CUMS-индуцированной депрессией, которым вводили лентивирус sh-HDAC1 NC), группа sh-HDAC1 (Крысы с CUMS-индуцированной депрессией, которым вводили лентивирус sh-HDAC1), группа анти-miR-NC (крысы с CUMS-индуцированной депрессией, которым вводили лентивирус NC против miR-124-5p), группа анти-miR-124-5p (CUMS- крысы с индуцированной депрессией, которым вводили лентивирус против miR-124-5p), группа против miR-124-5p + sh-NC (крысы с индуцированной CUMS депрессией, которым вводили лентивирус против miR-124-5p и NC лентивируса sh-NPY) и группа анти-miR-124-5p + sh-NPY (крысы с CUMS-индуцированной депрессией, которым вводили лентивирус против miR-124-5p и лентивирус sh-NPY).
Модели крыс с депрессией, вызванной CUMS, были созданы по методу Виллнера [18]. Крыс в нормальной группе кормили пищей и водой без каких-либо стимулов, в то время как крысы в других 7 группах получали CUMS в течение 35 дней в независимых клетках. Этих крыс стимулировали плаванием в ледяной воде при 4 ° C в течение 5 минут в контейнере (глубина воды 30 см), дневной и ночной инверсией (крысы оставались в темноте в течение 12 часов в течение дня и в светлом помещении, освещенном светом). лампы накаливания на 12 часов в течение ночи), зажим хвоста зажимом для ласточкина 30 с, встряхивание 1 мин, 24-часовое голодание и водная депривация, 30-минутная ультразвуковая стимуляция (50 Гц), влажные прокладки и кондиционирование при 17 ℃. Крыс случайным образом стимулировали одним из этих стимулов ежедневно, и в общей сложности один и тот же стимул применялся не более 5 раз.
Крыс анестезировали 2% пентобарбиталом натрия (50 мг / кг) и в двусторонний гиппокамп (переднезадний диаметр =4,8 мм, медиолатеральное расстояние =± 2,5 мм, расстояние между задней полостью и передним родничком =-3,5 мм) вводили лентивирус через 1 минуту. мкл / мин микрошприцем Hamilton с иглой 26G. Иглу держали в месте инъекции в течение 5 мин для предотвращения рефлюкса. Черепа заделали костным воском, надрезы зашили, и крыс вылечили на 7 дней [19]. лентивирус sh-HDAC1 и его NC, лентивирус против miR-124-5p и его NC, а также sh-NPY и его NC (титр:10 8 ЕД / мл) были приобретены у GenePharma Co. Ltd. (Шанхай, Китай).
Тесты поведенческих функций
Крыс взвешивали за день до моделирования CUMS (на 0-й день), после моделирования CUMS (на 36-й день) и после лентивирусного вмешательства (на 52-й день).
Тест открытого поля (OFT) мог обнаруживать вегетативное и исследовательское поведение крыс в новой среде, что обычно использовалось для оценки депрессивного поведения. Этот тест проводился в черном деревянном ящике открытого поля (100 см × 100 см × 50 см) с системой видеонаблюдения, размещенной над ящиком открытого поля для записи активности крыс. Дно открытого ящика было разделено на 25 сеток (20 см × 20 см) с белыми линиями. Крыс помещали в центральную сетку в заранее заданном случайном порядке и по количеству пересечений сетки (крысы входили в сетку с конечностями или двойными передними конечностями с одной задней конечностью) и по частоте вставания на дыбы (крысы поднимали передние конечности и стояли вертикально с одной из них). задняя конечность) записывались системой видеонаблюдения. Этот тест проводился в тихом и темном помещении. После каждого испытания бокс открытого поля промывали 75% спиртом для удаления запахов.
Ангедония была одним из важнейших симптомов депрессии. Тест предпочтения сахарозы (SPT) может оценивать депрессивное поведение крыс. Перед SPT каждой крысе давали 2 бутылки воды с сахаром (1% сахарозы, мас. / Мас.) На 24 часа и 1 бутылку стерильной воды и 1 бутылку воды с сахаром еще на 24 часа. После этого процент предпочтения сахарозы (SPP) крыс определяли следующим образом:после 23-часового голодания и лишения воды крысам давали 1 бутылку стерильной воды и 1 бутылку воды с сахаром (1% сахарозы) на 30 мин. Через 1 час расположение двух бутылок меняли, и крысы могли свободно перемещаться с этими двумя бутылками еще на 30 минут. В течение этого 1 часа измеряли расход (мл) этих двух бутылок со стерильной водой и сахарной водой (1% сахарозы) и рассчитывали SPP. SPP =потребление воды с сахаром / (потребление воды с сахаром + потребление стерильной воды) × 100%.
Тест в водном лабиринте Морриса (MWM) широко применялся для оценки способности крыс к пространственному обучению и памяти. Тест MWM проводился в круглом резервуаре для воды (150 см в диаметре и 60 см в высоту) с глубиной воды 40 см. Температуру воды поддерживали на уровне 22 ± 1 ° C, и воду окрашивали в черный цвет нетоксичным и легко очищаемым красителем. Резервуар и водная поверхность были разделены на 4 квадранта (квадрант SE, SW, NW и NE), каждый квадрант украшен соответствующим значком яркого цвета и особой формы на внутренней стене. Целевая платформа представляла собой круглую прозрачную платформу (диаметром 11 см), расположенную в центре северо-восточного квадранта и погруженную на 1,5 см ниже поверхности воды. Над центром резервуара была установлена система видеонаблюдения для записи скорости плавания, пути и времени, затрачиваемого на попадание крыс на целевую платформу и плавание в квадрантах. Крыс помещали в воду из 4 квадрантов в случайном порядке, и им позволяли исследовать и садиться на целевую платформу в течение 60 с. Время, затраченное крысами на посадку на платформу, регистрировалось как латентность побега. Если крысам не удавалось подняться на целевую платформу в течение 60 с, их направляли, и время побега регистрировалось как 60 с. После каждого теста крысам позволяли оставаться на целевой платформе в течение 15 секунд, и крыс тренировали 4 раза в день в течение 5 дней. Окончательная задержка перехода была средней задержкой перехода с 3-го по 5-е. На 6-е сутки испытаний MWM был проведен космический тест. Целевая платформа была удалена из резервуара для воды, и крысы вошли в воду из квадранта SW. Время, в течение которого крысы плавали в квадранте NE в течение 60 с, регистрировали как время исследования космоса.
Хронология этого эксперимента представлена на рис. 1.
Хронология настоящего исследования
Коллекция образцов
Через день после последнего теста MWM крыс умерщвляли и внутрибрюшинно вводили 2% пентобарбитал натрия (50 мг / кг). Грудную полость открывали для получения сердечной крови шприцем, который центрифугировали 3 раза при 3500 об / мин в течение 15 мин, супернатант хранили при -20 ° C. После сбора крови катетер вводили из левого желудочка в аорту и перфузировали 500 мл физиологического раствора для промывания цельной крови. Ткани гиппокампа отделяли и помещали в центрифужные пробирки объемом 1,5 мл, взвешивали и хранили при -80 ° C. Часть гиппокампа фиксировали 4% параформальдегидом в течение 2 часов, обезвоживали градиентом сахарозы и заливали парафином.
Определение индекса окислительного стресса и воспаления
Сыворотку повторно нагревали для обнаружения супероксиддисмутазы (SOD), малондиалхехиоза (MDA), глутатиона (GSH), интерлейкина (IL) -β, фактора некроза опухоли-α (TNF-α) и концентрации оксида азота (NO). Наборы для обнаружения SOD, MDA и GSH были предоставлены Институтом биотехнологии Beyotime (Шанхай, Китай), а наборы для иммуноферментного анализа (ELISA) для обнаружения IL-β, TNF-α и NO - Нанкинским институтом биоинженерии JianCheng (г. Нанкин, Китай)
Окрашивание гематоксилином – эозином (HE)
Парафиновые срезы депарафинизировали, гидратировали этанолом, промывали дистиллированной водой и окрашивали окрашивающим раствором гематоксилина в течение 20 мин. После этого срезы промывали водопроводной водой до посинения. Затем срезы помещали в 1% раствор соляной кислоты в этаноле на 10 с, промывали водопроводной водой и обезвоживали этанолом с последующим окрашиванием эозином в течение 2 мин, обезвоживанием высококонцентрированным спиртом и проницаемостью в ксилоле. Наконец, срезы были запечатаны и исследованы под биологическим микроскопом.
Обнаружение экспрессии нейротрансмиттера
Ткани гиппокампа взвешивали и гомогенизировали ультразвуком в физиологическом растворе (100 мкл / 10 мг). Гомогенат выдерживали при 4 ° C в течение 30 минут, центрифугировали при 12000 об / мин (4 ° C) в течение 3 минут для сбора супернатанта. Концентрацию белка в супернатанте определяли с помощью набора для определения белка бицинхониновой кислоты (BCA) (CWBIO, Пекин, Китай), а уровни экспрессии норадреналина (NE), серотонина (5-HT) и дофамина (DA) с помощью наборов для ELISA. Наборы NE, 5-HT и DA ELISA были предоставлены Liweiping Biotechnology Co., Ltd. (Пекин, Китай).
Количественная полимеразная цепная реакция с обратной транскрипцией (RT-qPCR)
РНК экстрагировали из тканей гиппокампа с помощью набора для экстракции РНК (Promega, Мэдисон, Висконсин, США), а концентрацию и чистоту РНК определяли с помощью ультрафиолетового спектрофотометра. Следуя инструкциям набора для обратной транскрипции (DRR047S, Takara, Dalian, China), была выполнена обратная транскрипция РНК в комплементарную ДНК (кДНК). мРНК обратно транскрибировали в кДНК с помощью набора для одностадийной ОТ-ПЦР GoldScript (Applied Biosystems, Карлсбад, Калифорния, США), а miРНК с помощью набора для количественного определения miRNA Hairpin-it ™ (GenePharma). Набор для обнаружения RT-qPCR (Promega) применяли для обнаружения экспрессии HDAC1, miR-124-5p и NPY в тканях. U6 был указан как внутренний контроль для miR-124-5p, тогда как β-актин для HDAC1 и NPY. Праймеры для ПЦР были получены от Sangon Biotech Co., Ltd. (Шанхай, Китай) (таблица 1). Относительная экспрессия целевых генов рассчитывалась по 2 - △△ Метод Ct.
Вестерн-блоттинг
Ткани гиппокампа разрезали на кусочки на льду и лизировали лизатом для анализа радиоиммунопреципитации (Beijing Solarbio Science &Technology Co. Ltd., Пекин, Китай) для экстракции белка. Концентрация белка определялась методом BCA. К общему белку (50 мкг) добавляли 5-кратный загрузочный буфер додецилсульфата натрия (SDS) в соотношении 1:4 и нагревали на кипящей водяной бане в течение 5 минут. Затем белок подвергали электрофорезу в полиакриламидном геле с додецилсульфатом натрия, наносили электроблоттингом на мембрану и блокировали 5% молоком в течение 1 часа. После этого белок исследовали первичными антителами против HDAC1 (1:1000, Cell Signaling Technology, Беверли, Массачусетс, США), NPY (1:800, NeoMakers, Фремонт, Калифорния, США) и β-актина (1:1000, Abcam, Cambridge, MA, UK) в течение ночи и повторно зондировали вторичным антителом, меченным пероксидазой хрена. Мембрана была разработана с помощью усиленной хемилюминесценции, а оптическая плотность была рассчитана с помощью программного обеспечения для анализа серого Quantity One. Экспрессия белка целевого гена выражалась как отношение значения серого к значению серого β-актина.
Анализ иммунопреципитации хроматина (ChIP)
Анализ ChIP выполняли в соответствии с инструкциями к набору EZ-ChIP (Millipore, Бедфорд, Массачусетс, США). Клетки HEK293T инкубировали с 1% формальдегидом в течение 10 мин и останавливали глицином. Затем клетки центрифугировали при 2000 об / мин в течение 5 минут и добавляли буфер для лизиса SDS для обработки ультразвуком. Центрифугировали при 10000 g при 4 ℃ в течение 10 минут, клетки (100 мкл) реагировали с 900 мкл буфера для разведения ChIP, 20 мкл 50 × PIC и 60 мкл агарозы ProteinA / ДНК спермы лосося при 4 ℃ в течение 1 ч и подвергали анализу. дают постоять 10 мин. Осадки центрифугировали при 700 об / мин в течение 1 мин с 20 мкл на входе. Добавляли пробирку с антителом HDAC1 (1 мкл) и антителом к иммуноглобулину G, а другую пробирку добавляли без антитела. Образцы в двух пробирках инкубировали в течение ночи, элюировали и не подвергали сшивке. После получения ДНК образец был протестирован методом RT-qPCR.
Анализ двойного люциферазного репортерного гена
На сайте биоинформатики (https://cm.jefferson.edu/rna22/Precomputed/) были проанализированы сайты связывания miR-124-5p и NPY. Плазмиды NPY-дикого типа (WT) и NPY-мутантные (MUT) были созданы Huayueyang Biotechnology Co., Ltd. (Пекин, Китай). В сочетании с имитатором NC или miR-124-5p плазмиды трансфицировали в клетки HEK293T. Активность люциферазы клеток определяли с помощью набора для определения люциферазы (BioVision) и люминометра Glomax20 / 20 (Promega).
Статистический анализ
Статистическое программное обеспечение SPSS 21.0 (IBM Corp. Armonk, NY, USA) использовалось для анализа данных. Результаты выражали как среднее значение ± стандартное отклонение. Сравнения между двумя группами были проверены t контрольная работа. Сравнения между несколькими группами оценивали с помощью одностороннего дисперсионного анализа (ANOVA), после чего проводились парные сравнения с помощью апостериорного критерия Тьюки. P представлены двусторонние тесты и P <0,05 считалось статистически значимым.
Результаты
Ингибирование HDAC1 или miR-124-5p увеличивает вес крыс с депрессией
Крыс взвешивали за 1 день до моделирования CUMS (день 0), через 1 день после прекращения моделирования (день 36) и через 7 дней после вмешательства лентивируса (день 52). Перед моделированием не было обнаружено различий в массе тела крыс в каждой группе (все P > 0,05). После моделирования вес крысы снизился в группе CUMS, группе sh-NC, группе sh-HDAC1, группе анти-miR-NC и группе анти-miR-124-5p по сравнению с нормальной группой (все P <0,05). Не было замечено никаких расхождений в весе крыс в группе CUMS, группе sh-NC, группе sh-HDAC1, группе анти-miR-NC и группе анти-miR-124-5p (все P > 0,05). После вмешательства вес крысы снизился в группе CUMS по сравнению с нормальной группой (все P <0,05). Не было замечено расхождений в весе крыс в группе CUMS, группе sh-NC и группе анти-miR-NC (все P > 0,05). Вес крысы увеличился в группе sh-HDAC1 и группе анти-miR-124-5p по сравнению с их группами NC (обе P <0,05), что указывает на то, что ингибирование HDAC1 или miR-124-5p увеличивает вес крыс с депрессией (рис. 2а).
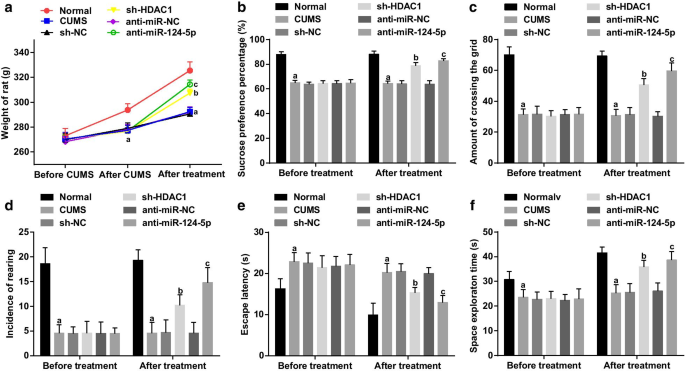
Ингибирование HDAC1 или miR-124-5p увеличивает вес и улучшает способности к обучению и памяти крыс с депрессией. а Влияние ингибирования HDAC1 или miR-124-5p на массу тела крыс с депрессией; б SPP до и после ингибирования HDAC1 или miR-124-5p; c Частота пересечения пояса в OFT до и после ингибирования HDAC1 или miR-124-5p; г Частота выращивания при OFT до и после ингибирования HDAC1 или miR-124-5p; е Латентность выхода в тесте MWM до и после ингибирования с помощью HDAC1 или miR-124-5p; е Время исследования космоса в тесте MWM до и после ингибирования HDAC1 или miR-124-5p; нет =10; а P <0,05 по сравнению с нормальной группой; б P <0,05 по сравнению с группой sh-NC; c P <0,05 по сравнению с группой анти-miR-NC. Сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа, а попарные сравнения - с помощью апостериорного теста Тьюки
Ингибирование ЛИБО HDAC1 или miR-124-5p улучшает способности к обучению и памяти у депрессивных крыс
Тесты SPT, OFT и MWM применялись для определения SPP, частоты пересечения сетки и частоты выращивания, а также задержки выхода и времени исследования космоса. Было отмечено, что (рис. 2b – f) до вмешательства, по сравнению с нормальной группой, SPP, частота пересечения сетки, частота выращивания и время освоения космоса уменьшились, тогда как латентность побега увеличилась в группе CUMS, sh-NC группа, группа sh-HDAC1, группа анти-miR-NC и группа анти-miR-124-5p (все P <0,05), что свидетельствует о развитии депрессивноподобного поведения у крыс. Не было расхождений в SPP, частоте пересечения сетки, частоте выращивания, времени освоения космоса и латентности побега среди группы CUMS, группы sh-NC, группы sh-HDAC1, группы anti-miR-NC и группы anti-miR-124. -5p группа (все P > 0,05). После интерференции, по сравнению с группой sh-NC и группой анти-miR-NC, SPP, частота пересечения сетки, частота вскармливания и время освоения космоса увеличивались, в то время как латентность побега сокращалась в группе sh-HDAC1 и группе анти-miR-124-. Группа 5p (все P <0,05). Никаких различий не наблюдалось в SPP, частоте пересечения сетки, частоте выращивания, времени освоения космоса и латентности побега среди группы CUMS, группы sh-NC и группы anti-miR-NC (все P > 0,05), предполагая, что подавление HDAC1 или подавление miR-124-5p может ослабить депрессивно-подобное поведение и улучшить способности к обучению и памяти у крыс.
Ингибирование HDAC1 или miR-124-5p ослабляет патологическое повреждение нейронов у крыс с депрессией
Наблюдение за поражениями гиппокампа с помощью окрашивания HE (рис. 3a) показало, что аккуратно расположенные нейроны в гиппокампе крыс в нормальной группе имели четкую морфологию, нормальную структуру, плотные слои, светлые ядра клеток и очевидные ядрышки. Нейроны крыс в группе CUMS были сморщены, уменьшились в количестве и рыхло расположены с неравномерно распределенным хроматином и истонченным слоем. Крысы в группах sh-NC и анти-miR-NC показали ту же ситуацию, что и группа CUMS. Крысы в группах sh-HDAC1 и anti-miR-124-5p демонстрировали увеличенный порядок нейронов и ослабленное повреждение по сравнению с их группами NC. Было указано, что нокдаун HDAC1 или ингибирование miR-124-5p облегчает поражение гиппокампа у крыс с депрессией.
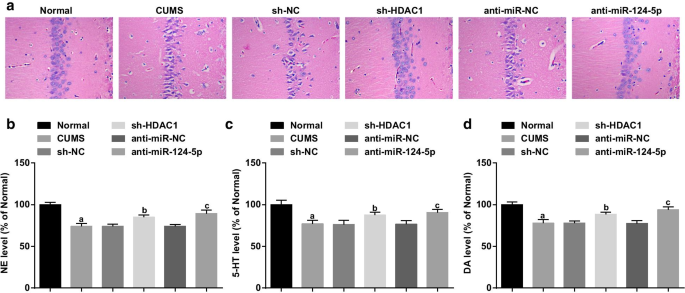
Ингибирование HDAC1 или miR-124-5p ослабляет патологическое повреждение гиппокампа и повышает экспрессию нейротрансмиттеров у крыс с депрессией. а Окрашивание HE поражений гиппокампа депрессивных крыс; б ELISA экспрессии NE в тканях гиппокампа; c ELISA экспрессии 5-HT в тканях гиппокампа; г ELISA экспрессии DA в тканях гиппокампа; нет =6; а P <0,05 по сравнению с нормальной группой; б P <0,05 по сравнению с группой sh-NC; c P <0,05 по сравнению с группой анти-miR-NC. Сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа, а попарные сравнения - с помощью апостериорного теста Тьюки
Ингибирование HDAC1 или miR-124-5p активирует экспрессию нейротрансмиттера у крыс с депрессией
Депрессия была связана с нарушениями нейротрансмиттеров головного мозга. Поэтому уровни нейротрансмиттеров DA, NE и 5-HT в гиппокампе крысы измеряли с помощью ELISA. Результаты показали, что (рис. 3b – d) с нормальной группой, напротив, пониженные уровни DA, NE и 5-HT были обнаружены у крыс из группы CUMS, группы sh-NC, группы sh-HDAC1, анти-miR. -NC-группа и анти-miR-124-5p группа (все P <0,05). Не было обнаружено различий в экспрессии нейромедиатора в группе CUMS, группе sh-NC и группе анти-miR-NC (все P > 0,05). По сравнению с группами sh-NC и анти-miR-NC уровни DA, NE и 5-HT увеличивались у крыс из групп sh-HDAC1 и anti-miR-124-5p (все P <0,05), что означает, что подавление HDAC1 или уменьшение miR-124-5p может повышать уровни DA, NE и 5-HT в гиппокампе крыс с депрессией.
Ингибирование HDAC1 или miR-124-5p подавляет окислительный стресс и воспаление у крыс с депрессией
Измеряли экспрессию связанных с окислительным стрессом и воспалительных факторов в сыворотке крови. Что касается нормальной группы, активность SOD и GSH была нарушена, в то время как уровни MDA, IL-1β, TNF-α и NO были увеличены в других модельных группах депрессии (все P <0,05). Активности SOD и GSH, а также уровни MDA, IL-1β, TNF-α и NO в группе CUMS, группе sh-NC и группе анти-miR-NC не показали различий (все P > 0,05). В отношении группы sh-NC и группы анти-miR-NC было замечено увеличение активности SOD и GSH, а также снижение уровней MDA, IL-1β, TNF-α и NO в sh- Группа HDAC1 и группа анти-miR-124-5p (все P <0,05) (рис. 4a – f), что указывает на то, что истощение HDAC1 или подавление miR-124-5p может облегчить окислительный стресс и воспаление у крыс с депрессией.
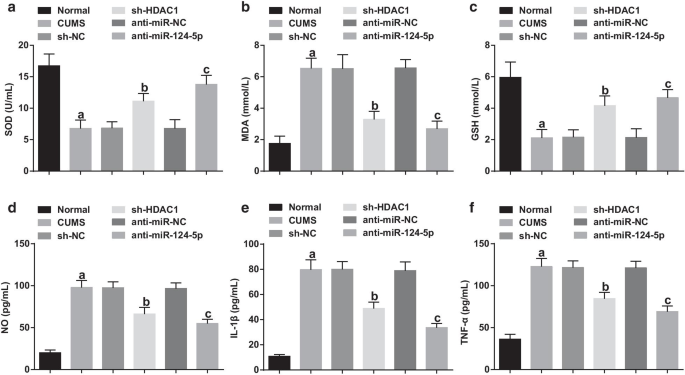
Ингибирование HDAC1 или miR-124-5p подавляет окислительный стресс и воспаление у крыс с депрессией. а - c Эффект ингибирования HDAC1 или miR-124-5p на концентрацию SOD, MDA и GSH в сыворотке крыс с депрессией; г - е Эффект ингибирования HDAC1 или miR-124-5p на концентрацию IL-1β, TNF-α и NO в сыворотке крыс с депрессией; нет =10; а P <0,05 по сравнению с нормальной группой; б P <0,05 по сравнению с группой sh-NC; c P <0,05 по сравнению с группой анти-miR-NC. Сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа, а попарные сравнения - с помощью апостериорного теста Тьюки
HDAC1 является посредником miR-124-5p для регулирования NPY
RT-qPCR и вестерн-блот-анализ были приняты для обнаружения HDAC1, miR-124-5p и NPY в гиппокампе. Было отмечено, что (рис. 5a, b) по сравнению с нормальной группой, HDAC1 и miR-124-5p увеличились, а NPY снизились в группе CUMS (все P <0,05). Экспрессия HDAC1, miR-124-5p и NPY не показала изменений в группе CUMS и группе sh-NC (все P > 0,05). По сравнению с группой sh-NC, группа sh-HDAC1 отражалась снижением HDAC1 и miR-124-5p и повышенными уровнями экспрессии NPY (все P <0,05). Приведенные выше данные свидетельствуют об успешном вмешательстве лентивируса и положительной связи между экспрессией miR-124-5p и HDAC1.
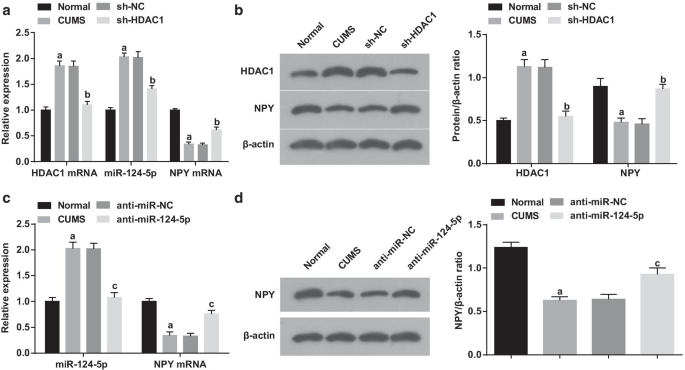
HDAC1 опосредует miR-124-5p для регуляции NPY. а Экспрессия мРНК HDAC1, miR-124-5p и NPY в тканях гиппокампа после ингибирования HDAC1 с помощью RT-qPCR ( n =6); б Экспрессия белков HDAC1 и NPY в тканях гиппокампа после ингибирования HDAC1 с помощью вестерн-блоттинга ( n =6); c Экспрессия мРНК MiR-124-5p и NPY в тканях гиппокампа после ингибирования miR-124-5p с помощью RT-qPCR ( n =6); г Экспрессия белка NPY в тканях гиппокампа после ингибирования miR-124-5p с помощью вестерн-блоттинга ( n =6); а P <0,05 по сравнению с нормальной группой; c P <0,05 по сравнению с группой sh-NC. Сравнения между несколькими группами оценивались с помощью однофакторного дисперсионного анализа, а попарные сравнения - с помощью апостериорного теста Тьюки
Кроме того, экспрессия miR-124-5p и NPY после подавления miR-124-5p была обнаружена с результатами (рис. 5c, d), демонстрирующими, что по сравнению с нормальной группой, повышенные miR-124-5p и уменьшенные NPY были представлены в группа CUMS (обе P <0,05). Напротив, группа анти-miR-124-5p имела тенденцию к уменьшению miR-124-5p и повышению NPY по сравнению с группой анти-miR-NC (обе P <0,05). Не было обнаружено расхождений в экспрессии miR-124-5p и NPY в группе CUMS и группе анти-miR-NC (обе P > 0,05). Результаты свидетельствуют об успешном вмешательстве лентивируса и отрицательной связи между NPY и miR-124-5p.
Анализ ChIP должен был проверить, может ли HDAC1 связываться с промотором miR-124-5p, и результаты показали, что (рис. 6a, b) HDAD1 был связан только с промотором miR-124-5p ( P <0,001), что указывает на то, что HDAC1 может напрямую регулировать miR-124-5p.
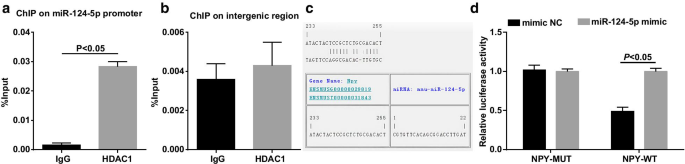
HDAC1 связывается с промотором miR-124-5p. а Изобилие связывания промоторной области HDAC1 и miR-124-5p в клетках HEK293T с помощью анализа ChIP ( n =3); б Изобилие связывания HDAC1 и неродственных межгенных областей в клетках HEK293T с помощью анализа ChIP ( n =3); c Сайт связывания MiR-124-5p и NPY на веб-сайте программного обеспечения биологической информации; г Нацеленная связь между miR-124-5p и NPY с помощью двойного анализа репортерного гена люциферазы ( n =3); Сравнения между двумя группами были проверены t тест
Связь нацеливания между miR-124-5p и NPY была предсказана и подтверждена с помощью инструмента RNA22 и двойного анализа репортерного гена люциферазы (рис. 6c, d). With respect to the cells co-transfection with NPY-3′UTR-WT and mimic NC, the cells with co-transfection of NPY-3′UTR-WT and miR-124-5p mimic showed impaired luciferase activity (P <0,05). No difference was recognized in the luciferase activity in the cells co-transfected with NPY-3′UTR-MUT and mimic NC, and the cells co-transfected with NPY-3′UTR-MUT and miR-124-5p mimic (P > 0.05).
Knockdown of NPY Abolishes the Protective Effects of Inhibited miR-124-5p on Depressed Rats
Spontaneous depletion of NPY and miR-124-5p was programmed to explore their interplay in depressed rats. It was exhibited that (Fig. 7a, b) lower NPY expression level was noticed in the anti-miR-124-5p + sh-NPY group versus the anti-miR-124-5p + sh-NC group (P <0,05). Body weight and behavioral function tests illustrated that (Fig. 7c–f) by comparison with the anti-miR-124-5p + sh-NC group, the rats in the anti-miR-124-5p + sh-NPY group presented reduced weight, SPP, frequency of crossing the grid, incidence of rearing and space exploration time with increased escape latency (all P <0,05). HE staining of the hippocampal lesions pictured that (Fig. 8a) in comparison with the anti-miR-124-5p + sh-NC group, the number of hippocampal neurons was reduced, and hippocampal neurons were darkly stained, sparsely and disorderly arranged in an irregular shape with reduced cell layers in the anti-miR-124-5p + sh-NPY group. Moreover, versus the anti-miR-124-5p + sh-NC group, the rats in the anti-miR-124-5p + sh-NPY group was accompanied by reduced DA, NE and 5-HT (all P < 0.05) (Fig. 8b). Besides, impaired SOD and GSH activities, and increased MDA, IL-1β, TNF-α and NO levels existed in the rats of the anti-miR-124-5p + sh-NPY group versus the anti-miR-124-5p + sh-NC group (all P < 0.05) (Fig. 8c, d). Collectively, knockdown of NPY abolished the protective effects of repressed miR-124-5p on depressed rat.

Knockdown of NPY abolishes the effects of inhibited miR-124-5p on body weight and behavioral function of depressed rats. а MiR-124-5p and NPY mRNA expression in hippocampal tissues after inhibition of both miR-124-5p and NPY by RT-qPCR (n = 6); б NPY protein expression in hippocampal tissues after inhibition of both miR-124-5p and NPY by western blot analysis (n = 6); c Body weight of depressed rats after inhibition of both miR-124-5p and NPY (n = 10); г SPP of depressed rats after inhibition of both miR-124-5p and NPY (n = 10); е Frequency of crossing the grid and incidence of rearing after inhibition of both miR-124-5p and NPY (n = 10); е Escape latency and space exploration time after inhibition of both miR-124-5p and NPY (n = 10); Comparisons between two groups were tested by t тест

Knockdown of NPY abolishes the effects of inhibited miR-124-5p on hippocampal damages of depressed rats. а HE staining of hippocampal lesions of rats after inhibition of both miR-124-5p and NPY (n = 6); б ELISA of NE, 5-HT and DA expression in hippocampal tissues of rats after inhibition of both miR-124-5p and NPY (n = 6); C. Effect of inhibited NPY and suppressed miR-124-5p on SOD, MDA and GSH concentration in serum (n = 10); D. Effect of inhibited NPY and suppressed miR-124-5p on IL-1β, TNF-α and NO concentration in serum (n =10). Comparisons between two groups were tested by t тест
Discussion
Depression is a type of psychiatric disorder comprising a variety of conditions with diverse symptoms [20]. Though emerging studies have implied the role of miRNAs in depression, the precise action of miR-124-5p has been rarely investigated. Hence, the present study is projected for better comprehension of the mechanism of miR-124-5p/HDAC1/NPY axis in depression with the major outcome elaborating that silencing of either HDAC1 or miR-124-5p up-regulated NPY to improve memory and learning abilities of depressed rats.
To begin with, this study discovered HDAC1 expression in hippocampal tissues with the findings suggesting the elevation in the HDAC1 expression. Subsequently, with the purpose to decipher the functional roles of HDAC1 in depressed rats, loss-of-function assays were performed and it was disclosed that knockdown of HDAC1 increased the body weight, improved learning and memory abilities, attenuated pathological damage, up-regulated neurotransmitter expression, and suppressed oxidative stress and inflammation in depressed rats. Currently, a study has implied an increment in the expression of HDAC1 in depressive-like and anxiety-like phenotypes resulted by stress-offspring [21]. Moreover, the expression of HDAC1 is documented to up-regulate in penumbra in photothrombotic stroke [22]. Besides that, the incremental HDAC1 mRNA expression is found in granule and pyramidal cells in temporal lobe epilepsy [23]. As to the functional role of HDAC1, it has been reported that virus-mediated overexpression of neuronal HDAC1 in the hippocampus of mice imposes influences on loss of contextual fear memory in particular [24]. Mechanically, HDAC inhibitors reverse cognitive deficits found in neurodegenerative diseases and age-related memory loss [25]. Actually, it is accepted that the HDAC1 suppression by tianeptinaline has advanced neuroplasticity and reinforced memory [26]. As mentioned in a prior study, it is concluded that repression of HDAC1 inhibits the pathogenic processes that lead to motor neuron degeneration in mitochondrial diseases [27]. Experimentally, the silencing of HDAC1 by 5-thienyl-substituted 2-aminobenzamide-type is partially involved in the prevention of neuronal cell death in Parkinson's disease models [28]. Further supported by those researches, the protective effects of silenced HDAC1 have been witnessed in brain diseases, including but not limited to depression.
Then, our study discovered a targeting relationship between HDAC1 and miR-124-5p, which was supported by a prior study which suggests miR-124 transcription is in the charge of EVI1, acting by connection with the deacetylase HDAC1 [29]. miR-124-5p was the overexpressed gene in depressed rats and knockdown of miR-124-5p had the similar functions of silenced HDAC1 in depressed rats. In fact, there is a study indicating that the expression of miR-124 in the hippocampus is up-regulated from 5 to 6 weeks in depression-like behavior phenotypes [7]. Another study has identified the increase in the miR-124 expression in female with cocaine use disorder [30]. Also, it is previously described that miR-124-3p is highly expressed in stressed rodents in major depressive disorder [31]. Regarding to the effects, knockdown of miR-124 in the prefrontal cortex is reported to attenuate depression-like behavior of mice [9]. Besides that, miR-124 knockdown is believed to serve as an antidepressant agent of chronic corticosterone-induced gypenosides in mice [32]. Drawn from a prior study, knockdown of miR-124 can result in improved behavioral susceptibility to a milder stress paradigm [33]. It is reported that suppression of miR-124 by lentivirus transfection in the hippocampus can protect ketamine-induced neurodegeneration in vivo and in vitro [34]. Anyway, miR-124-5p suppression is an active actor to attenuate depression.
Furthermore, NPY expression was verified to be regulated by HDAC1 and miR-124-5p. The deteriorated deficits associated with HDAC2 in histone acetylation may be related to the decreased expression of NPY and can used to control anxiety-like and drinking behaviors [14]. NPY was down-regulated in depressed rats and up-regulation of NPY promoted learning and memory ability recovery in depressed rats. In the development of depressive-like behaviors, the rat models are manifested with reduced NPY expression [17]. Academically, the expression of NPY is evidenced to decline in mice with depression [35]. Consistently, the above-mentioned study findings are as same as the previous literature to some extent.
The novel findings of study suggested that inhibited miR-124-5p or suppressed HDAC1 attenuated learning and memory abilities and increased body weight of depressed rats. In addition, knockdown of miR-124-5p or inhibition of HDAC1 suppressed oxidative stress and inflammation and promoted neurotransmitter expression of depressed rats. Moreover, HDAC1 mediated miR-124-5p to regulate NPY. In the rescue experiments, knockdown of NPY abolished the protective effects of inhibited miR-124-5p on depressed rats.
Conclusion
In summary, this study highlighted the effect of HDAC1/miR-124-5p/NPY axis in depression with the major findings suggesting inhibited miR-124-5p or suppressed HDAC1 attenuated learning and memory abilities, increased body weight, suppressed oxidative stress and inflammation, as well as promoted neurotransmitter expression in depressed rats. HDAC1/miR-124-5p/NPY axis may provide a reference to treat neurological disorder, which may also update the existed knowledge of depression. However, further studies are still required for thorough comprehension of the complex mechanism of HDAC1/miR-124-5p/NPY axis in depression.
Доступность данных и материалов
Не применимо.
Сокращения
- miR-124-5p:
-
MicroRNA-124-5p
- HDAC1:
-
Histone deacetylase 1
- NPY:
-
Neuropeptide Y
- CUMS:
-
Chronic unpredictable mild stress
- SD:
-
Sprague–Dawley
- SPF:
-
Specific pathogen-free
- NC:
-
Negative control
- OFT:
-
Open field test
- SPT:
-
Sucrose preference test
- SPP:
-
Sucrose preference percentage
- MWM:
-
Morris water maze test
- SOD:
-
Superoxide dismutase
- MDA:
-
Malondialchehyche
- GSH:
-
Глутатион
- IL-β:
-
Interleukin-β
- TNF-α:
-
Tumor necrosis factor-α
- NO:
-
Nitric oxide
- ELISA:
-
Иммуноферментный анализ
- HE:
-
Hematoxylin–eosin
- BCA:
-
Bicinchoninic acid
- NE:
-
Norepinephrine
- 5-HT:
-
Serotonin
- DA:
-
Дофамин
- RT-qPCR:
-
Reverse transcription quantitative polymerase chain reaction
- cDNA:
-
Complementary DNA
- SDS:
-
Додецилсульфат натрия
- ChIP:
-
Chromatin immunoprecipitation
- UTR:
-
Untranslated region
- MUT:
-
Mutant type
- WT:
-
Wild type
- ANOVA:
-
One-way analysis of variance
Наноматериалы
- Создание эффективных приложений Интернета вещей с помощью tinyML и автоматизированного машинного обучения
- ST:8-битные микроконтроллеры с богатым аналогом и DMA в недорогом пакете SO-8
- Настойчивость и настойчивость в руководстве
- Обучение без учителя с искусственными нейронами
- Повышение надежности и улучшение результатов обслуживания с помощью машинного обучения
- Умный наноматериал и нанокомпозит с продвинутой агрохимической активностью
- Модель удержания TaO / HfO x и TaO / AlO x RRAM с характеристиками самовосстанавливающегося переключателя
- Простой синтез окрашенного и проводящего композита CuSCN, покрытого наночастицами CuS
- Совместно модифицированные ТИМы RGO и трехмерных графеновых сетей с высокой производительностью
- Механизм проводимости и повышение выносливости в RRAM на основе HfO2 с лечением нитридом