


Анализ организации актина и фокальной адгезии в клетках U2OS на полимерных наноструктурах
Аннотация
Фон
В этой работе мы исследуем, как на ячейки U2OS влияют массивы полимерных наностолбиков, изготовленных на плоских стеклянных поверхностях. Мы сосредоточены на описании изменений в организации актинового цитоскелета, а также в расположении, количестве и форме фокальных спаек. Из наших результатов мы определяем, что клетки можно разделить на разные режимы в зависимости от их поведения при распространении и адгезии на наностолбиках. Количественный анализ предполагает, что клетки, засеянные на плотные массивы наностолбиков, подвешены на вершине столбов с фокальными адгезиями, формирующимися ближе к периферии клетки по сравнению с плоскими поверхностями или разреженными массивами столбов. Это изменение аналогично аналогичным ответам для клеток, засеянных на мягкие субстраты.
Результаты
В этой работе мы исследуем, как на ячейки U2OS влияют массивы полимерных наностолбиков, изготовленных на плоских стеклянных поверхностях. Мы сосредоточены на описании изменений в организации актинового цитоскелета, а также в расположении, количестве и форме фокальных спаек. Из наших результатов мы определяем, что клетки можно разделить на разные режимы в зависимости от их поведения при распространении и адгезии на наностолбиках. Количественный анализ предполагает, что клетки, засеянные на плотные массивы наностолбиков, подвешены на вершине столбов с фокальными адгезиями, формирующимися ближе к периферии клетки по сравнению с плоскими поверхностями или разреженными массивами столбов. Это изменение аналогично аналогичным ответам для клеток, засеянных на мягкие субстраты.
Заключение
В целом, мы показываем, что сочетание высокопроизводительного нанофабриката, передовой оптической микроскопии, инструментов молекулярной биологии для визуализации клеточных процессов и анализа данных может быть использовано для исследования того, как клетки взаимодействуют с наноструктурированными поверхностями, и в будущем поможет создать культуральные субстраты, которые вызывают определенные функция ячейки.
Абстрактное графическое изображение
Фон
In vivo клетки обычно находятся в сложной трехмерной среде, называемой внеклеточным матриксом (ЕСМ). ECM не только служит структурным каркасом для клеток, но также транспортером биомеханических и биохимических сигналов и, таким образом, регулирует ряд процессов, таких как морфогенез ткани, гомеостатис и дифференцировка. Он состоит из воды, полисахаридов и белков [1,2,3,4], а состав варьируется в зависимости от типа ткани.
Мотивированные необходимостью создания моделей культивирования клеток, которые лучше отражают условия in vivo, исследователи все чаще начинают изучать поведение клеток также в трехмерных матрицах и в «полу-трехмерных» системах. Выявлен ряд различий в клеточных фенотипах между плоскими субстратами и системами с более высокой размерностью [5, 6]. Например, известно, что такие характеристики, как жизнеспособность, пролиферация, дифференцировка и морфология, различаются между клетками на плоских поверхностях и клетками, встроенными в трехмерные матрицы [3, 7].
Подложки, подобные in vivo, варьируются от субстратов «полу-3D» / 2,5D, таких как плоские поверхности, украшенные различными наноструктурами, до систем «истинно-3D», таких как коллагеновые гели или матрицы матригелей [8,9,10,11]. Кроме того, контролируемое расположение лигандов на поверхности может дать новое понимание того, как клетки взаимодействуют с различными химическими паттернами [12,13,14]. Также было показано, что механические факторы, такие как жесткость структуры или даже химический состав поверхности, влияют на клеточную функцию [15,16,17]. С этой целью было разработано большое количество различных субстратов для клеточных исследований [3,18,19,20,21,22].
Также было высказано предположение, что системы трехмерного культивирования могут более точно предсказывать эффект лекарства in vivo, и, таким образом, эти системы могут найти применение в открытии лекарств [16, 23, 24]. Точное управление наноразмерными топографическими паттернами также можно использовать для регулирования морфологии клеток. Например, морщины и бороздки можно использовать для воссоздания полосатого расположения кардиомиоцитов и, таким образом, лучше представить физиологически релевантные условия для моделирования различных заболеваний [25, 26].
Цитоскелет клетки связан с ECM, чему способствуют фокальные адгезии (FA), мультибелковый комплекс, включающий интегрины клеточной поверхности и белки каркаса. В зависимости от сложного набора регуляторных механизмов, FAs формируются и разбираются со скоростью оборота, необходимой для поступательного движения, например, при миграции клеток. Известно, что ЖК оказывают механическое усилие на ЕСМ, и наоборот, сила воздействия на клетки ЕСМ, как известно, влияет на сродство к интегрину и авидность в мембране [27].
Одним из белков, которые, как известно, являются неотъемлемой частью ЖК, является винкулин. Это один из линкерных белков, участвующих в прикреплении F-актина к интегриновому комплексу. Недостаток винкулина изменяет морфологию, адгезию и подвижность клеток [28] и снижает способность клеток передавать силу субстрату [29,30,31]. Винкулин не только участвует в механическом соединении актинового цитоскелета с комплексами интегрина, он также обладает способностью связывать и связывать актиновые филаменты [32,33,34], изменять существующие актиновые связки [35], кэпировать актиновые филаменты, зарождаются новые сайты полимеризации актина [36] и привлекают модификаторы актина [37].
Клетки реагируют на 3D-матрицы, изменяя количество и тип адгезии клетка-субстрат и вызывая изменения в пространственной организации цитоскелета. Эти изменения, в свою очередь, влияют на распределение, размер и динамику образовавшихся спаек [4, 38,39,40,41]. Эта перестройка может привести к изменению пролиферации, морфологии и подвижности клеток [42].
Чтобы понять влияние сложной трехмерной среды на клетки, необходимо разработать новые модельные системы, в которых клеточные процессы можно было бы изучать и сравнивать с плоскими элементами управления. Поскольку известно, что клеточный ответ зависит от физических, механических и химических характеристик субстрата для культивирования, желательно изготавливать клеточные субстраты с точно контролируемыми свойствами [43,44,45]. Кроме того, очень выгодно, чтобы клетки и субстрат можно было легко изучить с помощью уже установленных методов анализа, таких как оптическая микроскопия.
Одним из типов подложек, которые недавно привлекли внимание, являются плоские поверхности, украшенные наностолбиками или нанопроволокой [18, 21, 46,47,48,49,50,51,52,53]. По сравнению, например, с гидрогелями, эти структурированные поверхности не имитируют истинную трехмерную среду, но имеют четко определенную топографию поверхности. Эти подложки обычно называют 2.5D. Такие системы уже применялись для облегчения доставки биологически релевантных молекул в клетки [54, 55], для мониторинга ферментативной активности [56], для проверки ядерной механики [57] и для изучения того, как настройка кривизны мембраны влияет на различные клеточные мембраны. процессы [58,59,60]. Создавая наноструктуры на прозрачных подложках, можно интегрировать этот подход с оптической микроскопией.
Число возможных комбинаций различных клеточных линий, типа и геометрии наноструктуры велико, а примеров из литературы предостаточно. Ли и др. описали поведение клеток на поверхностях, украшенных случайно расположенными наноструктурами фосфида галлия, и количественно оценили долю клеток с большими ЖК [61]. Морфология клеток и ЖК исследовалась на поверхности нанопроволок с различной плотностью поверхности. Результаты показали, что клетки, засеянные на поверхности с низкой плотностью, контактировали с субстратом и образовывали большие FA вокруг краев клеток. Большие FA были обнаружены в большой части ячеек на этих массивах. Для высоких поверхностных плотностей нанопроволок и ячейки были подвешены на вершине массивов нанопроволок, и наблюдались точечные FA под ячейками. Более низкая доля ячеек на этих массивах показала большие FA по сравнению с ячейками на поверхности с низкой плотностью поверхности нанопроволоки.
Buch-Månson et al. исследовали взаимодействие ячеек с наноструктурированной поверхностью для массивов кремниевых наноколонок, случайным образом расположенных на подложке Si [62]. В используемом процессе изготовления контролировалась поверхностная плотность, но не расстояние между столбами. Исследование ЖК показало, что клетки на массивах с промежуточной поверхностной плотностью имели наибольшее количество ЖК, которые также имели наиболее асимметричную форму. Было высказано предположение, что часть этих ТВС образовывалась на боковых стенках наноколонн. Этого не наблюдалось для поверхностей с низкой и высокой поверхностной плотностью наноколонок.
В предыдущей работе мы описали подробные протоколы изготовления полимерных наноструктур SU-8 на плоских стеклянных поверхностях [63] и исследовали поведение клеток для двух различных клеточных линий на этих поверхностях [45, 48]. В этой работе мы используем электронно-лучевую литографию (EBL) для изготовления поверхностей, украшенных вертикально ориентированными полимерными структурами SU-8, для изучения изменений в актиновой цитоскелетной организации и организации FA в линии эпителиальных клеток остеосаркомы U2OS. Мы проводим как качественный, так и количественный анализ изменений, вызванных поверхностью с различными топологическими ориентирами.
Результаты
Используя ранее установленные протоколы, мы изготовили стеклянные покровные стекла, украшенные точно определенными массивами вертикально ориентированных наностолбиков (НП) SU-8 с переменным разделением и определенной геометрией [63]. Поверхности с поверхностной плотностью НЧ 456, 205, 115 и 29 НЧ / 100 мкм 2 (соответствующие шагам 500, 750, 1000 и 2000 нм). Сначала мы исследуем общие тенденции в морфологии клеток, структуре актинового цитоскелета и взаимодействиях клетки с субстратом. Мы следим за этим путем количественного сравнения морфологии клеток и ЖК на различных наноструктурированных подложках и контроле из плоского стекла. Мы сочетаем микроскопию высокого разрешения с производством с высокой производительностью, чтобы провести качественный анализ не менее \ ({\ приблизительно 100} \) клеток для каждого типа поверхности, с получением изображений через 24 и 48 часов. Всего мы анализируем> 400 изображений с высоким разрешением и 20 наборов 3D-данных.
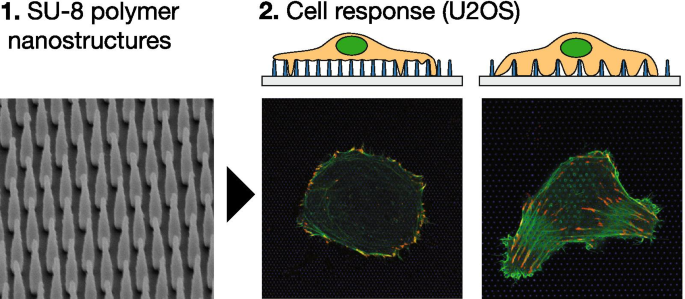
На рисунке 7 показано схематическое изображение массивов наночастиц (A, B) и электронно-микроскопические изображения изготовленных подложек (C, D). Стеклянные предметные стекла, содержащие структуры, изготовленные из наночастиц, монтировали с использованием парафина под полым дном, 35-миллиметровую культуру структурами, направленными вверх. Электронно-микроскопические изображения сверху вниз и названные сбоку, представленные на рис. 1C, D, показывают массивы наностолбиков с шагом и высотой 1000 нм. В таблице 1 приведены геометрические параметры массивов, используемых в данной работе, их классификация, а также соответствующая им плотность площади НП. Мы классифицируем массивы NP на плотные и разреженные в зависимости от наблюдаемого поведения клеточной адгезии (см. Ниже).
На рисунке 1 показаны типичные клетки U2OS, культивируемые на стекле (A) и наноструктурированной поверхности (B – F) в течение 24 часов. Клетки были котрансфицированы pCMV-LifeAct-GFP и pTAG-RFP-винкулин, что позволяет визуализировать F-актин и винкулин посредством продукции флуоресцентных слитых белков LifeAct-TagGFP2 (далее:LifeActGFP) и TagRFP-винкулин соответственно. Сеть F-актина и области, богатые винкулином, в FA четко выявляются. На периферии клетки сигнал LifeActGFP находится в непосредственной близости от мембраны, и поэтому мы используем этот сигнал для визуализации морфологии клетки. Сигнал от НП СУ-8 показан синим цветом (см. Раздел «Экспериментальная часть»).
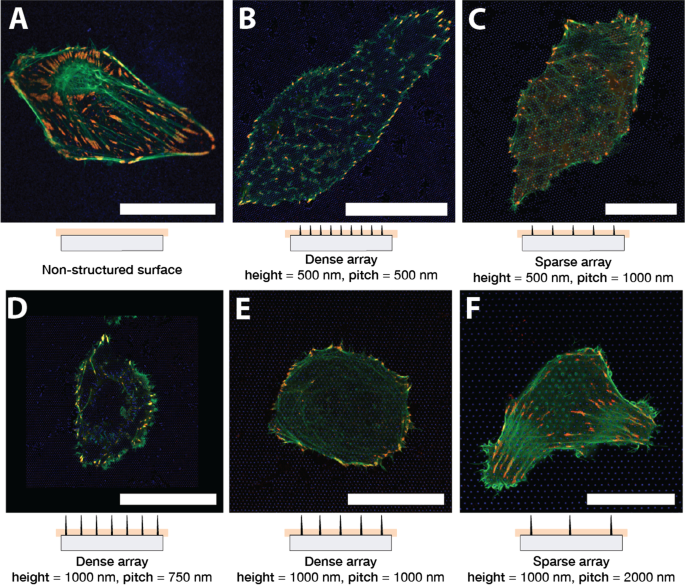
U2OS экспрессирует флуоресцентный LifeActGFP (зеленый) и TagRFP-винкулин (красный) на разных типах поверхности. Желтая окраска указывает на перекрывающиеся сигналы от каналов LifeActGFP и TagRFP-винкулин. Под каждой микрофотографией показан схематический вид сбоку соответствующего массива NP вместе с приблизительным положением плоскости сбора данных. Изображения клеток на A неструктурированная плоская стеклянная поверхность и на массивах столбов с разным шагом решетки и высотой конструкции: B Шаг 500 нм и высота 500 нм, C Шаг 1000 нм и высота 500 нм, D Шаг 750 нм и высота 1000 нм, E Шаг 1000 нм и высота 1000 нм, и F Шаг 2000 нм и высота 1000 нм. Все представленные изображения представляют собой репрезентативные клетки. Шкала 25 мкм. Обратите внимание, что C имеет другой масштаб, чем другие изображения
Предварительные тесты с использованием трансфицированных клеток показали, что клетки, засеянные как на стеклянной, так и на структурированной поверхности, по-видимому, полностью распространились примерно через 6 часов. Никаких четких различий в распределении клеток между разными поверхностями не наблюдалось, и визуальный осмотр не выявил признаков снижения жизнеспособности клеток из-за массивов столбиков на этом этапе или в более поздних экспериментах. В следующих экспериментах клетки трансфицировали через 6 часов после посева, а затем визуализировали через 24 часа, 48 часов после трансфекции, что соответствует трансфекции через 30 часов, 54 часа после посева. В дальнейшем эти два момента времени будут обозначаться временем наблюдения после посева, то есть 24 часа 48 часов.
Клетки высевали на массивы NP высотой 500 нм, 1000 нм и шагом 750 нм, 1000 нм, 2000 нм. После начального распространения клетки были либо круглыми, либо удлиненными, как и на плоских поверхностях. Было обнаружено, что эта общая морфология согласована в нескольких экспериментах. Клетки, засеянные на разреженные массивы NP, обычно имели форму, подобную клеткам на стеклянной поверхности, см. Фиг. 1F, на которой изображена типичная ячейка на решетке с шагом 2000 нм. Волокна F-актина присутствовали также в основании НЧ и вблизи поверхности стекла, что указывает на то, что клетки могли получить доступ к области, близкой к субстрату. Как наблюдалось ранее [45, 62, 64], клетки на плотных массивах обычно кажутся подвешенными на вершине NP (рис. 1B, D, E). Клетки в плотных массивах, по-видимому, имеют менее заметный F-актин вблизи поверхности стекла, что указывает на то, что актиновые волокна не образовывались между столбами в непосредственной близости от субстрата. Связь между высотой наночастиц и разделением определяли, прилипали ли клетки к субстрату или были подвешены на вершине массива наночастиц. Это, например, проиллюстрировано на рис. 1C, где более короткие наночастицы приводят к контакту клетки с субстратом, тогда как более длинные наночастицы препятствуют контакту, рис. 1E. Эти наблюдения актиновых волокон были дополнительно подтверждены z-стеками, выполненными для некоторых поверхностей, как показано на рис. 2.
Чтобы получить более подробное представление о том, как клетки прикрепляются к структурированной и неструктурированной поверхности, мы оценили распределение ЖК, визуализированное по присутствию слитого белка TagRFP-винкулин. Клетки на плоских поверхностях обычно образуют удлиненные ЖК, распределенные под всем телом клетки, как показано на рис. 1А.
На разреженных массивах U2OS могли контактировать с поверхностью стекла между НЧ и прикрепляться аналогично клеткам на стекле, как показано на рис. 1C, F. Для этих массивов НЧ ЖК образовывались на стекле между НЧ, и сигнал F-актина также был обнаружен на изображении, полученном вблизи основания НЧ. Это указывало на то, что клетки могли изгибать мембрану вокруг наноструктур. Однако клетки на более плотных массивах, таких как 750 нм, разделение 1000 нм и высота 1000 нм, явно препятствовали прилипанию к подложке между наноструктурами, как показано на изображениях рис. 1D, E, которые были получены вблизи основания НП. Однако по периферии клетки, как правило, могли прикрепляться к субстрату между наноструктурами, формирующими ЖК, что часто определялось симметрией нижележащего массива столбов.
Клетки, распространяющиеся на массивы NP с меньшей длиной и с расстоянием между столбами 1000 нм, образовывали адгезии как к периферии, так и под телом клетки. Ориентация волокон F-актина была направлена симметрией нижележащего массива, как показано на фиг. 1C. Однако расположение и ориентация винкулинсодержащих ЖК не демонстрирует какой-либо четкой картины, причем ЖК формируются между НЧ.
Клетки U2OS на столбах 500 нм с расстоянием между столбами 500 нм обычно образовывали меньшее количество и меньшие адгезии по сравнению с плоской поверхностью, как показано на рис. 1B. Для клеток, засеянных на этот массив, актиновые волокна в основном наблюдались вблизи поверхности стекла в положениях, где они заканчивались в FA. Опять же, это признак того, что актиновая сеть не могла контактировать с поверхностью, и поэтому предполагалось, что клетки подвешены на вершине массива. Однако клетки, по-видимому, имеют более неповрежденную сеть F-актина, формирующуюся над столбами. Части актиновой сети, которые наблюдались между столбами, по-видимому, выравниваются со структурами в нижележащем массиве NP. Это можно увидеть на рис. 1В, поскольку волокна и ЖК F-актина преимущественно формируются вдоль одного из направлений решетки, то есть параллельно открытым «линиям» массива столбов.
И на плотных, и на разреженных массивах мы наблюдали «кольцеобразные» структуры F-actin, которые формировались вокруг NP, которые выступают вверх в тело клетки. Кольцевая структура F-актина оказалась более заметной на разреженных массивах, как показано на рис. 1E, F.
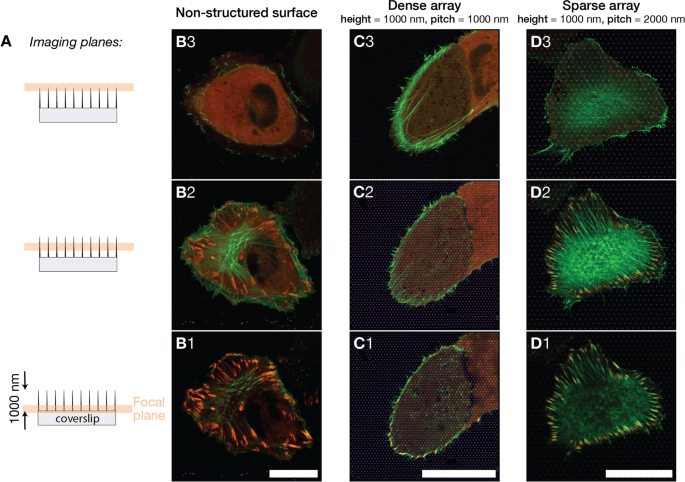
А Изображения клеток U2OS на разных поверхностях были получены в трех указанных фокальных плоскостях. Изображения были получены близко к основанию столба, примерно на половине высоты столба и близко к вершине столба. Плоскости изображения располагались на расстоянии 0,0 мкм, 0,4 мкм, 0,8 мкм от плоской поверхности стекла. Б - D Объединенные флуоресцентные изображения, показывающие, что U2OS экспрессирует флуоресцентный LifeActGFP (зеленый) и TagRFP-винкулин (красный) на разных типах поверхности, желтый цвет указывает на перекрытие LifeActGFP и TagRFP-винкулин. Изображенные поверхности были B бесструктурная стеклянная поверхность, C Решетка с наклонными столбами 1000 нм, D Решетка с наклонной стойкой 2000 нм. Представленные изображения являются репрезентативными для клеток на каждом типе поверхности. Вертикальное расстояние между фокальными плоскостями для каждого типа поверхности составляет примерно 400 нм. Масштабные линейки 25 мкм
На основании результатов, представленных выше, мы выбрали три поверхности для более подробного и количественного описания морфологии клеток и ЖК. Мы изучаем ячейки на плотных массивах (шаг 1000 нм, длина 1000 нм), разреженных массивах (шаг 2000 нм, длина 1000 нм) и сравниваем результаты с ячейками на плоских стеклянных поверхностях, используемых в качестве контроля.
Используя детектор Airyscan вместе со специальной постобработкой изображений, мы смогли получить изображение с разрешением по оси xy около 140 нм и разрешением по оси z около 400 нм [65]. На рисунке 2 показаны изображения клеток на трех поверхностях с разделением плоскостей изображения приблизительно на 400 нм. Исследование пучков F-актина при разных z . Клетки на плоской поверхности имели четко видимую сеть F-актина в той же фокальной плоскости или прямо над FAs (см. Рис. 2-B2). Для клеток на плотных массивах сеть F-актина была обнаружена в более высокой фокальной плоскости внутри клетки по сравнению с плоскостью FA, которая находилась в контакте со стеклянной подложкой (Fig. 2-C1 и C2 / C3). Для клеток на разреженных массивах ситуация была аналогична клеткам на стеклянном контроле и актиновой сети, и FA были обнаружены на той же высоте (рис. 2-D2). Эти данные подтверждают первоначальное наблюдение, что клетки в разреженных массивах прикрепляли поверхность между структурами, тогда как клетки в плотных массивах в основном могли прилипать к поверхности вокруг периферии клеток.
Для анализа и количественной оценки различий в FA и морфологии клеток для трех выбранных поверхностей использовался сценарий анализа изображений на основе Python (см. «Экспериментальная часть»). Для количественного анализа было проанализировано более 300 изображений с высоким разрешением. На этих изображениях было идентифицировано> 400 клеток и> 7700 FA, в таблице 2 указано количество обнаруженных клеток и FA для трех типов поверхности, включенных в анализ. Для всех поверхностей клетки визуализировали как через 24, так и через 48 часов после трансфекции. В следующем анализе сравниваются геометрические параметры, такие как площадь поверхности, округлость и соотношение сторон для ячеек на 3 типах поверхностей и через 24 часа, 48 часов. Дополнительный анализ можно найти в дополнительной информации. Площадь поверхности, округлость и соотношение сторон для ячеек показаны на Рис. 3, а для количества FA, объединенной площади FA на ячейку и доли площади FA на ячейку на Рис. 4. Геометрические параметры были определены, как описано в «Экспериментальной части». раздел.
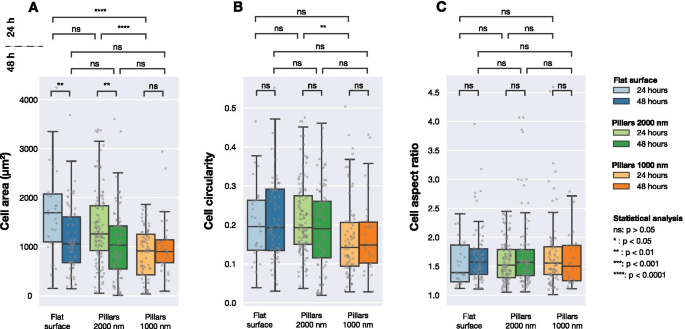
Расчетная площадь клеток, округлость клеток и соотношение сторон клеток для клеток U2OS, экспрессирующих флуоресцентные слитые белки LifeActGFP и TagRFP-винкулин на различных поверхностях, отображаемых через 24 и 48 часов после трансфекции. Обнаружение изображения выполнялось на основе интенсивности сигнала LifeActGFP. Каждая серая точка соответствует одной ячейке, а на прямоугольных диаграммах показаны медианные значения ( Q 2), а также первый ( Q 1) и третий квартиль ( Q 3). Статистические различия между распределениями оценивались с помощью непараметрического критерия Манна – Уитни
На рисунке 3 представлены данные, собранные как для плоских, так и для структурированных поверхностей. Как показано на фиг. 3A, значительные различия в площади клеток наблюдались после 24 часов культивирования клеток. Однако через 48 ч значимой разницы между средней площадью клеток на исследуемых поверхностях не было. При рассмотрении округлости клеток (рис. 3B) не было обнаружено значительных различий между различными поверхностями, за исключением клеток, засеянных на плотных и редких столбах, отображаемых через 24 часа. Ячейки на всех трех поверхностях имели одинаковое среднее соотношение сторон, как показано на рис. 3C.
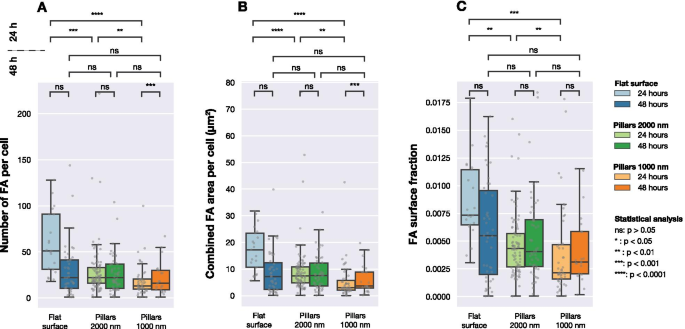
Количество FA, объединенная площадь FA на ячейку и доля площади FA в площади ячейки для клеток, отображаемых на трех различных типах поверхности (плоские и столбчатые массивы с шагом 1000 нм и 2000 нм). Каждая серая точка соответствует наблюдению из одной ячейки. Статистическая значимость между распределениями определялась с помощью непараметрического критерия Манна – Уитни
На рисунке 4 показано распределение количества обнаруженных FA на ячейку, общую площадь поверхности FA в каждой ячейке и отношение площади FA к площади ячейки. Через 24 часа количество ЖК, образованных клетками на трех разных поверхностях, значительно различается. Как показано на фиг. 4B, общая площадь поверхности FA на ячейку была разной для ячеек, засеянных на плоских и структурированных поверхностях. То же самое можно увидеть при сравнении относительного количества FA (общая площадь обнаруженных FA, деленная на общую площадь ячеек) для разных поверхностей, как показано на рис. 4C.
Однако через 48 ч культивирования значительных различий между популяциями клеток больше не наблюдалось. При рассмотрении количества ЖК на ячейку, объединенной площади ЖК на ячейку или доли поверхности ЖК не было обнаружено различий между тремя поверхностями.
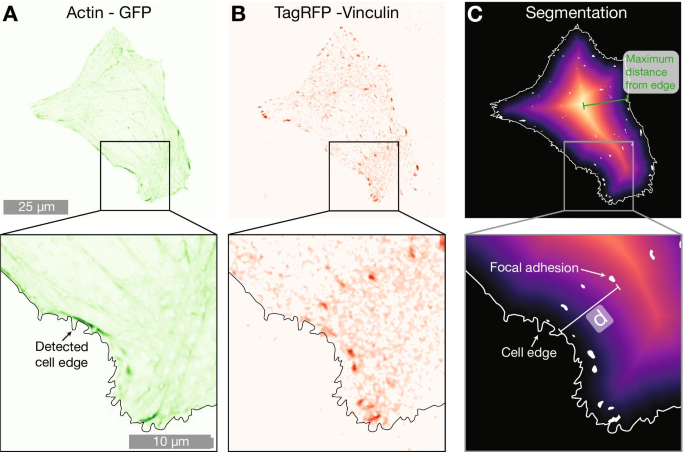
Пример ячейки на массиве столбов 2000 нм, выражающий A LifeActGFP (зеленый) и B TagRFP-винкулин (красный). На обоих рисунках наложен край обнаруженной ячейки, определенный с использованием сигнала от выражения LifeActGFP (как показано на A ). В Показывает карту расстояний от края обнаруженной ячейки, а также обнаруженных FA. Кратчайшее расстояние (обозначается d на рисунке) от каждого сегментированного винкулинового пятна (белые области) до периферии клетки (обозначены сплошной белой линией) был рассчитан для всех FA на изображениях
Чтобы понять, влияет ли присутствие NP на локализацию FA в клетке, мы провели дальнейший анализ, используя расположение FA. Данные микроскопии показали, что ЖК в клетках на плотных массивах НЧ располагались ближе к периферии клетки, как показано на рис. 1 и 2. Чтобы количественно оценить эту тенденцию, мы рассчитали кратчайшее расстояние от каждой FA до края ячейки. Это было выполнено, как проиллюстрировано на фиг. 5. F-актин был использован для определения местоположения периферии, и путем построения карт расстояний было вычислено расстояние между каждым центром детектированной FA и периферией клетки. Чтобы учесть различия в размерах ячеек, мы нормализовали расстояния между обнаруженным краем ячейки и FA на максимальное расстояние от края до геометрического центра для каждой ячейки (расстояние, эквивалентное радиусу для ячеек, имеющих круглую форму). Данные, нормированные на максимальное расстояние, представлены на рис. 6 и в таблице 3. Для ячеек на плоских поверхностях FA распределены больше к центру ячейки, тогда как как на разреженных массивах 2000 нм, так и на плотных массивах 1000 нм, FAs расположены ближе к периферии клетки. Этот эффект наиболее заметен для плотных массивов с шагом 1000 нм. Результаты альтернативного подхода к нормализации, в котором положения FA были нормализованы по площади поверхности клетки, включены в дополнительную информацию. Эти данные показывают те же качественные тенденции, что и данные, нормированные на максимальное расстояние до края в каждой ячейке, представленные на рис. 6.
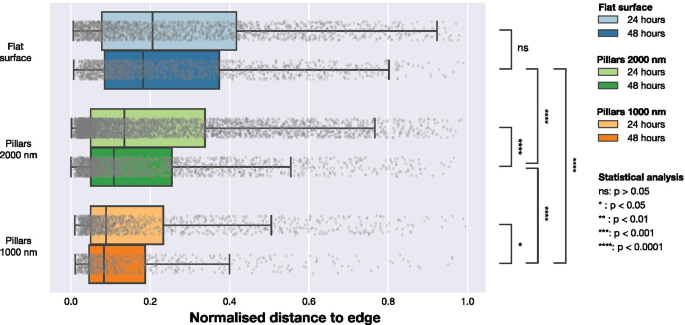
Распределение положений ТВС относительно ближайшего края ячейки, нормированное на максимальное расстояние от геометрического центра ячейки до края. Расстояния были получены путем расчета расстояния от каждой наблюдаемой FA до края клетки, определяемого сигналом LifeActGFP, и нанесены на график для трех типов поверхности через 24 часа и 48 часов. Серые точки представляют отдельные наблюдения за FA, а их распределения суммированы на прямоугольных диаграммах. Чтобы проверить вероятность того, что FA с разных поверхностей и временных точек были из одного распределения, тесты Манна-Уитни были выполнены для всех распределений с уровнями значимости, обозначенными на рисунке
Обсуждение
Организация актинового цитоскелета и образование спаек - это процессы, которые широко изучаются на плоских поверхностях. Настоящее исследование было разработано для изучения и количественной оценки изменений в организации актинового цитоскелета и фокальных адгезий на массивах наностолбиков. В этом исследовании мы не исследовали, как массивы наностолбиков влияют на миграцию клеток. Однако мы ожидаем, что клетки в плотных массивах могут иметь более высокую подвижность, как это наблюдалось ранее на тех же столбцах для эмбриональных фибробластов мыши [45].
Позволяя клеткам распространяться и прикрепляться к поверхности в течение длительного периода времени, можно наблюдать организацию актинового цитоскелета и присутствие полностью созревших ЖК. Через 24 часа после посева мы наблюдаем значительные различия в площади ячеек, округлости и соотношении сторон для ячеек, засеянных на поверхности. Однако через 48 часов не было обнаружено значительных различий, что указывает на то, что клетки через 24 часа еще не полностью прилипли к поверхности и что наноструктуры в основном влияют на организацию ЖК до того, как они полностью созреют. Ни через 24 часа, ни через 48 часов не наблюдается образования ЖК сверху или по бокам НЧ.
Изменения в организации актинового цитоскелета также связаны с тем, как клетка взаимодействует с окружающей средой. Например, и стресс-волокна, и ЖК растут при растяжении и, по-видимому, функционально взаимозависимы [66]. Другие сообщили, что более округлые клетки и локализация FA по краям часто наблюдаются для клеток, засеянных на мягких или податливых поверхностях [67]. В наших результатах мы наблюдаем аналогичную тенденцию. Ячейки в плотных массивах имеют тенденцию иметь волокна по краю ячейки, как показано на рис. 1E или 2B. Кажется, что FAs формируются близко к краю ячейки для этих ячеек. Мы предполагаем, что, когда у клеток отсутствует плоская поверхность, например, когда клетки подвешены на вершине столбов, распределение ЖК напоминает ЖК на мягких субстратах, как это использовали Прагер-Хоуторски и его сотрудники [67].>
Из наших результатов мы наблюдаем, что клетки, подвешенные на вершине столбов, действительно имеют развитую актиновую сеть над столбами, но без образования ЖК на самих столбах. Однако для клеток в разреженных массивах клетки оказываются менее подверженными влиянию НЧ, и как актиновая сеть, так и ЖК выглядят более «плоскими».
Взаимодействие между FAs и актиновым цитоскелетом сложное и еще не полностью охарактеризовано. FAs, связывающие актиновый цитоскелет с ECM, как известно, действуют как точки притяжения и способствуют образованию стрессовых волокон в клетках. Напротив, волокна актина снова влияют на организацию и созревание ЖК. Многочисленные исследования описывают, как клетки обычно подвешиваются на поверхности плотных массивов NP [61, 62, 68] и как клеточные мембраны взаимодействуют с одиночными NP [69,70,71]. Эти наблюдения подтверждаются теоретическими исследованиями [64], и поведение клеток на столбах достаточно хорошо изучено.
Механизм образования и прикрепления ЖК к субстрату вокруг края клетки на плотных массивах остается неясным. В этом отношении особенно интересно сравнение с клетками на мягкой подложке. For soft substrates, actin fibres are organised in a ring like fashion close to the cell edge and FAs form around the cell periphery [67]. On the nanopillar arrays similar type of architecture is observed, but the actin fibres are typically shorter. Similar qualitative trends in terms of actin organisation and FA formation were observed by Li et al. for cells seeded on random nanowire arrays made from gallium phosphide [61].
In our studies we also observed formation of F-actin rings around NPs. The formation of F-actin rings around NP has previously been described for fibroblasts on similar surfaces [45] and for U2OS cells on nanostructures with a range of structure sizes [58].
Contrasting our results to other studies highlight an important aspect of studies on cellular response to NP arrays:cellular response may vary considerably depending on cell type, NP material, NP geometry and as well as other parameters. For example, Buch-Månson et al. studied fibroblasts and investigations of FAs showed that cells suspended on arrays with intermediate NP density had the highest number of FAs. In our results we do not see a similar trend. However, these studies cannot be directly compared as Buch-Månson et al. studied another cell line using a system with different array geometry, surface porosity and NPs length [62].
There are also studies describing the effect FAs placement has on cells [41]. By modelling cells on planar substrates Stolarska et al. suggest that the cells can control intra-cellular stresses by three mechanisms:FA position, FA size and attachment strength. FAs around the periphery allows the cells to be more sensitive to changes in the micro-environment. This could also be an underlying mechanisms for cells on NPs. Yet, it is not obvious that the results for the planar substrate are directly transferable to NP decorated surfaces.
Cell-interactions with the surrounding environment, for flat substrate, NPs arrays or in vivo ECM, are regulated by a complex set of relations between actin organisation, membrane mechanics, cell dynamics and contact with FAs. To further explore these relations, applying flat surfaces structured with NPs could be one promising approach. Such surfaces may also aid in exploring discrepancies in the cellular response to environmental cues between different cell lines.
Conclusions
In order to create more physiologically relevant systems for cellular studies, a plethora of 3D and 2.5D approaches have been proposed. One approach is to use flat-surfaces decorated with vertically aligned nanostructures as a simple model system. High resolution live cell imaging of co-transfected U2OS cells expressing pCMV-LifeAct-GFP and pTAGRFP-Vinculin have been used to study the influence of nanopillar arrays on actin cytoskeleton focal adhesion organisation. Our present results indicate that the U2OS cells spreading on surfaces decorated with nanopillars can be categorised into three different regimes by how they respond to the nano-structures. These observed changes are quantified by analysing more than 400 high-resolution images, and indicate that tuning geometrical properties of the nanostructured surface can be used to direct cell behaviour.
More specifically, the U2OS cells were found to either contact the substrate, attach preferably around the cell edge, or be fully suspended on top of the vertical NP arrays. In the latter case, we hypothesise that the resulting reorganisation of FA and cytoskeleton is an effect analogous to what is seen for softer substrates.
Increased understanding of how cells behave on nano-structured surfaces, such as pillar arrays, could help us discover more details about complex cellular processes. For example, it is still poorly understood how changes in the actin cytoskeleton and its architecture influence cell signalling. By studying the cell response on nanostructured surfaces in a systematic way, the potential connection between actin cytoskeleton, cell adhesions and a plethora of biochemical signalling pathways could be further explored. We therefore envision that further development of the presented platform and analysis could have implications for advanced in vitro applications or for development of smarter in vivo biointerfaces.
Methods
Fabrication of Nanostructures and Sample Mounting
SU-8 nanostructures were fabricated as previously explained [63]. Briefly, 24 mm by 24 mm glass cover slips (#1.5, Menzel-Gläser, thickness 170 μm) were cleaned by immersion in acetone, isopropyl alcohol, rinsed in de-ionised water and dried. The cover slips were then oxygen plasma treated for 2 min (Diener Femto plasma cleaner, power 100 W, base pressure 0.3 torr), followed by dehydration for 10 min on a 150 °C hot plate. Samples were then placed in a desiccator containing an open vial of Hexamethyldisilazane (HMDS, Sigma Aldrich product no:440191). HMDS was applied by vapour deposition, the desiccator was pumped to low vacuum using a diaphragm pump for 5 min and the samples were kept in HMDS atmosphere for 60 min.
Substrates for EBL were prepared directly after HMDS treatment by spin coating SU-8 2001 (Microchem Corp.) to a desired thickness of 500 nm and 1000 nm. SU-8 was made fluorescent by adding either Oxazine 170 perchlorate, Rhodamine 800 or Coumarin 102 (all Sigma Aldrich) to a final concentration of 100 μg mL −1 resist. After spin coating samples were dehydrated on a hot plate at 95 °C. To mitigate charging during EBL exposure samples were then covered by a layer of conductive polymer AR-PC 5091 Electra 92 (AllResist GmbH) by spin coating at 2000 rpm for 60 s to thickness of 50 nm.
An Elionix ELS-G100 100 kV EBL-system was used to fabricate SU-8 nanopillars (NPs) with processing parameters as described in our previous work [63]. Table 1 summarise the arrays fabricated for this work. Pillar arrays were exposed using the Elionix dot-pattern generator where each pillar is exposed in a single exposure. Arrays were exposed over an area of 2000 μm by 4000 μm, with a current of 500 pA in write fields of 500 μm by 500 μm. NPs had a tip diameter of about 100 nm as a base diameter of 150 nm and 200 nm for structures of length 500 nm and 1000 nm respectively.
After EBL exposure, the samples were rinsed in DI-water to remove the conductive polymer, then post exposure baked for 2.3 min at 95 °C and developed twice in mr-Dev 600 (Micro Resist Technology GmbH) developer for 20 s, rinsed in isopropyl alcohol and dried. Samples were then treated with oxygen plasma (Diener Femto plasma cleaner, power 50 W, base pressure 0.3 torr) for 30 s to render SU-8 hydrophilic and to give it similar surface chemistry as glass by oxidising surface epoxy-groups to hydroxyl.
Fabricated structures were imaged using Scanning electron microscopy (SEM) and samples sputter coated with 5 nm Platinum/Palladium alloy deposited with a 208 HR B sputter coater (Cressington Scientific Instruments UK). SEM was performed with a FEI Apreo SEM, at 5 kV and 0.2 nA with sample 45° pre-titled stage and with additional tilting of 30°.

A Side view schematic representation of nano-structured surface mounted in petri dish. Glass slides are mounted using paraffin such that structures are pointing upwards. B Tilted schematic representation of nano-pillar array on flat surface, and two important parameters for the nano-pillar arrays (height and pitch). These figures are not drawn to scale. C , D Overview of the nanopillar arrays employed in this work. Top-down and tilted side-view scanning electron micrographs of fabricated nano-pillar array with pillars of height 1000 nm and pitch 1000 nm. Scalebars 2000 nm
When exposing the pillars, an indexing system was also exposed to make navigation during live-cell imaging more reliable. Arrays were optically inspected after fabrication to ensure free and standing pillars. The short Oxygen plasma treatment to render the SU-8 structures did not lead to any optically visible change to the structures. Lastly, the samples were mounted underneath 35 mm diameter dishes (Cellvis, Mountain View, CA, USA) with 14 mm holes and nano-structures pointing upwards, as indicated schematically in Fig. 7. As flat surfaces, areas outside the structured part of the same samples were used. Before usage, all dishes were disinfected with 70% ethanol twice and dried.
Cell Culture and Transfection
U2OS-cells (ATCC) were cultivated in Dulbecco’s modified Eagle’s Medium (DMEM Prod. 41965039, Fischer Scientific) with 10% fetal bovine serum (FBS) and kept at 5% CO2 and 37 °C. Before detachment, cells were washed with PBS and detached with Trypsin-ethylenediaminetetraacetic acid (trypsin-EDTA) and seeded on nanostrucutred or flat surfaces. For the diameter 14 mm glass wells 15,000 cells were seeded.
For the standard transfection experiments, cells were allowed 6 h for adhering to surfaces before transfection. U2OS cells were transiently transfected using Lipofectamine 2000 (Invitrogen, Fischer Scientific) by adapting the manufacturer protocol to our system. Briefly, 2 μL Lipofectamine 2000 was added to 50 μL Opti-MEM I Reduced Serum Media (Prod. 11058021, Gibco , Fischer Scientific) and incubated for 5 min at room temperature. Plasmid DNA coding for fluorescent LifeAct-TagGFP2 and TagRFP-vinculin fusion proteins were co-transfected by using 0.5 μg plasmid DNA (vinculin-pTagRFP and pCMVLifeAct plasmids) was diluted in 50 μL Opti-MEM I and incubated at room temperature for 5 min. For co-transfection of TagRFP-vinculin and pCMVLifeAct 0.5 μg of each plasmid was used.
The diluted DNA was added to the diluted Lipofectamine 2000 in a 1:1 ratio, and left to incubate for 20 min at room temperature. 40 μL of the combined transfection complex was then added to each well. After 18 h, 1.5 mL DMEM (Prod. 41965039) supplemented with 10% FBS and 1% 10000U/mL Penicillin-Streptomycin was added to each dish.
For reverse transfection experiments, the same amounts of reactants were used, but the transfection complex was added to a suspension of U2OS cells, and the suspension was then added to the wells.
Microscopy
Live cell imaging was performed usin g a Zeiss LSM 800 Airyscan with an inverted Axio Observer Z1 stand connect to a PeCon compact incubator. Imaging was performed in an humidified environment at 37 °C, with 5% CO2 поток. High resolution imaging was performed using a Zeiss Plan-Apochromat 63x/1.4NA DIC M27 oil objective with Cargille Immersion Oil Type 37 (n =1.51) suited for use at 37 °C. All images were taken using the system optimised pixel size both in-plane (typically 34 nm) and for stacks in the vertical axis (typically 180 nm).
To minimise imaging bias, imaging was performed in a standardised manner where each pillar array was raster scanned and cells expressing both LifeActGFP and Vinculin RFP were imaged. The high resolution images were then processed using a Zeiss algorithm for reconstruction of AiryScan images and exported as CZI-files for further manual and automatised image processing.
Image Analysis
For all cells, cell shape was based on the expression LifeActGFP fusion protein and expression of TagRFP-vinculin was used to identify FAs. Segmentation of images was performed using a script written in Python 3 [72] using CZIfile [73] (version 2017.09.12) for reading the microscopy images in Zeiss-format. The python packages Scipy [74] and Scikit-image [75] were used for multi-dimensional image processing and image segmentation respectively.
To reduce the influence from fluorescence cross-talk from pillars (due to Oxazine 170 perchlorate, Rhodamine 800 or Coumarin 102), the pillar/surface channel was used as a background and subtracted from the TagRFP-vinculin imaging channel. A median filter (size:10 pixels) was applied to remove noise from the TagRFP-vinculin channel, followed by classification of the image into regions based on their intensity value using a Multi-Otsu approach. Multi-Otsu thresholding with three classes was applied. The first class was typically the background, the second class constituted the cytosolic vinculin, whereas vinculin rich areas in FAs appeared brighter and could be classified into a third class. The quality of the image segmentation was briefly assessed by comparison to manual segmentation.
Area of cells and vinculin rich regions were described by counting pixel numbers and from this the actual area was found by correcting for the pixel size. Shape geometries were described by fitting each region with an ellipse with the same second-moment as the segmented region. In order to describe the cell area geometry, three measures were used:(1) Aspect ratio defined as the ratio of the ellipse major axis to the minor axis. (2) Circularity given as,
$$\begin{aligned} C =\frac{4\pi *\text{Area}}{\text{Perimeter}^2}, \end{aligned}$$ (1)and roundness given as,
$$\begin{aligned} R =\frac{4*\text{Area}}{\pi *\text{MajorAxis}^2}. \end{aligned}$$ (2)Segmented vinculin areas with a fitted ellipse that were too round (aspect ratio \(\le 1.5\)) or too elongated (aspect ratio \(\ge 8.5\)) were rejected. In addition, vinculin areas smaller than 0.05 μm 2 were filtered out. In order to find the distance between each vinculin area and the cell edge, the shortest euclidean distance between each centroid (the centre of the fitted ellipse for each vinculin area) and the cell edge was calculated.
Statistical Analysis
Statistical comparisons of distributions were performed by using the non-parametric two-tailed Mann-Whitney test neither assuming normal distribution nor equal standard deviation. P -values \(\ge {0.05}\) were considered to represent a non-significant (ns) difference between the two populations. Significant values were denoted with * for p in 0.01 to 0.05, ** for p in 0.001 to 0.01, *** for p in 0.0001 to 0.001 and lastly **** for p \(\le {0.0001}\).
Abbreviations
- DMEM:
-
Dulbecco’s Modified Eagle’s Medium
- EBL:
-
Electron Beam Lithography
- ECM:
-
Extracellular Matrix
- EDTA:
-
Ethylenediaminetetraacetic Acid
- FA:
-
Focal Adhesion
- FBS:
-
Fetal Bovine Serum
- GFP:
-
Green Fluorescent Protein
- HMDS:
-
Hexamethyldisilazane
- NP:
-
Nanopillar
- RFP:
-
Red Fluorescent Protein
- SEM:
-
Scanning Electron Microscopy
Наноматериалы
- Примеры схем и списков соединений
- Батареи специального назначения
- Солнечный элемент
- Графен и полимерные композиты для суперконденсаторов:обзор
- Изучение in vitro влияния наночастиц Au на клеточные линии HT29 и SPEV
- Синтез нанокристаллов ZnO и применение в инвертированных полимерных солнечных элементах
- Изучение силы адгезии и стеклования тонких пленок полистирола с помощью атомно-силовой микроскопии
- Стимуляция роста клеток SH-SY5Y наночастицами золота, модифицированными 6-меркаптопурином и проникающим в нейро…
- Влияние упругой жесткости и поверхностной адгезии на отскок наночастиц
- Последние достижения в синтетических методах и применении серебряных наноструктур