


Новый механизм, лежащий в основе формирования боковых корней томатов, вызванных многостенными углеродными нанотрубками:участие оксида азота
Аннотация
Многочисленные исследования показали, что многослойные углеродные нанотрубки (MWCNT) токсичны для растений. Однако вопрос о том, влияют ли MWCNT на формирование боковых корней (LR), которые являются важным компонентом адаптируемости корневой системы к различным сигналам окружающей среды, и каким образом. В этом отчете мы обнаружили, что MWCNT могут проникать в корни проростков томатов. Введение MWCNT способствовало образованию LR томатов примерно дозозависимым образом. Производство эндогенного оксида азота (NO) запускалось MWCNT, что подтверждено методом реагентов Грейсса, электронным парамагнитным резонансом (ЭПР) и лазерной сканирующей конфокальной микроскопией (LSCM) вместе с поглотителем NO. Между MWCNT и NO существует причинно-следственная связь в индукции развития LR, поскольку запускаемый MWCNT синтез NO и образование LR, очевидно, блокировались удалением эндогенного NO с его поглотителем. Активность NO-генерирующего фермента нитратредуктазы (NR) увеличивалась в ответ на MWCNT. Вольфраматное ингибирование NR не только ухудшает продукцию NO, но также устраняет образование LR, запускаемое MWCNT. Добавление N G Метиловый эфир -нитро-1-аргинина (l-NAME), ингибитор фермента, подобного синтазе оксида азота (NOS) млекопитающих, не влиял на образование LR. В совокупности мы предположили, что NO может действовать как нижележащая сигнальная молекула в MWCNT, контролируя развитие LR, по крайней мере частично, через NR.
Введение
Углеродные нанотрубки имеют множество биологических и биомедицинских применений [1, 2]. Благодаря уникальной способности легко проникать через клеточные мембраны, биобезопасность углеродных нанотрубок всегда является предметом дискуссий [3, 4]. Между тем, поскольку производство и использование углеродных нанотрубок быстро растет, становится важным охарактеризовать подробные механизмы их цитотоксичности у людей и млекопитающих, а в последнее время и у растений [3,4,5,6,7,8,9]. Хорошо известно, что растения и их сообщества очень важны для человека и окружающей среды, и они также подвержены риску воздействия углеродных нанотрубок из-за их накопления в почве из-за внесения биотвердых удобрений [6, 10, 11]. Поскольку многослойные углеродные нанотрубки (MWCNT), состоящие из нескольких свернутых слоев графена, являются важными членами углеродных нанотрубок, они широко исследовались. Исследования на млекопитающих показали, что воздействие как MWCNT, так и одностенных углеродных нанотрубок вызывает окислительное повреждение и активацию NF-κB в кератиноцитах человека и клетках A549 [9, 12]. МУНТ и однослойные углеродные нанотрубки могут сливаться с плазматической мембраной, вызывая повреждение клеток из-за перекисного окисления липидов и окислительного стресса [9, 11, 13, 14]. Цитотоксичность и окислительный стресс, вызванные MWCNT, а также умеренные воспалительные реакции наблюдались в эндотелиальных клетках пупочной вены человека [15]. Предыдущее исследование показало, что основная токсичность MWCNT в красном шпинате в основном связана с избыточным продуцированием активных форм кислорода (АФК), и токсические эффекты могут быть устранены добавлением аскорбиновой кислоты [7]. В этом смысле MWCNT рассматриваются как новый стрессовый фактор для организмов, будь то животных или растений.
Формирование бокового корня (LR), важный детерминант корневой архитектуры, рассматривается как индикатор адаптивного ответа на различные стрессы [16]. У высших растений на формирование LR влияют фитогормоны и широкий спектр факторов окружающей среды, включая доступность воды, питательные вещества и абиотический стресс, такой как гипоксия и стресс от тяжелых металлов [17,18,19]. Между тем, имеется множество доказательств того, что образование LR не только действует как физическая поддержка, но также способствует поглощению воды и питательных веществ для роста и развития растений [19,20,21]. Различные факторы окружающей среды могут запускать несколько специфических фенотипов морфогенного ответа, вызванного стрессом (SIMR), включая стимулирование образования LR и ингибирование удлинения корня [17]. Регуляция образования LR также строго контролируется фитогормонами, такими как ауксин, предполагалась активация регуляторных генов клеточного цикла в ответ на ауксин [19, 22]. Между тем, участие некоторых небольших молекул в органогенезе корней, инициируемом ауксином, было подтверждено на растениях огурца, томата, сои и рапса [23,24,25,26,27]. Эти небольшие молекулы включают перекись водорода (H 2 О 2 ), оксид азота (NO), оксид углерода (CO) и газообразный водород (H 2 ).
Среди них NO, газ свободных радикалов, как было показано, выполняет множество физиологических функций в растениях [28, 29]. Помимо усиления адаптации растений к стрессам, функции NO включают стимулирование развития корневых волосков, придаточного укоренения и образования боковых корней [30,31,32,33], хотя ферментные ресурсы биосинтеза NO в них вышеупомянутые процессы остаются неуловимыми. У животных синтез NO из l-аргинина катализируется гемсодержащим ферментом синтазой оксида азота (NOS) [34]. Хотя ген (ы), кодирующий ферменты NOS, не был идентифицирован у растений, активность, подобная NOS у млекопитающих, выявляется широко [35, 36], а ингибиторы NOS млекопитающих, такие как N G гидрохлорид метилового эфира нитро-1-аргинина (l-NAME) может ингибировать образование NO в растениях [25, 33, 36,37,38,39]. Важно отметить, что многочисленные генетические данные показали, что NO может продуцироваться нитратредуктазой (NR), хорошо известным ферментом, ответственным за метаболизм азота в растениях [28]. Участие NR-опосредованной продукции NO в закрытии устьиц и акклиматизации к холоду было продемонстрировано генетически [37, 38]. Наше предыдущее исследование показало, что NR-зависимый синтез NO участвует в индуцированном ауксином опосредованном газообразным водородом образовании боковых корней [39].
До сих пор сообщалось о различных ответах в образовании, стимулировании или ингибировании LR у разных видов растений при добавлении наноматериалов, включая MWCNT [40,41,42,43], наночастицы золота (Au NP, [44]), оксид цинка. наночастицы (ZnO NP [45, 46];), наночастицы диоксида титана (TiO 2 NP [46];) и оксид графена (GO [47,48,49];) (Таблица 1), и ни одно исследование еще не предоставило окончательного доказательства роли NO в вышеуказанных ответах. В этом исследовании обнаружение эндогенного NO методом реактива Грейсса, лазерной сканирующей конфокальной микроскопии (LSCM) и анализом электронного парамагнитного резонанса (ЭПР) показали, что уровень NO был повышен в проростках томатов, обработанных MWCNT. После этого наблюдали образование LR. Мы дополнительно изучаем участие NO в образовании LR, запускаемом MWCNT, путем манипулирования эндогенными уровнями NO с помощью поглотителя NO и антагонистов, которые ингибируют NR и активность NOS, подобную млекопитающим. Дальнейший эксперимент показал, что NR-зависимый NO может быть, по крайней мере частично, существенным для образования LR в ответ на MWCNTs. Таким образом, эта работа открывает новое окно для понимания биологического воздействия наноматериалов на растения.
Материалы и методы
Химические вещества
Если не указано иное, все остальные химические вещества были получены от Sigma-Aldrich (Сент-Луис, Миссури, США). МУНТ, приобретенные у Sigma-Aldrich, были охарактеризованы, как описано ранее [50]. Внешний диаметр, внутренний диаметр и длина МУНТ составляли 6–12 нм, 2,5–5 нм и 1–9 мкм соответственно. После обработки ультразвуком полученную коллоидную суспензию гомогената стерилизовали и использовали.
Другие углеродные наночастицы были получены от Nanjing XFNANO Materials Tech Co., Ltd., включая однослойные углеродные нанотрубки (ОСУНТ, XFS22; чистота> 95%, диаметр 1-2 нм, длина 5-30 мкм, особая площадь поверхности> 1075 мкм. 2 / г), графен (XF001W; чистота ~ 99%, диаметр 0,5–5 мкм, толщина ~ 0,8 нм, однослойность ~ 80%, площадь поверхности по БЭТ 500 ~ 1000 м 2 /г; удельное электрическое сопротивление ≦ 0,30 Ом · см) и активированный уголь (AC, XFP06; чистота> 95%, размер частиц 5 ± 1 мкм, объем пор 1–1,2 см 3 / г, апертура 2,0–2,2 нм, площадь специальной поверхности ~ 1500–1700 м 2 / г).
Кроме того, нитропруссид натрия (SNP) использовался как соединение, высвобождающее NO [30,31,32,33]. Калиевая соль 2- (4-карбоксифенил) -4,4,5,5-тетраметилимидазолин-1-оксил-3-оксида (cPTIO) рассматривалась как поглотитель NO [51,52,53,54]. Вольфрамат (Tg; ингибитор NR [28, 33, 37, 55,56,57];) и N G Гидрохлорид метилового эфира нитро-L-аргинина (НАЗВАНИЕ; ингибитор NOS-подобного фермента млекопитающих [25, 33, 36,37,38,39];) также применялся. В этом исследовании концентрации вышеуказанных химикатов были определены в пилотных экспериментах, в результате которых наблюдались значительные отклики.
Растительный материал, условия роста и определение образования LR
Помидор ( Solanum lycopersicum L.) семена «Цзяншу 14» были любезно предоставлены сельскохозяйственным институтом Цзянсу, Нанкин, провинция Цзянсу, Китай. Отобранные семена одинакового размера проращивали в дистиллированной воде при 25 ± 1 ° C в темноте в течение 3 дней. Отобранные идентичные проростки с корешками 2–3 мм затем переносили в 6 мл обрабатывающих растворов, содержащих указанные концентрации MWCNT, 200 нМ 1-нафтилуксусной кислоты (NAA; хорошо известный ауксин), 0,1 мМ SNP, 0,2 мМ cPTIO, 20 мкМ вольфрамат (Tg), 0,2 мМ NAME и другие углеродные наночастицы, включая однослойные углеродные нанотрубки (ОСУНТ) с концентрацией 5 мг / мл, графен и активированный уголь (AC), по отдельности или в комбинации для указанных временных точек. Проростки выращивали в инкубаторе с освещением (25 ± 1 ° C) при интенсивности света 200 мкмоль м -2 . s −1 при 14/10 ч (свет / темнота) фотопериод.
После обработки были сделаны снимки, а затем определено количество и длина появившихся боковых корней (> 1 мм) на проросток с помощью программного обеспечения Image J (http://rsb.info.nih.gov/ij/) [39 , 58]. Как описано ранее, для последующего анализа использовались только сегменты, индуцируемые боковым корнем.
Визуализация распределения MWCNT с помощью просвечивающей электронной микроскопии
Распределение MWCNT в корнях проростков томатов охарактеризовали с помощью просвечивающей электронной микроскопии (ТЕМ; JEOL, JEM-200CX, Токио, Япония). Подготовка проб для ПЭМ-анализа проводилась в соответствии с предыдущим протоколом [59].
Визуализация эндогенного NO с помощью лазерного сканирующего конфокального микроскопа
Визуализацию NO проводили с использованием довольно специфичного флуоресцентного зонда NO 4-амино-5-метиламино-2 ', 7'-дифторфлуоресцеина диацетата (DAF-FM DA). После тщательной промывки зонда изображения получали с помощью конфокального микроскопа Zeiss LSM 710 (Carl Zeiss, Оберкохен, Германия, возбуждение при 488 нм, испускание при 500–530 нм для анализа NO). В нашем эксперименте случайным образом были отобраны 20 отдельных образцов, которые измерялись за один сеанс лечения. Фотографии демонстрируют идентичные результаты.
НЕТ содержания, определенного с помощью анализа реагента Грисса
Согласно методам, описанным ранее [50], содержание NO определяли с помощью анализа реагента Грисса. Важно отметить, что для того, чтобы избежать помех, вызванных концентрированным содержанием нитратов и нитритов в растениях, идентичные образцы, предварительно инкубированные в 200 мкМ cPTIO (поглотитель NO) в течение 30 минут, считались холостыми. После добавления реактива Грисса в течение 30 минут, поглощение регистрировали при 540 нм, и определяли содержание NO путем сравнения со стандартной кривой NaNO 2 .
Определение NO с помощью электронного парамагнитного резонанса (ЭПР)
Согласно нашим предыдущим методам [39, 55, 60], определение уровня NO проводилось с помощью электронного парамагнитного резонанса (ЭПР). Слой органического растворителя использовали для определения NO на спектрометре Bruker A300 (Bruker Instrument, Карлсруэ, Германия) при следующих условиях:комнатная температура; частота СВЧ 9,85 ГГц; мощность СВЧ 63,49 мВт; и частота модуляции 100,00 кГц.
Определение активности нитратредуктазы (NR)
Активность NR детектировали спектрофотометрически при 540 нм по предыдущей методике [57]. Полученный нитрит определяли спектрофотометрически при 540 нм путем добавления 1 мл 1% (мас. / Об.) Сульфениламида в 3 M HCl вместе с 1 мл 0,02% (об.) Н- (1-нафтил) этилендиамина.
Статистический анализ
Где указано, результаты были выражены как средние значения ± стандартная ошибка трех независимых экспериментов с тремя биологическими повторностями для каждого. Статистический анализ проводился с использованием программы SPSS Statistics 17.0. Для статистического анализа используется множественный тест Дункана ( p <0,05) было выбрано соответствующим образом.
Результаты
MWCNT не только входят в корневые ячейки, но также способствуют формированию LR
Формирование LR является одним из основных факторов, определяющих архитектуру корневой системы. Чтобы исследовать влияние MWCNT на образование LR, 3-дневные проростки томатов инкубировали с диапазоном концентраций MWCNT (0,05, 0,5, 5 и 50 мг / мл) в течение 3 дней. Применение 1-нафтилуксусной кислоты (НАК) рассматривалось как положительный контроль. В нашем эксперименте количество и длина LR определялись как два параметра формирования LR. Как показано на фиг. 1, по сравнению с контрольными образцами, экзогенные MWCNT значительно индуцировали образование LR примерно дозозависимым образом с максимальным эффектом при 5 и 50 мг / мл. Подобный индуцибельный ответ наблюдался при введении 200 нМ NAA. Учитывая стоимость MWCNT и индуцируемую реакцию при образовании LR, в следующих экспериментах применяли MWCNT 5 мг / мл.
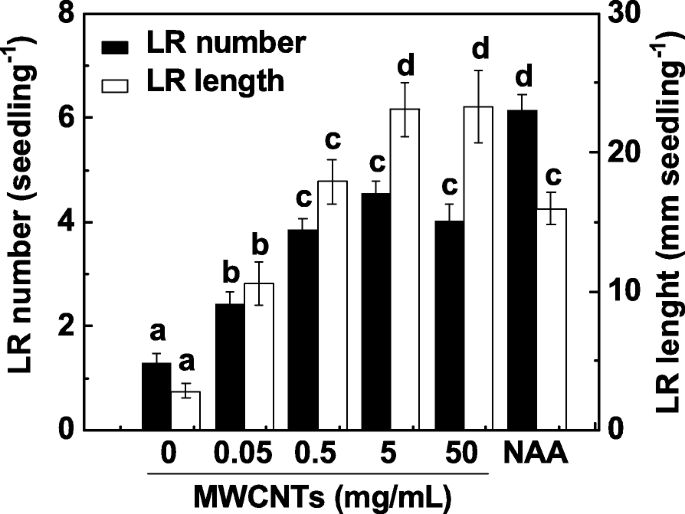
MWCNT-индуцированное образование LR томатов было примерно дозозависимым. Трехдневные проростки томатов обрабатывали 200 нМ NAA и указанными концентрациями MWCNT соответственно. Затем через 3 дня обработки определяли количество и длину появившихся боковых корней (> 1 мм) на проросток. Всего было 30 (10 × 3) растений в трех биологических повторах, эксперименты проводили 3 раза. Данные - это средние значения ± SE. В каждой серии экспериментов столбцы, обозначенные одной и той же буквой, существенно не различались при p Уровень <0,05 согласно множественному тесту Дункана
Чтобы подтвердить конкретную функцию MWCNTs в индукции образования LR, мы дополнительно исследуем, обладают ли другие аллотропии MWCNT также такими индуцибельными эффектами. Как показано на рис. 2а, все эти углеродные наноматериалы проявляют токсическое действие на рост побегов (данные не показаны). Интересно, что применение МУНТ, однослойных углеродных нанотрубок (ОСУНТ), графена и активированного угля с одинаковой концентрацией (5 мг / мл) может по-разному приводить к увеличению количества и длины LR по сравнению с контрольными установками, не содержащими химикатов. (Рис. 2б). Среди этих химикатов максимальный индуцибельный ответ был обнаружен у проростков томатов, инкубированных с MWCNT.
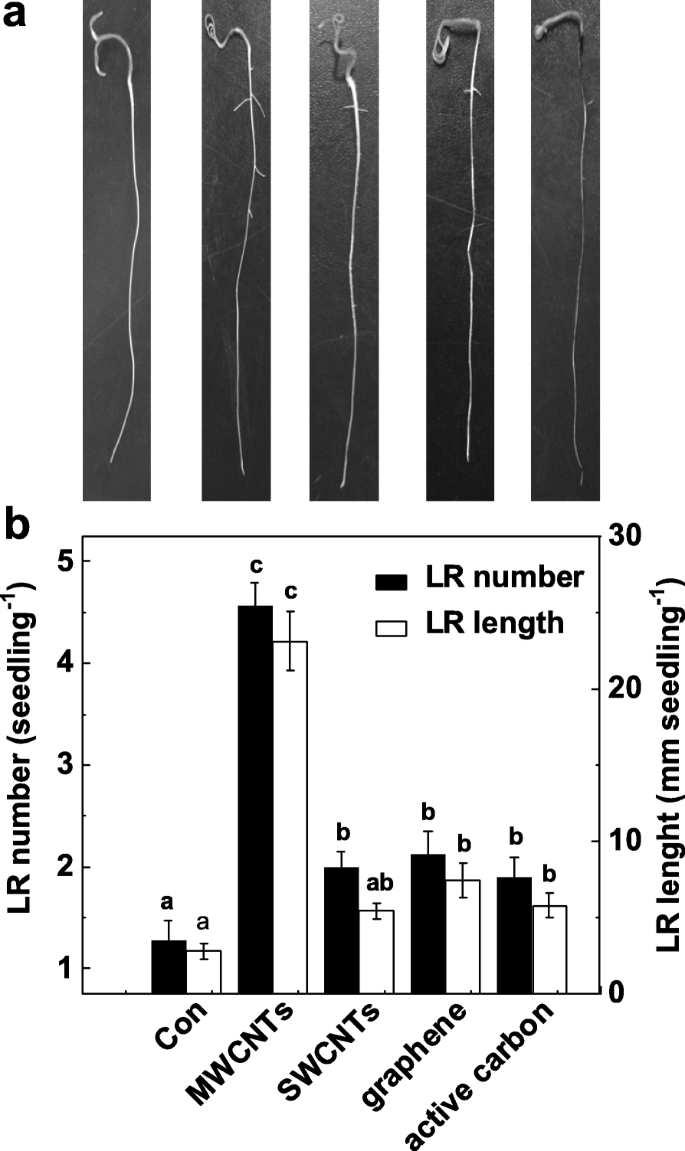
Изменения в формировании боковых корней в ответ на различные углеродные наночастицы. Трехдневные проростки томатов обрабатывали дистиллированной водой (Con), 5 мг / мл MWCNT, однослойными углеродными нанотрубками (SWCNT), графеном и активированным углем (AC) соответственно в течение еще 3 дней. а Затем были сделаны репрезентативные фотографии. б Затем определяли количество и длину появившихся боковых корней (> 1 мм) на проросток. Масштабная линейка =50 мм. Всего было 30 (10 × 3) растений в трех биологических повторах, эксперименты проводили 3 раза. Данные - это средние значения ± SE. В каждой серии экспериментов столбцы, обозначенные одной и той же буквой, существенно не различались при p Уровень <0,05 согласно множественному тесту Дункана
Распределение MWCNT можно легко оценить с помощью просвечивающей электронной микроскопии (ПЭМ). Результаты, показанные на фиг. 3, показали, что MWCNT при экзогенном применении могут абсорбироваться проростками томатов, и было обнаружено, что MWCNT распределяются в клетках корня. Этот результат можно понять, поскольку корни проростков непосредственно культивируют в жидком растворе, содержащем MWCNT.
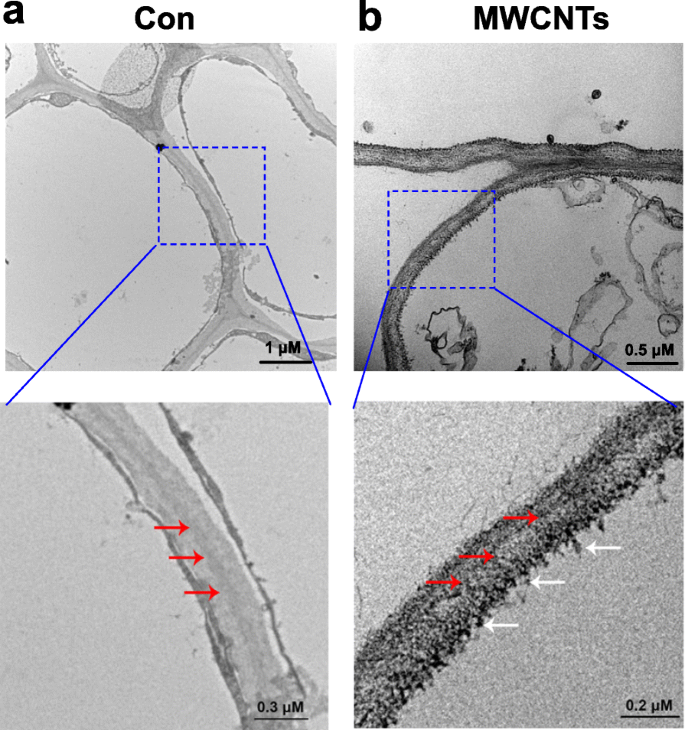
Распределение MWCNT в корнях томатов. ПЭМ-изображения 3-дневных проростков томатов, обработанных дистиллированной водой (Con; a ) или 5 мг / мл MWCNT ( b ) за 1 день. Красная стрелка указывает на клеточную стенку, а белая стрелка указывает на MWCNT
Синтез NO, индуцированный MWCNT, и последующее образование LR были чувствительны к cPTIO, поглотителю NO
Чтобы исследовать, участвует ли NO также в индуцированном MWCNT образовании LR, оценивали функцию NO в образовании LR, вызванную MWCNT, путем манипулирования уровнями эндогенного NO с использованием NO-высвобождающего соединения и поглотителя. Подобно предыдущим результатам [31], введение нитропруссида натрия (SNP) могло привести к индукции образования LR, и при совместном применении SNP и MWCNT наблюдался аддитивный ответ (рис. 4). Когда добавляли калиевую соль 2- (4-карбоксифенил) -4,4,5,5-тетраметилимидазолин-1-оксил-3-оксида (cPTIO; поглотитель NO), промотирующие реакции в образовании LR, вызванные MWCNT, были значительно ослаблен. В одиночку cPTIO может ингибировать развитие LR по сравнению с контролем, не содержащим химикатов, что указывает на важную роль эндогенного NO в органогенезе корней.
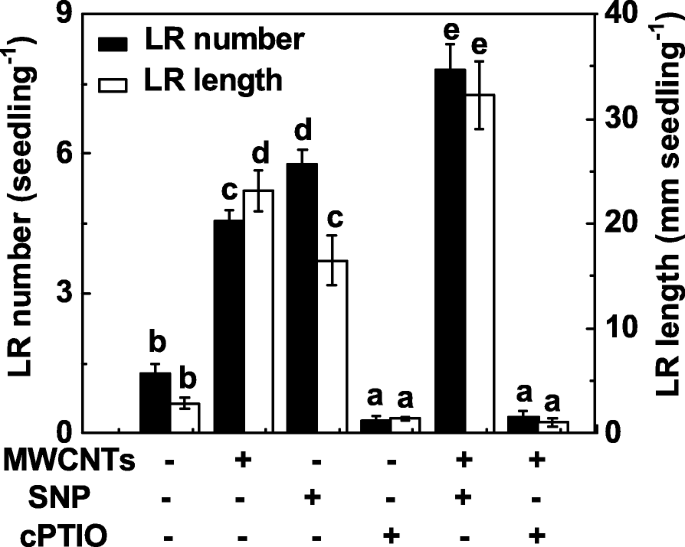
MWCNT-индуцированное образование LR было чувствительно к удалению эндогенного NO с помощью cPTIO, его поглотителя. Трехдневные проростки томатов обрабатывали дистиллированной водой, 5 мг / мл MWCNT, 0,1 мМ SNP, 0,2 мМ cPTIO по отдельности или в комбинации в течение 3 дней. Затем определяли количество и длину появившихся боковых корней (> 1 мм) на проросток. Всего было 30 (10 × 3) растений в трех биологических повторах, эксперименты проводили 3 раза. Данные - это средние значения ± SE. В каждой серии экспериментов столбцы, обозначенные одной и той же буквой, существенно не различались при p Уровень <0,05 согласно множественному тесту Дункана
Чтобы дополнительно оценить важную роль эндогенного NO в ответе MWCNT, сначала с помощью метода реагентов Грейсса была определена динамика выработки NO in vivo. Во время вышеуказанного определения идентичный фильтрат, предварительно обработанный cPTIO, считался холостым для получения точных результатов. Было замечено, что продукция NO в корнях проростков томатов резко увеличивалась до 24 часов после обработки MWCNT, а затем восстанавливалась до исходных уровней (48 часов; рис. 5а). Выше максимального уровня эндогенного NO, вызванного MWCNT в течение 24 часов, очевидно, отменяется cPTIO, поглотитель NO, что свидетельствует о специфической роли NO.
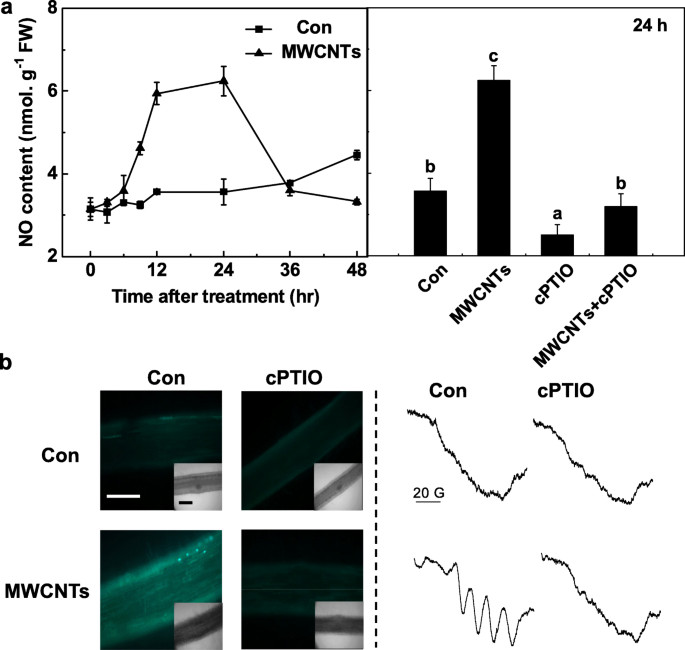
MWCNT-индуцированная продукция NO блокируется cPTIO, поглотителем NO. Трехдневные проростки томатов обрабатывали дистиллированной водой и МУНТ 5 мг / мл с 0,2 мМ cPTIO или без него, соответственно. а Изменения активности NR (слева) и продукции NO (справа), определенные с использованием метода реагентов Грейсса. б После воздействия в течение 24 часов сигнал NO анализировали с помощью LSCM (слева) и ЭПР (справа). Шкала шкалы =0,1 мм. Данные - это средние значения ± SE. Столбцы, обозначенные одной и той же буквой, существенно не различались на p Уровень <0,05 согласно множественному тесту Дукана
Для подтверждения приведенных выше результатов были приняты как LSCM, так и ESR. Во-первых, изменения уровней эндогенного NO в корнях проростков томатов отслеживали путем маркировки NO с помощью проницаемого для клеток, достаточно NO-специфичного флуоресцентного зонда DAF-FM DA и визуализации с помощью LSCM. Подобно предыдущим результатам (фиг. 5a), в присутствии cPTIO, увеличенная DAF-FM-зависимая интенсивность флуоресценции, запускаемая MWCNTs, была в значительной степени устранена (фиг. 5b). Эти результаты предполагают, что флуоресценция, запускаемая DAF-FM, связана с эндогенными уровнями NO в корнях проростков томатов.
Продукция NO, индуцированная MWCNT, была подтверждена спектроскопией ЭПР. Как и ожидалось, корни проростков, обработанные в течение 24 ч МУНТ, имели типичный триплет комплекса NO со сверхтонкой структурой. Однако добавление cPTIO аннулировало вышеуказанный сигнал, указывая на то, что воздействие MWCNT действительно привело к сильному образованию NO (рис. 5b). В совокупности эти данные предполагают, что синтез NO может потребоваться для инициируемого MWCNT образования LR в проростках томатов.
NR может нести ответственность за индуцированное MWCNT образование NO и последующее образование LR
Поскольку NR и подобная млекопитающим NOS являются двумя основными ферментами, связанными с синтезом NO в растениях, в последующем эксперименте были применены как вольфрамат (ингибитор NR), так и NAME (ингибитор NOS млекопитающих). Здесь обработка вольфраматом существенно подавляла стимулирование образования LR в обработанных MWCNT корнях проростков томатов (фиг. 6). Для сравнения, индукция образования LR, запускаемая MWCNT, не сильно ингибировалась добавлением NAME, что указывает на то, что NOS, подобная млекопитающим, может не быть целевым синтетическим ферментом NO, ответственным за продукцию NO, вызванную MWCNT. Также было замечено, что небольшое, но не значимое снижение образования LR наблюдалось в проростках томатов при отдельном применении вольфрамата или NAME.
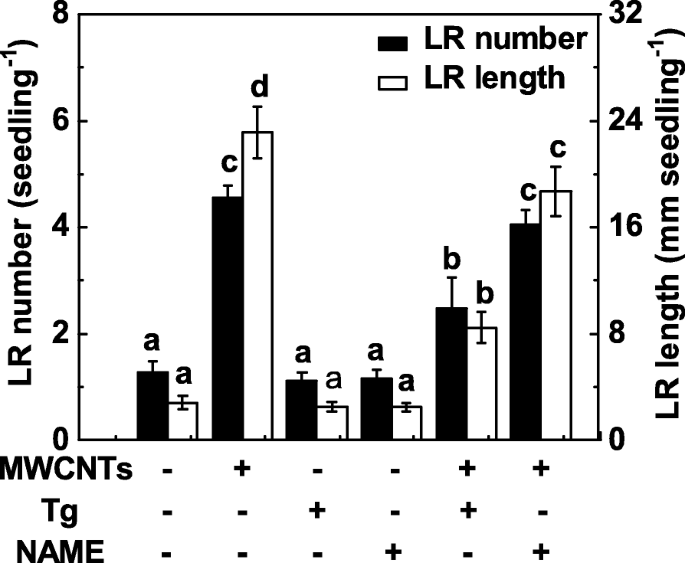
Изменения образования LR в ответ на MWCNT и два ингибитора синтеза NO. Трехдневные проростки томатов обрабатывали 5 мг / мл MWCNT, 20 мкМ вольфрамата (Tg), 0,2 мМ NAME по отдельности или в комбинации в течение 3 дней. Затем определяли количество и длину появившихся боковых корней (> 1 мм) на проросток. Всего было 30 (10 × 3) растений в трех биологических повторах, эксперименты проводили 3 раза. Данные - это средние значения ± SE. В каждой серии экспериментов столбцы, обозначенные одной и той же буквой, существенно не различались при p Уровень <0,05 согласно множественному тесту Дункана
Роль NR в инициированном MWCNT образовании LR была дополнительно исследована путем мониторинга продукции NO в ответ на нанесенные MWCNT с вольфраматом или без него. По сравнению с изменениями в продукции эндогенного NO (рис. 5a), анализ динамики активности NR показал аналогичную тенденцию (рис. 7a), также достигающую пика через 24 часа после обработки MWCNT. Эти результаты предполагают, что MWCNT-индуцированное увеличение продукции NO может в основном быть результатом повышенной активности NR. Соответственно, ингибирование NR-зависимой продукции NO вольфраматом было подтверждено с использованием метода реагентов Грейсса (рис. 7b), LSCM и EPR (дополнительный файл 1:рисунок S1).
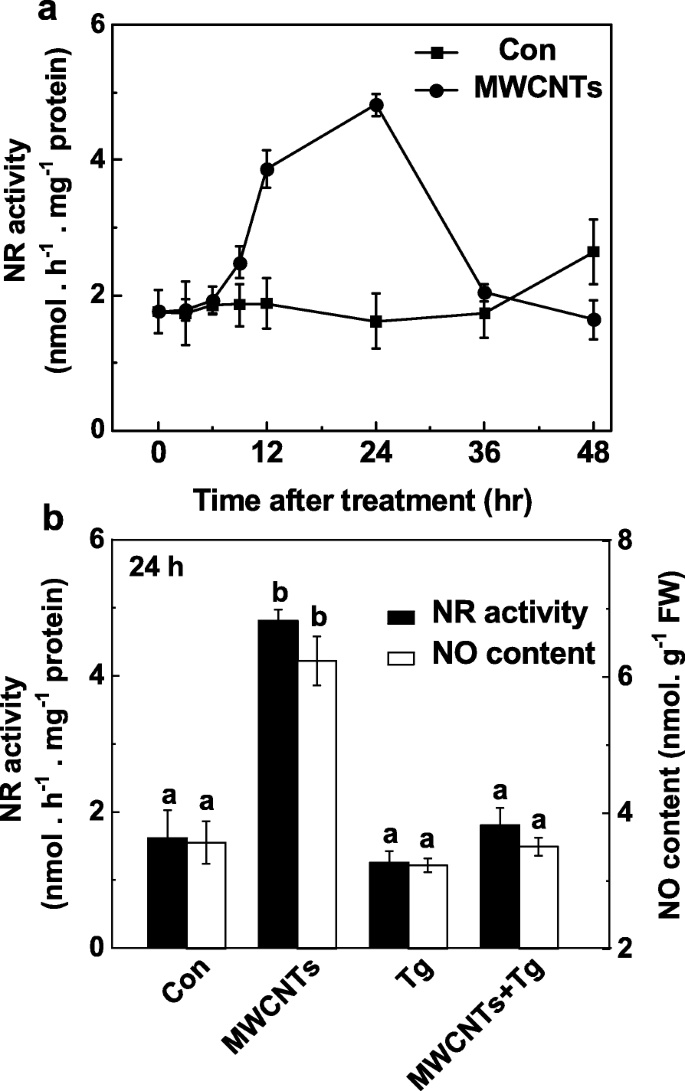
MWCNT-индуцированная продукция NO блокируется вольфраматом, ингибитором NR. Трехдневные проростки томатов обрабатывали дистиллированной водой и MWCNT с концентрацией 5 мг / мл с 20 мкМ вольфрамата (Tg) или без него. Изменения в активности NR ( a ) и НЕТ производства ( b ) определяли методом реактива Грейсса. Данные - это средние значения ± SE. Столбцы, обозначенные одной и той же буквой, существенно не различались на p Уровень <0,05 согласно множественному тесту Дукана
Обсуждение
Фитотоксичность - важный фактор в понимании потенциального воздействия наночастиц на окружающую среду [4, 7, 61,62,63]. Многочисленные доказательства показали, что MWCNT токсичны для растений, в том числе вызывают окислительное повреждение, ингибируют прорастание семян, рост корней и развитие [11, 63, 64]. Однако, будучи фенотипом SIMR, разветвление корней за счет образования боковых корней является важным компонентом приспособляемости корневой системы к различным сигналам окружающей среды [17]. В этой работе мы интегрировали биологический, фармакологический и биохимический анализ, чтобы показать участие NR-опосредованного производства NO в MWCNT-индуцированном образовании LR, по крайней мере частично, в наших экспериментальных условиях. Кроме того, функция NO в органогенезе корней, стимулируемая MWCNT, подчеркивает центральную роль этого второго мессенджера, участвующего в процессе развития растений и адаптации к стрессу [29,30,31,32,33, 37, 38].
Во-первых, мы подтвердили, что МУНТ 5 мг / мл (OD 6–12 нм) могут проникать в ткани корня (рис. 3). После этого наблюдали индукцию образования LR томатов (рис. 1), имитирующую индукционную роль NAA и SNP (рис. 4), хорошо известного соединения, высвобождающего NO [30, 31]. Подобные индуцирующие реакции были обнаружены у смолистых деревьев [41], салата [42] и арабидопсиса [43] при заражении MWCNTs (OD 6-13 нм, около 9,5 нм и 30-40 нм, соответственно). Например, применение либо первичных MWCNT (p-MWCNT), либо MWCNT с карбоксильными группами (c-MWCNT) (средний диаметр 9,5 нм) может способствовать развитию LR в проростках салата [42]. Напротив, ингибирование первичного корня и даже образование LR одновременно были обнаружены в растениях сои при воздействии MWCNT (OD 20–70 нм [40];). Сравнивая с данными по внешнему диаметру MWCNT (Таблица 1), мы предположили, что MWCNT-эффекты на образование LR варьировались в зависимости от их диаметра, показывая промотирование с меньшим диаметром и ингибирование с большим диаметром. Безусловно, связанный с этим механизм требует тщательного изучения. Подобное явление было подтверждено в отношении солеустойчивости растений [50]. В сочетании с приведенными выше результатами был сделан вывод, что функция наноматериалов может отличаться от вида, и наоборот, разные типы наноматериалов могут вызывать различные биологические эффекты. Однако нелегко исключить другие влияющие факторы, такие как разные дозы MWCNT [48] и даже условия роста растений.
По сравнению с другими наноматериалами, включая SWCNT, графен и AC с идентичной концентрацией, максимальная индукция образования LR и даже токсические эффекты на рост побегов наблюдались в MWCNTs (рис. 2). Это может быть связано с особыми физическими характеристиками MWCNTs, одного типа наноматериалов, которые имеют высокую электропроводность, большую удельную поверхность, высокое соотношение сторон и замечательную термическую стабильность [65]. О токсическом воздействии наноматериалов широко сообщалось в отношении огурцов, капусты, моркови, лука и т. Д. [66, 67].
Многочисленные доказательства показали, что NO, действуя как сигнальная молекула, может регулировать широкий спектр процессов растения от адаптации к окружающей среде до развития, последний из которых включает прорастание семян и органогенез корней [29, 68,69,70,71,72,73 ]. Наш последующий эксперимент показал, что NO может участвовать в индуцированном MWCNT образовании LR. Хотя было применено несколько методов визуализации продукции NO в растительных клетках, недостатки, в том числе отсутствие чувствительности и вмешательство со стороны NO-независимых молекул, могут существовать в каждом методе [74]. Таким образом, в наших экспериментальных условиях были применены три метода, отвечающие за визуализацию и определение NO, включая метод реагента Грейсса, LSCM и ЭПР, вместе с применением cPTIO, поглотителя NO. Используя три метода, мы наблюдали, что повышенная выработка эндогенного NO, индуцированная MWCNT в проростках томатов, устраняется cPTIO (рис. 5), поглотителем NO [30,31,32]. Важно отметить, что этот процесс коррелировал с биологическим ответом на развитие LR, индуцированного MWCNT, который был серьезно заблокирован при одновременном применении cPTIO (рис. 4).
Дальнейшая оценка этих ответов и потенциального источника (ов) NO, индуцированного экзогенно нанесенными MWCNT, показала, что продукцию NO и последующее образование LR можно отнести к активности NR. У растений продукция NO в основном генерируется из NR и NOS-подобного белка млекопитающих [28]. Однако ген NOS у растений до сих пор не идентифицирован [35, 75, 76], хотя некоторые эксперименты с использованием ингибиторов фермента NOS млекопитающих предоставили некоторые доказательства L-аргинин-зависимого пути продукции NO [36, 76]. Подтверждено, что NR является наиболее важным источником NO в растениях [28]. Предыдущие исследования показали, что NR-зависимая продукция NO действует как связанный с нитратами сигнал, участвующий в регуляции архитектуры корня [32, 33]. Кроме того, NR-зависимая продукция NO была тесно связана с акклиматизацией к холоду [38], толерантностью к солености [50] и закрытием устьиц, вызванным абсцизовой кислотой [77]. Наши результаты также показали, что вольфрамат (ингибитор NR), очевидно, нарушает MWCNT-индуцированное образование LR, особенно в длине LR (рис. 6). Напротив, при использовании l-NAME (ингибитор NOS млекопитающих) наблюдалось только небольшое уменьшение длины LR и не наблюдалось значительной разницы в количестве LR. Соответственно, биохимический анализ показал, что активность NR явно увеличивалась за счет MWCNT (фиг. 7a), параллельно с изменениями в продукции NO (фиг. 5a). Вышеуказанные ответы могут быть полностью заблокированы вольфраматом (рис. 7a, дополнительный файл 1:рис. S1). Таким образом, мы пришли к выводу, что повышенная продукция эндогенного NO, индуцированная MWCNT, в основном связана с путем NR. Certainly, further genetic evidence should be investigated.
Conclusion
In summary, we provide evidence to show that MWCNT-induced NO production via NR might be required for tomato lateral root formation and this was summarized in Fig. 8. Importantly, above findings provide insights into the intricate molecular mechanism of MWCNTs functions in plants.

Schematic representation of the proposed MWCNT-induced tomato lateral root formation mainly via NR-dependent NO production. The role of mammalian NOS-like enzyme was preliminarily ruled out
Доступность данных и материалов
Все данные полностью доступны без ограничений.
Сокращения
- CO:
-
Carbon monoxide
- cPTIO:
-
2-(4-Carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide potassium salt
- DAF-FM DA:
-
4-Amino-5-methylamino-2′,7′-difluorofluorescein diacetate
- EPR:
-
Electron paramagnetic resonance
- GO:
-
Оксид графена
- H 2 :
-
Hydrogen gas
- H 2 О 2 :
-
Hydrogen peroxide
- l-NAME:
-
N G -Nitro-l-arginine methyl ester
- LR:
-
Lateral root
- LSCM:
-
Laser scanning confocal microscopy
- MWCNT:
-
Multi-walled carbon nanotubes
- NAA:
-
1-Naphthylacetic acid
- NO:
-
Nitric oxide
- NOS:
-
Nitric oxide synthase
- NR:
-
Nitrate reductase
- ROS:
-
Активные формы кислорода
- SIMR:
-
Stress-induced morphogenic response
- SNP:
-
Sodium nitroprusside
- SWCNTs:
-
Single-walled carbon nanotubes
- ТЕМ:
-
Просвечивающая электронная микроскопия
- Tg:
-
Tungstate
Наноматериалы
- Углеродные нанотрубки расширяют границы гибкой электроники
- Углерод в океане, полученный в атомном масштабе
- Преимущества протезов из углеродного волокна
- Оксинитрид бора и углерода как новый безметалловый фотокатализатор
- Раскрытие атомной и электронной структуры углеродных нановолокон с набором чашек
- Улучшенное фотоэлектрохимическое определение мочевой кислоты на электроде из стеклоуглерода, модифицирова…
- Исследование новой червеобразной мицеллярной системы, усиленной наночастицами
- Каркасы nHAC / PLGA, гибридизированные с оксидом графена, способствуют пролиферации клеток MC3T3-E1
- Сравнительное исследование электрохимических, биомедицинских и тепловых свойств природных и синтетических…
- Уменьшите чувствительность CL-20 за счет улучшения теплопроводности за счет углеродных наноматериалов