


Тиакаликс [4] арены устраняют ингибирующее влияние катионов Zn на активность миозиновой АТФазы
Аннотация
Многочисленные женские репродуктивные аномалии вызваны нарушениями гладкой мускулатуры матки (миометрия). Тяжелые металлы отрицательно влияют на сократительную способность гладких мышц матки. Хотя цинк является важным биогенным элементом для большинства организмов, высокие дозы этого элемента токсичны. Исследование 0,5-5 мМ Zn 2+ влияние на активность АТФазы миозина S1 из матки обнаружило, что 5 мМ Zn 2+ катионы обладают наиболее выраженным ингибирующим действием. Расчет кинетических параметров ( K м и V макс , ATP ) выявили, что кажущаяся максимальная скорость гидролиза АТФ, катализируемого миозином, в присутствии 5 мМ Zn 2+ уменьшилась в 1,6 раза. Значение К м для гидролиза АТФ миозином S1 в присутствии Zn 2+ статистически не меняется, хотя имеет тенденцию к снижению. Было установлено, что активность АТФазы миозина S1 матки не зависит от концентрации Mg 2+ в присутствии 5 мМ Zn 2+ . Также было продемонстрировано, что тетрагидрокситиакаликс [4] арен-тетрасульфосфонат (C-798) и тетрагидрокситиакаликс [4] арен-тетрафосфонат (C-800) восстанавливали активность миозин S1-АТФазы до контрольного уровня в присутствии 5 мМ Zn 2+ . Один из наиболее вероятных механизмов восстановления защитного действия этих тиакаликс [4] аренов основан на их способности хелатировать катионы тяжелых металлов из инкубационной среды. Молекулярный докинг C-798 и C-800 в область миозина S1 показал, что эти тиакаликс [4] арены могут взаимодействовать с катионной связью Zn с помощью аминокислотных остатков миозина вблизи активного сайта АТФазы. Следовательно, тиакаликс [4] арены могут ослаблять взаимодействие между этим катионом и миозином S1. Было высказано предположение, что полученные результаты могут быть использованы для дальнейших исследований с целью использования этих тиакаликс [4] аренов в качестве фармакологических соединений в случае отравления высокими концентрациями цинка.
Фон
Проблема загрязнения окружающей среды тяжелыми металлами и поиск путей снижения их воздействия на живые объекты является актуальной [1, 2].
Цинк является важным биогенным элементом для большинства организмов. [3]. Ионы цинка образуют комплексы с множеством белков, которые выполняют жизненно важные метаболические функции. Ион цинка входит в состав не менее 300 металлоферментов, которые катализируют более 50 различных биохимических (физиологических) реакций [4, 5] .
Однако цинк - тяжелый металл. Его можно найти в группе IIb периодической таблицы элементов вместе с двумя токсичными металлами кадмием и ртутью. Тем не менее, цинк считается относительно нетоксичным для человека [6]. Этот элемент ядовит только в чрезмерных дозах [7].
Устный LD 50 для цинка составляет около 3 г / кг массы тела согласно базе данных TOXNET Национальной медицинской библиотеки США. Он более чем в 10 раз выше кадмия и в 50 раз выше ртути [6]. Превышение нормальной концентрации этого микроэлемента у человека чаще всего вызвано приемом лекарственных препаратов и биологически активных добавок, содержащих в своем составе избыточный цинк. Регистрировались как отдельные случаи отравления цинком в результате употребления в пищу продуктов, хранящихся в оцинкованных или полностью оцинкованных емкостях. Оксид, хлорид и сульфат цинка широко используются в промышленности для производства стекла; в производстве искусственных волокон, цинковых красок, керамики, спичек и стоматологического цемента; в целлюлозно-бумажной промышленности, для консервирования древесины, лужения и пайки.
Потребление высокой концентрации цинка изменило иммунный ответ [8]. Согласно некоторым эпидемиологическим исследованиям, повышенные уровни Zn, Al, Cu и Fe в головном мозге могут способствовать развитию или прогрессированию болезни Альцгеймера [9, 10].
Тяжелые металлы могут влиять на женскую репродуктивную функцию на разных стадиях, таких как начало жизни плода, раннее развитие и созревание. Катионы тяжелых металлов также могут быть причиной недостаточной фертильности, бесплодия, задержки внутриутробного развития, самопроизвольных абортов, пороков развития, врожденных дефектов, послеродовой смерти, преждевременного старения, а также нарушений обучения и поведения [11, 12].
Сократительная функция матки связана с активностью белкового комплекса - актомиозина, в котором миозин проявляет ферментативную активность, а именно способность гидролизовать АТФ. Миозин-АТФаза, локализованная в каталитическом домене субфрагмента-1 (S1 или голова), преобразует химическую энергию, депонированную в макроэргических связях АТФ, в механическое движение. В результате миозин движется по актиновой нити, вызывая сокращение мышц. Поэтому гидролиз АТФ, катализируемый миозином, считается одним из важных процессов в молекулярном механизме функции миометрия [13, 14].
Субфрагмент-1 миозина представляет собой N-концевую часть тяжелой цепи миозина, которая состоит из двух доменов:N-концевого глобулярного моторного (каталитического) домена, содержащего сайт АТФ-азы и сайт связывания актина, и регуляторного домена, или рычаг, отвечающий за движение миозина по актиновым филаментам. Ядро моторного домена миозина образовано центральным семицепочечным β-слоем, окруженным α-спиралями. Большой структурный домен, который составляет шесть из семи нитей центрального β-листа, обычно называют верхним доменом 50 кДа (U50). Большая щель отделяет верхний домен 50 кДа от четко определенного структурного нижнего домена 50 кД (L50), который образован аминокислотными остатками от 465 до 590. Актин-связывающая область и нуклеотид-связывающий сайт миозина находятся на противоположные стороны семинитевого β-листа с фосфатной частью нуклеотида в задней части нуклеотид-связывающего кармана. P-петля, переключатель 1 и переключатель 2 расположены в верхнем домене 50 кДа, близко к вершине большой щели. Все три нуклеотид-связывающих мотива контактируют с фосфатной частью нуклеотида в задней части нуклеотид-связывающего кармана и действуют как сенсоры γ-фосфата [15].
В наших предыдущих исследованиях было обнаружено, что катионы тяжелых металлов ингибируют активность миозин-АТФазы гладких мышц матки [16, 17], что может отрицательно влиять на сократительные свойства миометрия.
Неблагоприятное влияние тяжелых металлов на сократительную способность матки требует разработки фармакологических веществ, которые могут устранить эти вредные эффекты.
Каликсарены в настоящее время привлекают внимание исследователей как предполагаемые искусственные эффекторы различных биохимических процессов. Эти соединения представляют собой синтетические макроциклические фенольные олигомеры, имеющие чашеобразную структуру. Их верхний и нижний края могут быть функционализированы различными химическими заместителями. Каликс [4] арены образованы четырьмя функционализированными ареновыми фрагментами и характеризуются довольно гибкой конформацией макроцикла. Каликс [4] арены показали низкую токсичность матрикса и способность проникать в клетки. Следовательно, эти соединения рассматриваются как перспективные агенты для разработки новых эффективных лекарств [18, 19].
Перспективным классом таких веществ являются водорастворимые тиакаликсарены [18], обладающие группами комплексообразования с металлами на макроциклической молекулярной платформе. Каликсарены из-за их способности образовывать супрамолекулярные комплексы с (био) катионами металлов также использовались в биомедицинских исследованиях в качестве экстрагентов тяжелых металлов [20,21,22].
Ранее мы показали, что тетрагидрокситиакаликс [4] арен-тетрасульфонат (С-798) устраняет ингибирующие эффекты Pb 2+ , CD 2+ , и Ni 2+ о гидролизе АТФ, катализируемом миозином S1 из миометрия свиньи [23].
Это исследование было направлено на изучение влияния высоких концентраций катионов цинка и их совместного действия с тетрагидрокситиакаликс [4] арен-тетрасульфонатом (С-798) и тетрагидрокситиакаликс [4] арен-тетрафосфонатом (C-800) на миозин. Активность S1-АТФазы из матки. Это исследование было необходимо для проверки способности этих тиакаликсаренов устранять побочные эффекты высоких концентраций цинка на ферментативную активность миозина матки.
Тиакаликс [4] арены C-798 и C-800 состоят из чашки, образованной четырьмя фенольными фрагментами, модифицированными по верхнему краю четырьмя анионными сульфонатными и четырьмя фосфонатными группами соответственно. Оба тиакаликс [4] арена имеют гидроксильные группы и атомы двухвалентной серы, плотно расположенные на нижнем ободе, что позволяет им хелатировать тяжелые металлы с образованием стабильных комплексов металлов [21] (рис. 1).
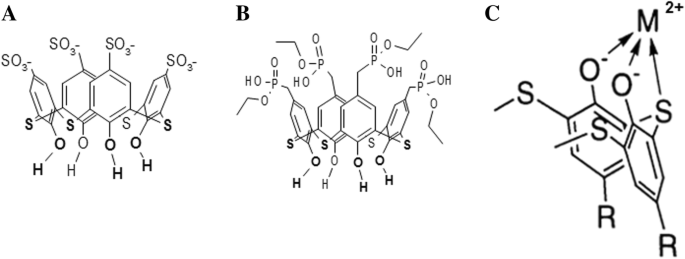
Химическая структура тетрагидрокситиакаликс [4] арентетрасульфоната (C-798) ( a ), тетрагидрокситиакаликс [4] арентетрафосфонат (C-800) ( b ), и схема хелатного комплекса тиакаликсарена с катионом металла на нижнем ободе (перевернутое положение) ( c )
Данная работа является результатом совместного проекта Института биохимии Палладина и Института органической химии НАН Украины, посвященного взаимодействию миозин-АТФазы миометрия с каликс [4] аренами, которые являются ингибиторами или активаторами (эффекторами) миозиновой АТФазы матки.
Результаты
Зависимость активности АТФазы миозина S1 от Zn 2+ Концентрация
Было обнаружено, что наиболее выраженный ингибирующий эффект на активность АТФазы миозина S1 из матки был при 5 мМ (43 ± 8%, M ± SD) для катионов Zn. Диапазон концентраций Zn 2+ составляла 0,5-5 мМ в инкубационной среде (содержащей 3 мМ АТФ, 5 мМ Mg 2+ и 0,01 мМ Ca 2+ ). Сто процентов - это значение активности АТФазы без добавления катионов Zn (контроль) (рис. 2). Таким образом, неблагоприятные эффекты катионов Zn на гидролиз АТФ миозина S1 были дополнительно изучены с 5 мМ Zn 2+ .
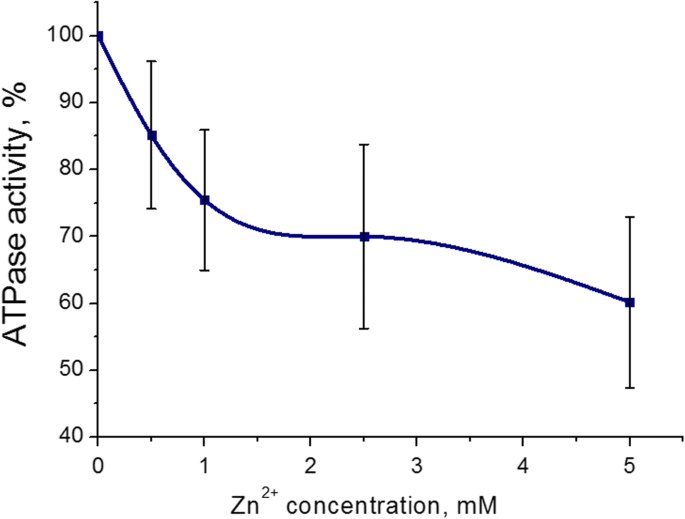
Миозин S1 АТФазная активность миометрия в присутствии 0,5–5,0 мМ катионов Zn (M ± SD, n =6). 100% - значение активности АТФазы без добавления катионов Zn
Зависимость активности АТФазы миозина S1 от концентрации АТФ в присутствии 5 мМ Zn 2+
Zn 2+ влияние на сродство активности АТФазы миозина S1 к его субстрату (АТФ). Увеличение концентрации АТФ в инкубационной среде с 0,5 до 5 мМ при фиксированном MgCl 2 концентрация (5 мМ) как в контроле, так и в присутствии 5 мМ Zn 2+ привели к куполообразному графику с максимальным значением активности АТФазы при 3 мМ АТФ. Величина ферментативной активности на этом пике в присутствии цинка была на 30% ниже, чем у контроля (рис. 3). Графики зависимости активности АТФазы миозина S1 от концентрации АТФ в контроле и наличия 5 мМ Zn 2+ в восходящей части были линеаризованы по методу Лайнуивера – Берка [27]. Расчет кинетических параметров, а именно мнимой постоянной Михаэлиса ( K м ) и максимальная скорость АТФазы миозина S1 для АТФ ( V макс, ATP ) показал, что V макс, ATP ферментативной активности миозина в присутствии 5 мМ Zn 2+ снизилась в 1,6 раза (38 ± 7 и 22 ± 6 нмоль Pi / мин на 1 мг белка в контроле и наличие Zn 2+ соответственно, n =5). Значение К м для гидролиза АТФ миозином S1 статистически не изменяется, хотя имеет тенденцию к снижению (0,49 ± 0,15 мМ в контроле, 0,38 ± 0,12 мМ в присутствии Zn 2+ ; M ± SD; нет =5).
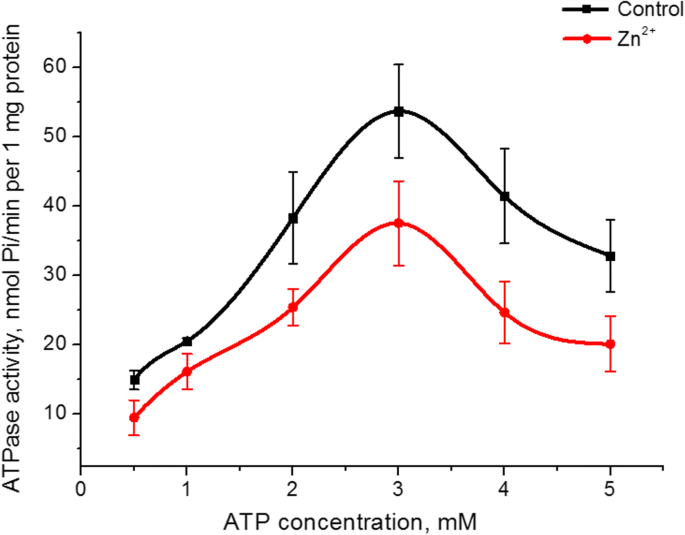
Влияние 0,5–5 мМ АТФ на активность АТФазы миозина S1 из матки в присутствии 5 мМ Zn 2+ по сравнению с контролем (M ± SD, n =5)
Зависимость активности АТФазы миозина S1 от Mg 2+ Концентрация в присутствии 5 мМ Zn 2+
Эффект 5 мМ Zn 2+ на Mg 2+ Изучена концентрационная зависимость от АТФазной активности миозина матки. Повышение Mg 2+ концентрация в инкубационной среде от 0,5 до 5 мМ при фиксированной концентрации АТФ (3 мМ) в присутствии 5 мМ Zn 2+ не приводит к изменению активности АТФазы миозина S1. В то же время Mg 2+ Выявлена концентрационная зависимость активности АТФазы в контроле (в стандартных условиях). Наивысший уровень гидролиза миозинового АТФ в контроле был достигнут при 3 мМ Mg 2+ . (Рис.4). Следовательно, активность фермента миозина S1 в матке не зависит от концентрации Mg 2+ в присутствии цинка в высоких концентрациях (5 мМ).
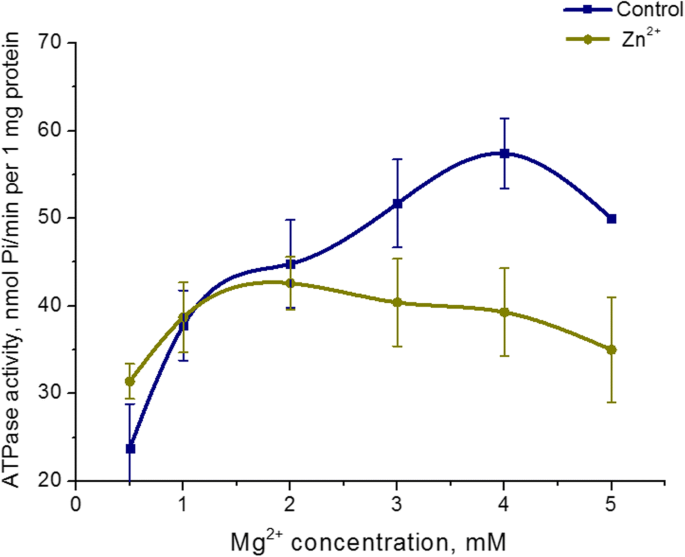
Зависимость активности АТФазы миозина S1 от Mg 2+ концентрация в присутствии 5 мМ Zn 2+ по сравнению с контролем (M ± SD, n =6)
Zn-связывающие сайты в Myosin S1
Компьютерное моделирование показывает, что катионы Zn имеют несколько участков связывания в миозиновой головке. Один из них расположен в нижней части щели между верхним и нижним субдоменами 50 кДа, рядом с сайтом связывания нуклеотидов и непосредственно возле P-петли. Zn 2+ координирован с атомами кислорода Glu177 (длина связи 0,23 и 0,39 нм), с атомом кислорода Ser178 (длина связи 0,31 нм) и Arg236 (длина связи 0,32 нм).
Другая Zn-связывающая область расположена внизу верхнего субдомена размером 50 кДа (Leu218-Asp463, Glu605-Phe621) и рядом с переключателем 1 (Gly233-Phe246) и P-петлей. Zn 2+ может координироваться с атомом кислорода Glu327 (длина связи 0,21 нм), с атомом кислорода Glu326 (длина связи 0,34 нм) и атомом кислорода Asp323 (длина связи 0,32 нм). Катион Zn может также взаимодействовать с миозином S1 в области, которая контактирует с переключателем 2, взаимодействуя с Glu 465 (0,24 нм), Asp468 (длина связи 0,31 нм) и Leu653 (длина связи 0,37 нм). Эта связывающая область находится рядом с сайтом связывания актина и зазором между верхним и нижним субдоменами 50 кДа. Дно этой щели находится в АТФ-связывающем кармане. Эти связующие Zn 2+ Домены миозина S1 играют важную роль в связывании и гидролизе АТФ. Эти области претерпевают сложные конформационные преобразования в процессе передачи энергии от сайта гидролиза АТФ к связывающей актин поверхности.
Тиакаликс [4] арены устраняют ингибирующее действие Zn 2+ на активность миозин-АТФазы
Сто микромолей растворов C-800 или C-798 в 50 мМ трис-HCl буфере (pH 7,2) добавляли в инкубационную среду, содержащую 5 мМ катионов Zn, для устранения негативного воздействия Zn 2+ на АТФазную активность миозина S1 гладких мышц матки. В качестве контроля его использовали в качестве ферментативной активности без добавления цинка и / или тиакаликс [4] аренов в среду инкубации. Было показано (рис. 5), что соединение C-800 не влияет на АТФазную активность миозина S1 миометрия. Хотя соединение C-798 проявляет небольшой ингибирующий эффект на активность АТФазы миозина S1, что, скорее всего, связано с экстракцией определенного количества Mg 2+ [21], необходимого для связывания АТФ в активном центре и его гидролиза, из инкубационной среды. Тем не менее, 100 мкМ C-798, а также C-800 устраняют ингибирующие эффекты 5 мМ Zn 2+ о процессе гидролиза АТФ, катализируемого миозином S1.
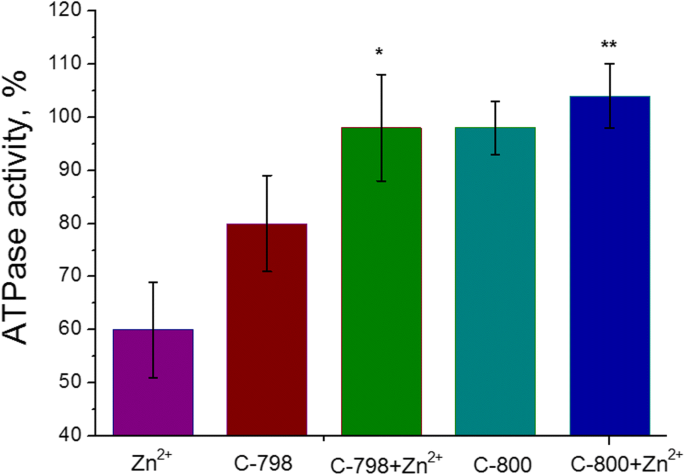
Влияние 100 мкМ C-798 и C-800 на АТФазную активность миозина S1 в присутствии 5 мМ Zn 2+ (M ± SD, n =5–6). 100% - значение активности АТФазы без добавления катиона Zn. Разница между «Zn» и «Zn + C-798», а также между значениями «Zn» и «Zn + C-800» статистически значима ( p <0,05) и обозначены * и ** соответственно
Вероятные механизмы восстанавливающего действия C-798 и C-800 на активность АТФазы миозина S1 в присутствии Zn 2+
Один из наиболее вероятных механизмов восстанавливающего действия C-798 и C-800 на активность АТФазы миозина S1 в Zn 2+ Присутствие может заключаться в способности тиакаликс [4] аренов связывать катионы Zn и, следовательно, исключать эти катионы из среды инкубации. Было интересно, могут ли эти тиакаликс [4] арены связываться с катионами цинка, которые уже связаны с миозином.
Компьютерное моделирование показало, что тиакаликс [4] арены C-798 и C-800 с мостиковыми атомами серы между ароматическими кольцами находятся в конформации «конуса», стабилизированной внутриводородными связями между фенольными группами. Получена минимизированная по энергии структура этих каликс [4] аренов. Полная энергия C-798 после минимизации составила 64,5 ккал / моль. Наличие ионизированных групп на краях каликс [4] арена (в частности, нижних) значительно увеличивает вклад электростатических взаимодействий в общую энергию взаимодействия хозяин-гость. Мы также провели «минимизацию» C-798 - Zn 2+ сложный; его общая энергия составляла 83 ккал / моль.
C-798 был встроен в структуру миозина S1, взаимодействующего с катионом Zn, который ранее был связан с белком в области петли P. В этом случае Zn 2+ взаимодействует с атомами кислорода нижнего обода и мостиковой серой C-798 (O3, 0,21 нм; S1, 0,30 нм; O2, 0,34 нм). Показано, что Zn 2+ в некоторой степени отклоняется от аминокислотных остатков P-петли и ослабляет ее взаимодействие с атомом кислорода Glu177 (длина связи 0,43 нм) (рис. 6).
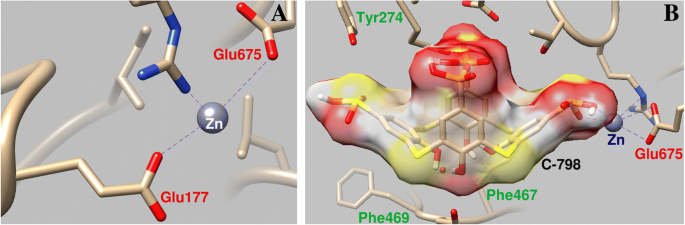
Геометрические параметры взаимодействия Zn 2+ с областью P-петли миозина S1 ( a ) и влияние C-798 на взаимодействие катиона Zn с этой областью ( b )
Фиксация C-798 в «полости» АТФ-связывающей области миозина происходит при участии нескольких аминокислотных остатков. В частности, гидрофобная корзина тиакаликсарена закреплена остатками ароматических аминокислот миозина Phe467 и Phe469; Отрицательно заряженные атомы кислорода тиакаликсарена взаимодействуют с положительно заряженными аминокислотными остатками Arg570, Asn572 и His689.
Изучение влияния C-798 на изменение Zn 2+ положение во время стыковки в области рядом с сайтом связывания миозина АТФ продемонстрировало, что Zn 2+ в присутствии тиакаликс [4] арен взаимодействует с атомами кислорода третьей сульфонильной группы (O16–0,26 нм; O15–0,27 нм), практически не взаимодействует с атомами кислорода Asp134 и Glu326, а координация с Glu327 значительно слабее. (Длина связи 0,43 нм) (рис.7). В этом случае тиакаликсарен фиксируется в «полости» белка с участием нескольких аминокислотных остатков. В частности, отрицательно заряженные атомы кислорода тиакаликсареновых сульфонильных групп взаимодействуют с положительно заряженными аминокислотными остатками миозина Lys188, Lys195 и Gln221.
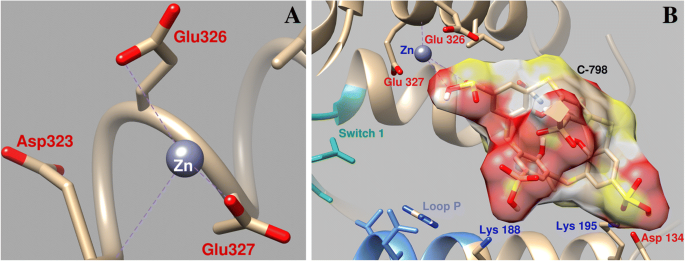
Геометрические параметры взаимодействия Zn 2+ в области около переключателя миозина 1 и P-петли миозина S1 ( a ) и влияние C-798 на взаимодействие Zn 2+ с этим регионом ( b )
Показано, что условная энергетическая сфера катиона Zn контактирует и даже несколько перекрывается с поверхностью распространения электростатических взаимодействий из-за присутствия атомов кислорода и атомов серы в C-798. Это свидетельствует о том, что исследуемый катион находится в достаточно тесном взаимодействии с отрицательно заряженными атомами верхней и нижней коронок С-798. Вероятно, что тиакаликсарен притягивает к себе Zn 2+ , в результате чего взаимодействие катионов с аминокислотными остатками фермента ослабляется.
Следовательно, результаты стыковки C-798 в области миозина S1, которая содержит Zn 2+ связаны, указывают на возможность взаимодействия функциональных групп C-798 с катионом Zn. При этом связь катиона Zn с аминокислотными остатками миозина S1 значительно ослабляется, а расстояние между ними увеличивается. В результате может быть устранено неблагоприятное влияние катиона Zn на АТФазную активность миозина.
Мы также провели компьютерное моделирование эффекта каликс [4] арена C-800 с докингом в области, близкой к АТФ-связывающей области миозина S1, для изменения геометрии катиона Zn. В то же время Zn 2+ находится в контакте с Arg236 (длина связи 0,37 нм), Glu675 (длина связи 0,41 нм) и атомами верхних функциональных остатков каликс [4] арена (H26, 2,36 нм; H30, 2,96 нм; C30, 0,31 нм; O5, 3,7 нм; O16, 0,4 нм; O7, 0,48 нм; и O11, длина связи 0,51 нм). Zn 2+ взаимодействует с каликс [4] ареном С-800, аналогично С-798. Катион также удаляется из предыдущего сайта связывания в этой области и находится в контакте с Asp468 (длина связи 0,2 нм) и атомами нижнего каликсаренового корона (C4, 1,96 нм; C14, 2,07 нм; S3, 2,16 нм; O2). 2,26 нм; C3 2,97 нм; C20 3,10 нм; O4 4,2 нм; C14 3,0 нм; и O3 4,1 нм).
Докинг C-798 и C-800 в области субфрагмента-1 миозина показал, что эти тиакаликс [4] арены могут взаимодействовать с катионами Zn, связывающимися с аминокислотными остатками миозина вблизи активного сайта АТФазы. Следовательно, их защитный эффект может быть результатом ослабления взаимодействия между этими катионами и миозином S1.
Обсуждение
Многочисленные женские репродуктивные аномалии вызваны нарушениями гладкой мускулатуры матки (миометрия). Тяжелые металлы отрицательно влияют на сократительную способность гладких мышц матки. Цинк тяжелого металла является важным биогенным элементом для большинства организмов; высокие дозы этого элемента токсичны [5]. Несколько исследований миллимолярных концентраций Zn 2+ на живых объектах были описаны ранее [32]. Мы обнаружили, что 5 мМ Zn обладают наиболее выраженным ингибирующим действием на активность миозина S1-АТФазы из матки. Таким образом, неблагоприятные эффекты катионов Zn на гидролиз АТФ миозина S1 были дополнительно изучены с этой концентрацией Zn 2+ . Расчет кинетических параметров АТФазы миозина S1 для АТФ показал, что V макс, ATP ферментативной активности миозина в присутствии 5 мМ Zn 2+ уменьшилась в 1,6 раза. Значение К м для гидролиза АТФ статистически не изменяется, хотя имеет тенденцию к снижению.
Миозин в физиологических условиях представляет собой Mg 2+ -зависимая АТФаза. Катион магния участвует в связывании АТФ в активном центре миозина, а также в гидролизе АТФ. Mg 2+ координируется в активном центре фермента с боковыми цепями аминокислотных остатков миозина Thr-186 и Ser-237, а также с β- и γ-фосфатными группами молекулы АТФ с образованием β- и γ- бидентатный комплекс, а также с активными молекулами воды, одна из которых осуществляет нуклеофильную атаку на γ-фосфатный АТФ [33, 34]. Мг 2+ взаимодействует с отрицательно заряженными фосфорными группами АТФ, поляризует их и, таким образом, способствует нуклеофильной атаке на терминальный γ-фосфат [14].
Было обнаружено, что активность АТФазы миозина S1 не чувствительна к присутствию Mg 2+ при концентрации Zn 5 мМ 2+ в отличие от контроля, когда цинк отсутствовал в инкубационной среде [35, 36].
Активность миозин-АТФазы зависит от природы катионов металлов и хорошо коррелирует с их ионным радиусом. Ионные радиусы Mg 2+ и Zn 2+ в растворах очень похожи (0,070 и 0,076 нм соответственно) [37]. Следовательно, взаимодействие Zn 2+ катионы с Mg 2+ -связи миозина. Таким образом, Mg 2+ сайты связывания могут быть заняты Zn 2+ катионы в высоких концентрациях. АТФазная активность миозина S1 в таких условиях может быть нечувствительной к катионам магния. Миозин содержит два сайта с высоким сродством к Mg 2+ , и Mg 2+ связанный на этих участках, играет важную физиологическую роль в процессе передачи энергии во время сокращения мышц. Есть еще несколько Mg 2+ -связывающие сайты помимо сайта АТФазы в молекуле миозина, различающиеся энергией связывания ионов магния и их сродством [35]. Следовательно, можно предположить, что Zn 2+ может также связываться с другими функционально важными участками миозина S1, которые влияют на связывание и гидролиз АТФ.
Компьютерное моделирование показывает, что у катионов Zn есть несколько связывающих участков в головке миозина, расположенных рядом с сайтом связывания АТФ, а именно P-петля, а верхний и нижний 50-кДа субдомены переключаются 2. Эти связывающие Zn 2+ Домены миозина S1 играют важную роль в связывании и гидролизе АТФ. Эти области претерпевают сложные конформационные преобразования в процессе передачи энергии от сайта гидролиза АТФ к связывающей актин поверхности.
Результаты анализа, полученные с помощью Zn 2+ стыковка с миозином S1 указывает на то, что ключевую роль в связывании этого катиона с молекулой миозина играет его взаимодействие с отрицательно заряженными группами аминокислотных остатков фермента, в частности, Glu и Asp.
Вредное влияние токсичных концентраций Zn 2+ катионов на активность АТФазы миозина S1 требует поиска фармакологических соединений, которые могут устранить действие этого металла. Объектами нашего исследования были тетрагидрокситиакаликс [4] арен-тетрасульфонат (C-798) и тетрагидрокситиакаликс [4] арен-тетрафосфонат (C-800), способные хелатировать переходные и тяжелые металлы с образованием стабильных комплексов металлов (рис. 1). Верхняя макроциклическая корона C-798 и C-800 содержит четыре анионных сульфонатных группы или четыре фосфоната соответственно, которые обеспечивают хорошую растворимость в воде тиакаликсарена и адгезию к молекулам белка за счет электростатических контактов с положительно заряженными атомами азота аминокислотных фрагментов [21]. ].
Докинг C-798 и C-800 в области S1 миозина показал, что эти тиакаликс [4] арены могут взаимодействовать с катионами Zn, связывающимися с аминокислотными остатками миозина вблизи активного сайта АТФазы. Следовательно, их защитный эффект может быть результатом ослабления взаимодействия между этими катионами и миозином S1. Было высказано предположение, что полученные результаты могут быть использованы для дальнейших исследований с целью использования этих тиакаликс [4] аренов в качестве фармакологических соединений в случае отравления высокими концентрациями цинка.
Выводы
Высокая концентрация (5 мМ) катиона Zn подавляла активность АТФазы миозина S1 из матки. Ингибированное влияние Zn связано с уменьшением максимальной скорости гидролиза АТФ, катализируемого миозином S1, в присутствии 5 мМ Zn 2+ . Значение К м для АТФ статистически не изменяется, хотя имеет тенденцию к снижению.
Тетрагидрокситиакаликс [4] арен-тетрасульфосфонат (C-798) и тетрагидрокситиакаликс [4] арен-тетрафосфонат (C-800) восстанавливали активность АТФазы миозина S1 до контрольного уровня в присутствии 5 мМ Zn 2+ .
Катионы Zn имеют несколько участков связывания в миозине S1, расположенных рядом с активным центром АТФазы. Докинг C-798 и C-800 в область миозина S1, которая содержит Zn 2+ Связанный, указывает на возможность взаимодействия этих тиакаликс [4] ареновых функциональных групп со связанными катионами Zn. Катионная связь Zn с аминокислотными остатками миозина S1 значительно ослабляется, а расстояние между ними увеличивается. В результате может быть устранено неблагоприятное влияние катиона Zn на АТФазную активность миозина.
Предполагается, что полученные результаты могут быть использованы для дальнейших исследований с целью использования этих тиакаликс [4] аренов в качестве фармакологических соединений при отравлении высокими концентрациями цинка.
Методы
Реагенты
Были использованы следующие реагенты:сывороточный альбумин, ЭГТА, ЭДТА, АТФ, аскорбиновая кислота, Трис, трицин, дитиотреитол, акриламид (Sigma, США), глицин (Merck, Германия), N, N'-метиленбисакриламид (Acros Organics, Бельгия. ) N, N, N ′, N′-тетраметилендиамин (Reanal, Венгрия) и реактивы отечественного производства (марка R). Растворы готовили на воде, очищенной на системе Crystal Bio (Adrona, Латвия). Водопроводимость была менее 0,1 мкСм. Концентрацию катионов двухвалентных металлов в растворе определяли по методу Мора.
Выделение субфрагмента-1 актомиозина и миозина
Актомиозин выделяли из гладкой мускулатуры матки свиней модифицированным методом Барани, как описано в [17]. Миозин S1 был получен из актомиозина свиньи модифицированным методом Сузуки [24]. The purity of the samples was controlled by PAAG-SDS electrophoresis [25].
ATPase Activity Assay
ATPase activity of myosin S1 was determined in a 96-well plate at 37 °C in an incubation medium (total volume 0.1 ml) of the following composition (mM):Tris-HCl buffer (pH 7.2), 20; KCl, 100; CaCl2 , 0.01; MgCl2 , 5; and ATP, 3 (standard conditions). Protein (myosin S1) concentration was 20 μg/ml. Incubation time was 5 min. Samples containing all components of the incubation medium without myosin S1 were taken as control of non-enzyme hydrolysis of ATP. The amount of inorganic phosphate released during ATP hydrolysis reaction was determined by the Chen method [26] by the measurement of optical absorbance of the solution at 820 nm using a microplate reader μQuwant (Biotek @ Instruments, Inc., USA) and specified as Pi nmol/min per 1 mg of protein.
The Zn 2+ and thiacalix[4]arene effects on the ATPase activity of myosin S1 were studied using standard incubation medium with solutions of ZnCl2 and thiacalix[4]arenes at the corresponding concentrations. The value of ATPase activity in the absence of ZnCl2 and/or calix[4]arenes in the incubation medium was taken as 100% (control).
Kinetic and Statistical Analysis
The values of the imaginary constant of Michaelis (K м ) and maximal rate of myosin S1 ATPase for ATP (V max, ATP ) were calculated using the graph of the dependence of ATPase activity on the ATP concentration according to Lineweaver–Burk method [27]. Statistical processing of the obtained data was performed using standard methods of variation statistics. Experimental data were analyzed by using the standard software “MS Office” and “Statistica 4.5.” The statistical comparisons were performed using two-way analysis of variances (ANOVA).
Thiacalix[4]Arene Synthesis and Characterization
Tetrahydroxy-thiacalix[4]arene-tetrasulphonate and tetrahydroxy-thiacalix[4]arene-tetraphosphonate were synthesized and characterized using NMR techniques and IR spectroscopy in the Phosphoranes Chemistry Department of the Institute of Organic Chemistry, NAS of Ukraine. Infrared and NMR spectroscopy confirmed the structure of these synthesized thiacalix[4]arenes. This thiacalix[4]arenes were dissolved in water.
Computer Modeling
Computer modeling of the interaction between ligands (thiacalix[4]arenes, Zn 2+ , model bindings) and receptor (myosin S1) was performed using AutoDock software, version 4.2 [28]. We used the three-dimensional enzyme structure with the 1b7t identifier in RSCB PDB in our research [29]. Computer modeling of the thiacalix[4]arene structural peculiarities was carried out using HyperChem 7.01. Molecular dynamics calculations were performed by the MM2 method with the semi-empirical methods (CNDO).
Program AutoDockTools was used for preliminary “processing” of interacting molecules. One hundred runs of Lamarkian genetic algorithms (population size, 100; the maximal number of energy evaluations, 10 6 ) were conducted. To analyze and visualize the docking results, we used the programs Chimera [30] and Yassara [31]. Calculation of the minimal total binding energy was implemented considering Van der Waals forces, electrostatic and hydrophobic interactions, and hydrogen bonds. The optimal ligand positions in the complex “receptor-ligand” were determined according to the energy values obtained by docking software calculator for binding energy in complex “receptor-ligand.” Thus, we selected a series of complexes with the lowest total energy and then calculated the optimal geometry of the complexes and determined the most energetically preferred arrangement of the ligands in the space of myosin subfragment-1 binding domain.
Сокращения
- C-798:
-
Tetrahydroxythiacalix[4]arene-tetrasulfosphonate
- C-800:
-
Tetrahydroxythiacalix[4]arene-tetraphosphonate
- CNDO:
-
Complete Neglect of Differential Overlap (methods)
- K м :
-
Michaelis constant, the substrate concentration at which the reaction rate of the enzyme is half of the maximal velocity
- L50:
-
Lower 50-kDa domain of myosin
- LD50 :
-
Lethal dose is the amount of an ingested substance that kills 50% of a test sample
- MM2:
-
A class of force fields
- Myosin S1:
-
Myosin subfragment-1
- NASU:
-
National Academy of Science of Ukraine
- PDB:
-
Protein Data Bank
- P-loop:
-
Phosphate-binding loop of myosin
- RCSB:
-
Research Collaboratory for Structural Bioinformatics
- U50:
-
The upper 50-kDa domain of myosin
- V макс :
-
Maximal velocity of the enzyme
- V max, ATP :
-
V макс for ATP
Наноматериалы
- Оцинкованная сталь и оцинкованная сталь:в чем разница?
- Воздействие COVID-19 на производство
- Доставка наночастиц артесуната увеличивает противоопухолевую эффективность за счет активации митохондриал…
- Влияние отношения Li / Nb на получение и фотокаталитические характеристики соединений Li-Nb-O
- Положительные эффекты автоматизации сельского хозяйства
- Отключения электроэнергии:5 способов предотвратить негативные последствия
- Влияние радиальной тяги на центробежные насосы
- Эффект горения плазмы и кислорода/топлива
- Влияние общих легирующих элементов
- Каковы последствия текущего дефицита никеля?