


Тетраэдры ДНК, модифицированные Gint4.T, загруженные доксорубицином, подавляют пролиферацию клеток глиомы, воздействуя на PDGFRβ
Аннотация
Глиома - одна из самых смертоносных внутренних опухолей головного мозга из-за ее инвазивного роста. Эффект от лечения глиомы слабый из-за наличия гематоэнцефалического барьера и гематоэнцефалического барьера, а также недостаточного нацеливания лекарств. Тетраэдры ДНК (TDN) демонстрируют большой потенциал для доставки лекарств и могут быть новой терапевтической стратегией при глиоме. В этом исследовании мы использовали TDN для доставки доксорубицина (DOX) для терапии глиомы. Gint4.T, аптамер, который может распознавать рецептор тромбоцитарного фактора роста β на опухолевой клетке, был использован для модификации TDN (Apt-TDN) для адресной доставки лекарств. TDN были самоорганизованы путем одностадийного синтеза, который показал небольшой размер (10 нм) и отрицательный заряд. Тест фетальной бычьей сыворотки показал ее стабильность в качестве средства доставки лекарственного средства. Apt-TDN может эффективно использоваться ячейками U87MG. По сравнению с DOX и DOX @ TDN (TDN, нагруженный DOX), DOX @ Apt-TDN (TDN, модифицированный Gint4.T, нагруженный DOX) показал более высокую скорость апоптоза, более высокую остановку клеточного цикла и большую цитотоксичность по отношению к клеткам U87MG. В заключение, наши результаты показали, что DOX @ Apt-TDN представляет собой новую терапию с многообещающим клиническим применением для пациентов с глиомами.
Введение
Глиома, опухоль, происходящая из нейроэпителия, является наиболее распространенным внутричерепным злокачественным новообразованием. Почти 1/3 всех опухолей головного мозга представляют собой глиомы, и примерно 4/5 первичных злокачественных опухолей головного мозга - глиомы [1,2,3,4]. В настоящее время наиболее эффективным методом лечения глиомы является хирургическая резекция и одновременная послеоперационная химиолучевая терапия, но, к сожалению, прогноз для пациентов остается плохим. Традиционная химиотерапия глиомы не дает хороших результатов из-за плохого нацеливания на опухоль, осложнений из-за гематоэнцефалического барьера (BBB) и гематоэнцефалического барьера (BTB), а также недостаточного нацеливания лекарств. ГЭБ является наиболее важным фактором, препятствующим доставке почти всех макромолекул (включая лекарства и гены) в паренхиму мозга и их правильному функционированию. Лекарства, которые не могут проникать через ГЭБ, необходимо вводить в достаточно высоких дозах для достижения эффективной лечебной концентрации в интересующей области. Однако избыток лекарств может вызвать серьезные системные побочные эффекты и нежелательное накопление лекарств в непораженных тканях. Более того, существующие традиционные препараты против глиомы не обладают достаточными возможностями нацеливания [3, 4]. Наночастицы стали наиболее многообещающим инструментом для переноса лекарств. Из-за своего преимущества в размере наночастицы могут пересекать ГЭБ и оказывать противоопухолевый эффект. Кафа и др. [5] разработали химически функционализированные многостенные углеродные нанотрубки (f-MWNT), нацеленные на ANG, и подтвердили их способность пересекать BBB в экспериментах in vivo и in vitro. Однако эти наноматериалы могут распространяться в различных органах по всему телу или даже попадать в центральную нервную систему (ЦНС), где они могут вызывать нейротоксичность [6].
ДНК является идеальным материалом для создания наноструктур, поскольку ее сборка может точно контролироваться спариванием оснований Уотсона-Крика [7]. На сегодняшний день разработан и продемонстрирован ряд двумерных (2D) и трехмерных (3D) наноструктур ДНК [8,9,10]. Тетраэдрические наноструктуры ДНК (TDN) привлекли значительное внимание из-за их биосовместимости, стабильности, большого количества функционализированных сайтов модификации и низкой иммуногенности [11,12,13]. Turberfield et al. синтезировали тетраэдрические наноструктуры ДНК с высоким выходом методом одностадийного синтеза [14]. Walsh et al. обнаружили, что ДНК-тетраэдр, содержащий зонд нуклеиновой кислоты, может проникать в клетки млекопитающих без необходимости в реагенте для трансфекции [15]. Ли и др. продемонстрировали, что самоорганизующиеся тетраэдрические наночастицы могут быть использованы для адресной доставки миРНК in vivo [16]. TDN показали отличные перспективы применения в молекулярной диагностике, молекулярной доставке и таргетной лекарственной терапии. Они также широко используются при исследовании опухолей различных органов, например рака груди [17]. Точно так же TDN также использовались при изучении заболеваний нервной системы. Тиан и др. [18] использовали тетраэдры ДНК в качестве основы и модифицировали их с помощью ангиопепа-2 (ANG), чтобы успешно сконструировать нанозонд ANG-TDN, который может воздействовать на белок 1, связанный с рецептором липопротеинов низкой плотности (LRP-1), для нацеленной визуализации. Исследования показали, что ANG-TDN может пересекать BBB. Ma et al. [19] предположили, что тетраэдрические наноструктуры ДНК могут проникать в нервные стволовые клетки (НСК) без использования агентов трансфекции, где они способствуют миграции, пролиферации и дифференцировке нервных стволовых клеток [20] и имеют большой потенциал для восстановления и регенерации нервной ткани. . Поэтому мы исследовали возможность использования TDN в терапии глиомы и повышения его способности к нацеливанию.
Рецептор фактора роста тромбоцитов β (PDGFRβ) является важным членом семейства тирозиновых протеинкиназ, который участвует в клеточной пролиферации, миграции и ангиогенезе. Несколько исследований показали, что PDGFβ является многообещающей мишенью для противоопухолевой терапии из-за его роли в ангиогенезе [21, 22]. Аптамеры представляют собой короткие одноцепочечные олигонуклеотиды ДНК или РНК, которые производятся путем систематической эволюции лигандов методом экспоненциального обогащения (SELEX). Аптамеры подобны антителам, которые обладают высокой аффинностью и специфичностью по отношению к своим мишеням [23]. Благодаря своим уникальным свойствам аптамеры играют важную роль в адресной доставке химиотерапевтических агентов, миРНК и наночастиц, содержащих лекарственные средства. Gint4.T, аптамер РНК, который может специфически связывать PDGFRβ, также является специфическим антагонистом PDGFRβ [24]. Монако и др. [25] предположили, что аптамеры Gint4.T могут преодолевать гематоэнцефалический барьер (BBB) и специфически распознавать PDGFRβ. Конъюгированные с Gint4.T полимерные наночастицы (PNP) могут легко поглощаться клетками глиобластомы (GBM). В этом исследовании мы сообщаем о новой системе, содержащей лекарственные препараты, которая сочетает в себе аптамер Gint4.T и TDN. Модифицированный Gint4.T TDN (Apt-TDN), нагруженный DOX (DOX @ Apt-TDN), проявлял повышенное специфическое клеточное поглощение и цитотоксичность в отношении клеток U87MG.
Методы
Материалы
Все олигонуклеотиды ДНК и олигонуклеотиды 2’F-Py РНК были приобретены у Sangon Biotech (Шанхай, Китай), и все олигонуклеотидные последовательности перечислены в таблице 1. Раствор для окрашивания геля ДНК GelRed был приобретен у Sangon Biotech. И фетальная бычья сыворотка (FBS), и среда Игла, модифицированная Дульбекко (DMEM), были приобретены в Thermo Fisher (Нью-Йорк, США). Доксорубицин (DOX) был приобретен у Mengbio Technology (Чунцин, Китай). Клетки U87MG были приобретены из библиотеки клеток Шанхайской академии наук (Шанхай, Китай). DAPI был приобретен у компании Zhongshan Golden Bridge Biotechnology (Пекин, Китай).
Подготовка наноструктур ДНК
Для сборки тетраэдра ДНК (таблица 1) 2 мкл каждого олигонуклеотида (S1, S2, S3 и S4) добавляли к 42 мкл буфера TM (10 мМ Tris-HCl, 5 мМ MgCl 2 , pH =8). Затем раствор ДНК нагревали до 95 ° C в течение 5 мин и затем охлаждали до 4 ° C в течение 2 мин с помощью ПЦР-аппарата Bio-Rad (Калифорния, США) [26, 27]. Конечная концентрация TDN составляла 2 мкМ. TDN ’был подготовлен таким же образом, за исключением того, что S1 был заменен на S1’. Для синтеза Apt-TDN аптамер Gint4.T добавляли к TDN в равном молярном соотношении, и смесь инкубировали при 37 ° C в течение 60 мин. Перед синтезом аптамер был подвергнут короткой стадии денатурации-ренатурации (85 ° C в течение 5 минут, быстрое охлаждение в течение 2 минут и последующее нагревание до 37 ° C в течение 10 минут) [25].
Электрофорез в агарозном геле
Гель агарозы (3%) прогоняли в буфере 0,5 × TEB при 100 В в течение 30 мин. Температуру устройства для электрофореза поддерживали на уровне 0 ° C, помещая устройство в ледяную баню. Перед электрофорезом к агарозному гелю добавляли GelRed для окрашивания цепей ДНК. Когда процесс был завершен, для получения изображения геля использовали флуоресцентный сканер Bio-Rad (Калифорния, США).
Динамическое рассеяние света
Malvern Zetasizer ZS90 (Малверн, Великобритания) использовали для измерения гидродинамического размера и дзета-потенциала TDN. Всего 1 мл TDN (100 нМ) был подвергнут анализу динамического светорассеяния (DLS).
Получение изображений методом атомно-силовой микроскопии
TDN разбавляли до 100 нМ буфером TM (буфер Tris-HCl, содержащий MgCl 2 ). Затем 10 мкл каждого образца TDN добавляли к свежеотщепленной слюде и инкубировали в течение 10 мин. Затем образцы были отображены на приборе для атомно-силовой микроскопии (АСМ) в режиме переменного тока (Agilent 5500, США).
Измерение способности загружать лекарство подготовленного TDN
Доксорубицин растворяли в деионизированной воде, чтобы получить раствор для хранения 500 мкМ. Доксорубицин в различных концентрациях (от 1 до 20 мкМ) смешивали с TDN (100 нМ) или Apt-TDN (100 нМ) в течение 6 ч при комнатной температуре (24–26 ° C). Затем смешанные растворы центрифугировали при 12000 × g в течение 10 мин для получения ТДН, нагруженного лекарством. Затем 50 мкл супернатантов удаляли и смешивали с PBS в соотношении 1:1. Микропланшетный ридер Varioskan LUX (Калифорния, США) использовали для измерения интенсивности флуоресценции доксорубицина ( λ бывший =480 нм и λ em =590 нм) для определения количества доксорубицина в супернатантах [28]. Концентрацию доксорубицина, загруженного в TDN, рассчитывали по стандартной кривой и интенсивности флуоресценции. Мы также смешали доксорубицин с TDN в увеличивающихся молярных соотношениях.
Стабильность сыворотки TDN in vitro
TDN смешивали с полной средой и инкубировали при 37 ° C в течение 0, 2, 4, 6, 8, 10, 12 или 24 часов. Растворы TDN смешивали с FBS в соотношении 1:1 и инкубировали при 37 ° C в течение 1, 3, 5 или 7 часов. После инкубации смеси обрабатывали в 3% агарозном геле.
Цитотоксичность TDN in vitro
Чтобы определить цитотоксичность TDN, клетки U87MG в концентрации 1 × 10 4 клеток / лунку высевали на 96-луночный планшет. Среду для культивирования клеток удаляли, добавляли свежую среду, содержащую 0-500 нМ TDN, и инкубировали еще 24 и 48 часов после инкубации в течение ночи. Затем в каждую лунку добавляли 10 мкл раствора CCK-8 и смесь инкубировали в течение 1 ч. Затем измеряли оптическую плотность при 450 нм с помощью считывающего устройства для микропланшетов.
Флуоресцентная визуализация
Поглощение DOX и TDN клетками изучали с помощью флуоресцентной микроскопии (Olympus, Токио, Япония). Клетки U87MG высевали на покровные стекла в 24-луночные планшеты со средой, содержащей 10% инактивированной нагреванием фетальной бычьей сыворотки и 1% пенициллина и стрептомицина, и выращивали в течение не менее 1 дня при 37 ° C в увлажненной атмосфере с 5% CO 2 пока клетки не достигли слияния не менее 75%. После инкубации питательные среды удаляли. Добавляли полную среду, содержащую 100 нМ Cy3-TDN и Cy3-Apt-TDN, и инкубировали в течение 3 часов. TDN и Apt-TDN были помечены Cy3 для обнаружения межклеточного поглощения наночастиц. Для оценки клеточного поглощения DOX, DOX (DOX 2 мкМ), DOX @ TDN (DOX 2 мкМ) и DOX @ Apt-TDN (DOX 2 мкМ) добавляли к клеткам U87MG и инкубировали в течение 3 часов. Через 3 часа обработки клетки фиксировали 4% параформальдегидом в течение 20 минут в темноте и затем окрашивали 4 ', 6-диамидино-2-фенилиндолом (DAPI) в течение 5 минут. Клетки трижды промывали PBS и наблюдали под флуоресцентным микроскопом.
Проточная цитометрия
Всего 1 × 10 6 Клетки U87MG имплантировали в 6-луночные планшеты. После инкубации в течение ночи культуральные среды удаляли, добавляли среды с добавлением 100 нМ Cy3-TDN, 100 нМ Cy3-Apt-TDN или 100 нМ Cy3-Apt-TDN + 1 мкМ свободного Apt и инкубировали в течение 3 часов. Затем клетки фиксировали 4% параформальдегидом в течение 20 минут и использовали проточную цитометрию для анализа процентного содержания Cy-3-положительных клеток.
Клеточный цикл и апоптоз
После обработки DOX, DOX @ TDN или DOX @ Apt-TDN в течение 24 часов, 5 × 10 5 клетки собирали и фиксировали в 75% ледяном этаноле в течение ночи. Затем клетки инкубировали с РНКазой и иодидом пропидия в течение 30 мин при 37 ° C в темноте. Клеточный цикл исследовали методом проточной цитометрии. Кроме того, после различных обработок клетки окрашивали аннексином V-FITC / DAPI и исследовали ранний апоптоз.
Анализы CCK-8
Чтобы определить жизнеспособность клеток, клетки U87MG (5 × 10 3 ) высевали на 96-луночные планшеты со 100 мкл среды и культивировали в течение ночи при 37 ° C в атмосфере, содержащей 5% CO 2 . Затем среду удаляли и добавляли свежую среду, содержащую DOX, DOX-TDN или DOX-Apt-TDN. После 24 часов инкубации добавляли 10 мкл раствора CCK-8, и клетки культивировали еще 1 час. Считывающее устройство для микропланшетов использовали для измерения оптической плотности при 450 нм.
Статистический анализ
Все эксперименты в этом исследовании были выполнены в трех экземплярах, и все данные представлены как среднее значение с его стандартным отклонением (среднее ± стандартное отклонение). Статистический анализ проводили с помощью программы SPSS 24.0 (IBM, США). Значимые различия были определены с использованием t Стьюдента. тест, с P <0,05, что указывает на значительные различия между группами.
Результаты
Синтез и характеристика TDN и Apt-TDN
TDN самособирался из четырех олигонуклеотидов (таблица 1) посредством одностадийного синтеза, как сообщалось ранее [18, 29]. Нацеленный на опухоль аптамер Gint4.T использовали для модификации TDN посредством спаривания оснований Уотсона-Крика. Тетраэдр ДНК содержит четыре грани, каждая из которых образована одним олигонуклеотидом. Таким образом, четыре олигонуклеотида гибридизуются друг с другом, образуя тетраэдр ДНК (рис. 1а). Анализ гель-электрофореза показал единственную заметную полосу на дорожках 4 и 5, предполагая, что TDN и Apt-TDN были успешно сконструированы. Подвижность Apt-TDN была снижена по сравнению с подвижностью TDN, что свидетельствует о том, что аптамер Gint4.T успешно модифицировал TDN.
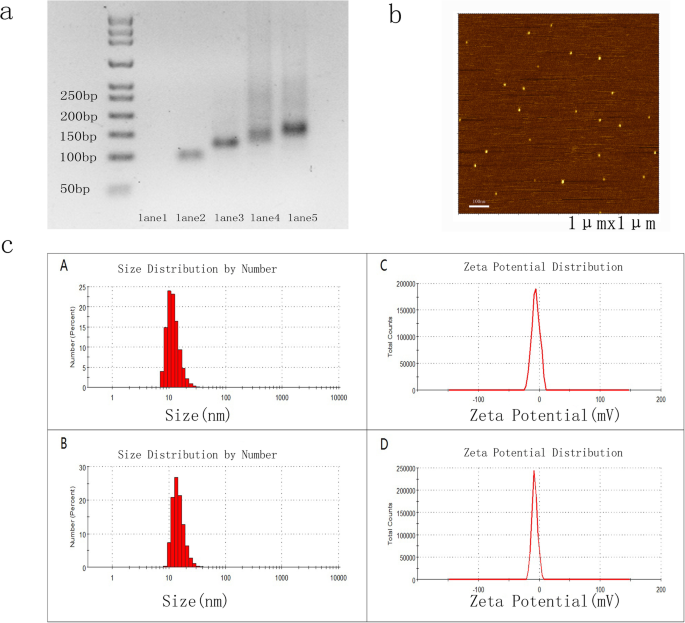
а Синтез ДНК-тетраэдра и Gint4.T-TDN. Дорожка 1:S1; дорожка 2:S1 + S2; дорожка 3:S1 + S2 + S3; полоса 4:S1 + S2 + S3 + S4 (TDN); полоса 5:TDN, смешанный с Apt-tail (Gint4.T); Апт-ТДН. Дорожка 1 не была видна, потому что красители нуклеиновых кислот не могут должным образом окрашивать однонитевую ДНК. б Изображения АСМ показали, что высота TDN и Apt-TDN составляла ~ 2 нм. c Определение размера частиц и дзета-потенциала TDN и Apt-TDN с помощью динамического рассеяния света (DLS). Средний размер частиц TDN и Apt-TDN составлял 10,10 нм (A) и 13,54 нм (B) соответственно. Средние дзета-потенциалы TDN и Apt-TDN составляли -5,69 мВ (C) и -7,3 мВ (D), соответственно
Размеры TDN и Apt-TDN определялись методами DLS и AFM. TDN и Apt-TDN показали размеры частиц 10,1 нм и 13,5 нм, соответственно, что отражает добавление лиганда Gint4.T (рис. 1c (A), (B)). Поскольку гидродинамический диаметр включает молекулы воды, частицы были больше, чем их теоретические размеры. Высота как TDN, так и Apt-TDN, определенная с помощью АСМ-изображений, составляла ~ 2 нм (рис. 1b), что указывает на то, что модификация аптамера не изменяет трехмерную структуру. Средние дзета-потенциалы TDN и Apt-TDN составляли -5,69 мВ (C) и -7,3 мВ (D), соответственно (рис. 1c (C) (D)). Основываясь на этих параметрах, мы сделали вывод, что TDN и Apt-TDN были успешно собраны.
Стабильность и цитотоксичность TDN in vitro
Анализ гель-электрофореза показал, что TDN оставался интактным при инкубации в полной среде в течение 24 часов при 37 ° C (рис. 2a (A)). Кроме того, когда концентрация фетальной бычьей сыворотки была увеличена до 50%, TDN оставался стабильным в течение как минимум 7 часов (рис. 2a (B)), что согласуется с предыдущими сообщениями [18, 26]. Для определения цитотоксичности наноструктуры был использован анализ CCK-8 для оценки жизнеспособности клеток U87MG после обработки TDN в нескольких концентрациях. Как показано на рис. 2b, не наблюдалось значительной цитотоксичности в клетках U87MG, обработанных TDN в концентрации 0–500 нМ в течение 24 и 48 часов. Следовательно, наночастицы ДНК могут использоваться в качестве стабильных и биологических носителей для доставки лекарств.
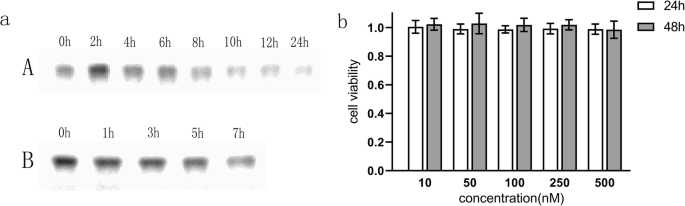
а Гель-электрофорез показал, что TDN оставался стабильным в течение 24 часов в полной среде при 37 ° C (A); TDN оставался стабильным в течение 7 часов, когда концентрация фетальной бычьей сыворотки была увеличена до 50% (B). б Клетки U87MG совместно культивировали с TDN в различных концентрациях (10–500 нМ) в течение 24 и 48 часов. Анализ CCK-8 показал, что активность клеток U87MG не изменилась, что указывает на биобезопасность TDN
Вместимость лекарств для TDN и Apt-TDN
Доксорубицин - это химиотерапевтический препарат широкого спектра действия, который может интеркалировать в двойные цепи ДНК. Мы рассчитали стандартную кривую доксорубицина (рис. 3a), а затем исследовали интеркаляцию доксорубицина в TDN. Количество интеркалированного доксорубицина в TDN и Apt-TDN постепенно увеличивалось с увеличением концентрации доксорубицина. Когда концентрация доксорубицина составляла 14 мкМ, количество доксорубицина, интеркалированного в TDN и Apt-TDN, достигало пика при 5,5 мкМ и 6,0 мкМ соответственно, а затем выходило на плато (фиг. 3b), указывая на то, что цепи ДНК были полностью заняты. Между тем, мы смешивали доксорубицин с TDN в увеличивающихся молярных соотношениях. Спектр флуоресценции доксорубицина сканировали для анализа. Как показано на фиг. 3c, спектр флуоресценции доксорубицина гасили доксорубицином при молярном соотношении 0,05. На основании этих результатов мы пришли к выводу, что примерно 55 молекул доксорубицина содержались в одном TDN, в то время как 60 молекул содержались в одном Apt-TDN.
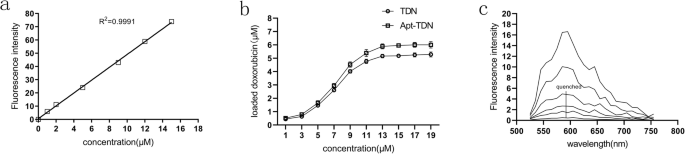
а Стандартная кривая концентраций DOX в буфере PBS; λex =480 нм и λem =590 нм. Количество DOX, переносимое TDN и Apt-TDN. б DOX интеркалирован в двухцепочечную ДНК TDN и Apt-TDN. Когда концентрация DOX достигает 14 мкМ, а концентрация интеркалированного DOX в TDN и Apt-TDN достигает пика на уровне 5,5 мкМ и 6,0 мкМ, соответственно, один тетраэдр ДНК может нести 55 молекул Dox, в то время как один тетраэдр ДНК, модифицированный аптамером, несет 60 DOX. молекулы. c Спектры флуоресценции DOX в супернатанте. Доксорубицин смешивали с TDN в возрастающих молярных соотношениях (0, 0,0005, 0,001, 0,005, 0,01 и 0,05 сверху вниз). Когда молярное соотношение составляло 1:20, флуоресценция гасилась
Целевое использование сотовой связи Apt-TDN
ДНК представляет собой отрицательно заряженную макромолекулу, что затрудняет ее проникновение в отрицательно заряженную клеточную мембрану. Обычно отдельные молекулы ДНК должны получать доступ к клеткам с помощью реагента для трансфекции. Здесь мы обозначили TDN и Apt-TDN с помощью Cy3 для мониторинга внутриклеточного поглощения наночастиц. После инкубации с клетками U87MG в течение 3 часов в цитоплазме клетки появился красный сигнал флуоресценции Cy3, что указывает на то, что TDN связывает цитомембрану и попадает в клетку без помощи агентов трансфекции (рис. 4a). Apt-TDN показал более высокую красную флуоресценцию, что свидетельствует о том, что присутствие аптамера Gint4.T значительно увеличивает поглощение тетраэдра ДНК клетками U87MG. Однако при добавлении свободного аптамера флуоресценция Су3 снижалась до уровня, наблюдаемого для одного TDN. Из-за конкурентного ингибирования свободным аптамером мы делаем вывод, что аптамер в TDN не может способствовать захвату TDN. Основываясь на этом конкурентном ингибировании, мы доказали, что Apt-TDN может нацеливаться на клетки U87MG. Проточная цитометрия дополнительно подтвердила, что процент Cy3-положительных клеток U87MG был выше в группе Apt-TDN, чем в группе TDN. Free Apt снизил процент Cy3-положительных клеток U87MG в группе Apt-TDN (рис. 4b).
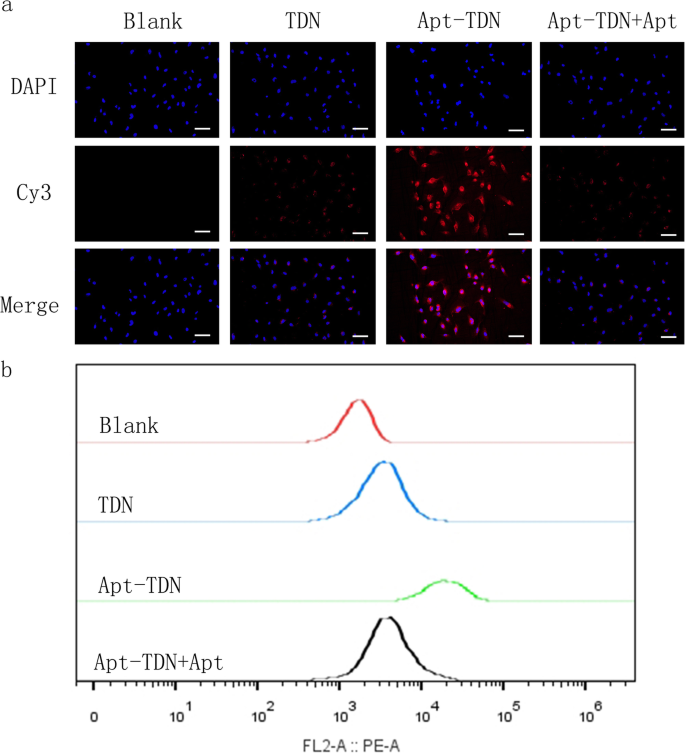
а Поглощение клетками U87MG TDN и Apt-TDN (TDN-Gint4.T). TDN поступал в клетки U87MG напрямую без агентов трансфекции, и поглощение Apt-TDN (связанного с аптамером Gint4.T) было значительно увеличено и конкурентно ингибировано свободным Apt (Gint4.T), что указывает на то, что аптамер Gint4.T играет роль значительная роль в нацеливании на клетки. Масштабная линейка представляет 50 мкм. б Кривые проточной цитометрии показывают внутриклеточное поглощение TDN, Apt-TDN и Apt-TDN + Apt после инкубации в течение 3 часов
Использование сотовой связи DOX @ TDN и DOX @ Apt-TDN
Мы использовали характеристический спектр флуоресценции доксорубицина для оценки эффективности поглощения лекарственного средства. Через 3 часа обработки внутриклеточный доксорубицин был визуализирован с помощью флуоресцентной микроскопии (рис. 5а). Свободный доксорубицин мог проникать в клетки U87MG и располагался в ядре. При добавлении DOX @ TDN флуоресценция была выше, чем у свободного доксорубицина. Этот результат свидетельствует о том, что наночастицы ДНК увеличивают поглощение доксорубицина клетками. Когда был добавлен DOX @ Apt-TDN, красный сигнал в ядре был даже выше, чем у клеток, к которым был добавлен DOX @ TDN. Полуколичественный анализ внутриклеточного поглощения DOX дополнительно подтвердил, что Apt-TDN способствовал более чем двукратному увеличению внутриклеточного поглощения DOX по сравнению с таковым для одного лекарственного средства. Мы предполагаем, что это связано с Gin4.T-специфическим связыванием с рецепторами, после чего в клетку может попасть больше наночастиц. После переваривания в лизосомах доксорубицин может высвобождаться в цитоплазму и функционировать в ядре.
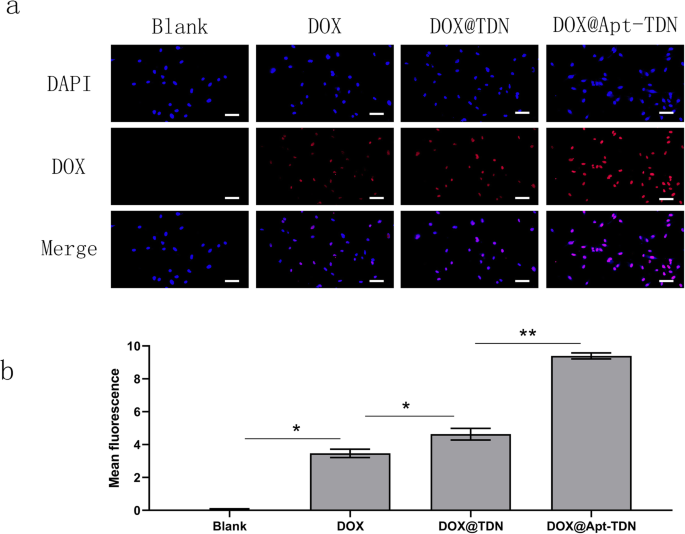
а Поглощение клетками DOX, DOX @ TDN и DOX @ Apt-TDN. Модифицированный аптамером Gint4.T, Apt-TDN может доставлять больше доксорубицина в клетки U87MG, чем TDN. Кроме того, TDN может переносить в клетки больше лекарства, чем одно лекарство. Масштабные линейки указывают 50 мкм. б Полуколичественный анализ интенсивности флуоресценции доксорубицина с обработкой PBS, DOX, DOX @ TDN и DOX @ Apt-TDN (по сравнению с контрольным образцом:* p <0,05, ** p <0,01)
Цитотоксичность DOX, DOX @ TDN и DOX @ Apt-TDN
Для исследования цитотоксичности три группы клеток U87MG обрабатывали разными концентрациями доксорубицина (рис. 6а). IC 50 значения для доксорубицина составляли 13,39 мкМ для обработки DOX, 7,826 мкМ для обработки DOX @ TDN и 4,205 мкМ для обработки DOX @ Apt-TDN. Среди трех обработок DOX @ Apt-TDN показал наивысшую цитотоксичность через 24 часа, что указывает на специфичность Apt-TDN для клеток U87MG. Через 24 часа клетки в группах DOX, DOX @ TDN и DOX @ Apt-TDN собирали и использовали для исследования раннего апоптоза. Наши данные показали, что скорость раннего апоптоза была выше в группе DOX @ Apt-TDN, чем в двух других группах (рис. 6b). Более того, доля клеток в фазе G0 / G1 была увеличена в группе DOX-Apt-TDN по сравнению с группами DOX и DOX-TDN ( p <0,01), а соотношение клеток в фазе S было уменьшено в группе DOX-Apt-TDN ( p <0,01) (рис. 6в). Процент клеток на фазе G2 не изменился (данные не показаны).
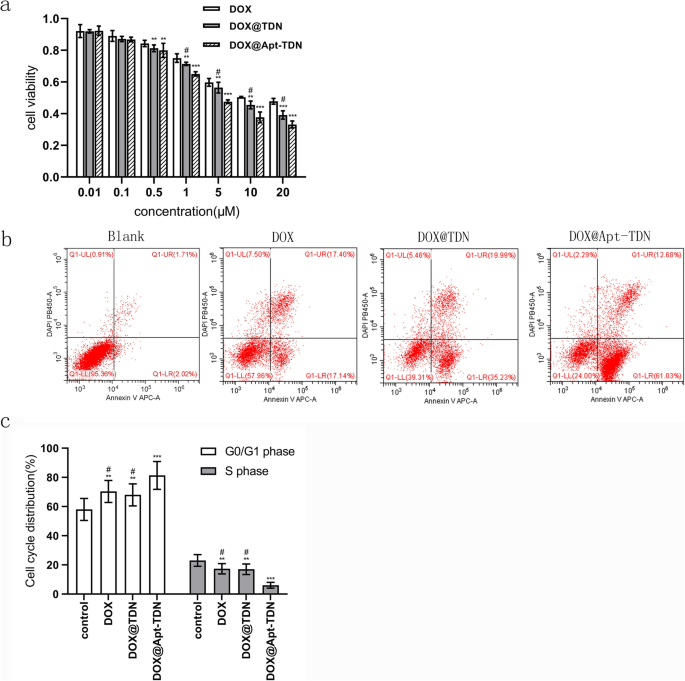
а Цитотоксичность DOX, DOX @ TDN и DOX @ Apt-TDN в различных концентрациях. Скорость ингибирования клеток U87MG значительно увеличивалась с увеличением концентрации DOX, но группы DOX @ TDN и DOX @ Apt-TDN проявляли значительно повышенную цитотоксичность по сравнению с группой DOX. Уровень ингибирования клеток в группе DOX @ Apt-TDN также был значительно выше, чем в группе DOX @ TDN (по сравнению с DOX, ** p < 0,05; по сравнению с DOX, *** p <0,01; по сравнению с DOX @ Apt-TDN, # p <0,05). б Апоптоз клеток U87MG после инкубации с PBS, DOX, DOX @ TDN и DOX @ Apt-TDN в течение 24 часов. c Гистограммы проточной цитометрии клеточного цикла U87MG после инкубации с PBS, DOX, DOX @ TDN и DOX @ Apt-TDN в течение 24 часов (по сравнению с контролем, ** p < 0,05; по сравнению с контролем, *** p <0,01; по сравнению с DOX @ Apt-TDN, # p <0,01)
Обсуждение
С помощью экспериментов in vitro или in vivo необходимо определять стабильность лекарственного средства и его носителя. После сборки TDN его стабильность сначала определяли in vitro. Это исследование показало, что трехмерная структура наночастиц ДНК может улучшить их стабильность в сыворотке за счет ингибирования связывания ферментов. Биологическая безопасность соответствующей наноструктуры - важнейшее требование для ее применения. Не наблюдалось значительной цитотоксичности в клетках U87MG, совместно культивируемых с различными концентрациями TDN в течение 24 или 48 часов. Ни одна из последовательностей ДНК, использованных в этом исследовании, не кодирует какую-либо генетическую информацию, и в тесте на цитотоксичность не сообщалось о побочных эффектах. Таким образом, TDN может служить безопасным и стабильным носителем лекарства.
Эффективность нацеливания модифицированной аптамером наноструктуры имеет решающее значение для селективной доставки лекарств к раковым клеткам. В отличие от антител, аптамеры химически стабильны, недороги и могут производиться массово. Кроме того, в отличие от других материалов, аптамеры могут легко связывать тетраэдр ДНК, используя принцип комплементарности оснований. Таким образом, комбинация аптамеров и тетраэдров ДНК закладывает основу для адресной доставки лекарств и лечения следующего поколения. Наши результаты показали, что TDN может проникать в клетки без агентов трансфекции, что аналогично результатам Walsh et al. и Ma et al. [15, 19]. По сравнению с TDN, поглощение Apt-TDN клетками U87MG было значительно увеличено. После добавления свободного аптамера это увеличение исчезло. Этот результат предполагает, что Gint4.T является специфическим антагонистом PDGFRβ [25]. Наше исследование продемонстрировало, что аптамер Gint4.T может нацеливаться на клетки U87MG из-за высокой экспрессии PDGGRβ на поверхности клеток глиомы. Camorani et al. [24] также показали, что аптамер Gint4.T может нацеливаться на опухолевые клетки, специфически взаимодействуя с внеклеточным доменом PDGFRβ. Преимущества нацеливания на аптамер Gint4.T были до некоторой степени продемонстрированы исследованиями in vitro, но для подтверждения этих результатов необходимы дальнейшие испытания in vivo. Это исследование также подтвердило, что DOX @ Apt-TDN более цитотоксичен, чем DOX или DOX @ TDN, что, вероятно, связано с двумя факторами. Во-первых, аптамер Gint4.T может специфически связывать внеклеточные домены PDGFR, блокируя пролиферацию опухолевых клеток и подавляя рост опухолевых клеток [24]. Это согласуется с нашими экспериментальными результатами. По сравнению с контрольной группой, изменения клеточного цикла U87MG после обработки DOX, DOX @ TDN и DOX @ Apt-TDN значительно увеличились в опухолевых клетках фазы G0 / G1, уменьшились в клетках фазы S и заблокировали опухолевые клетки в G0. Фаза / G1, что указывает на то, что они могут ингибировать клеточный цикл клеток U87MG и подавлять их пролиферацию. По сравнению с группами DOX и DOX @ TDN, группа DOX @ Apt-TDN имела значительно более сильную способность ингибировать пролиферацию клеток U87MG. Кроме того, Gint4.T специфически связывает опухолевые клетки и усиливает нацеливание на клетки комплекса DOX @ Apt-TDN. Таким образом, мы можем повысить эффективность нацеливания на лекарства и снизить дозу системного введения противоопухолевых препаратов, чтобы предотвратить их системные побочные эффекты.
Выводы
Модификация с помощью Gint4.T увеличивала специфичность и эффективность TDN при лечении глиомы за счет нацеливания на PDGFRβ, который в больших количествах экспрессируется на клетках глиомы. Кроме того, при загрузке Apt-TDN он может значительно усиливать антиглиомный эффект DOX. Следовательно, DOX @ Apt-TDN может служить многообещающей терапевтической стратегией против глиомы для пациентов. Недостатком этого исследования является то, что оно было проверено только in vitro. В более позднем исследовании мы продолжим исследования на животных моделях.
Доступность данных и материалов
Соответствующие данные включены в статью.
Сокращения
- BBB:
-
Гематоэнцефалический барьер
- BTB:
-
Барьер опухоли крови
- TDN:
-
Тетраэдрические наноструктуры ДНК / Тетраэдры ДНК
- DOX:
-
Доксорубицин
- PDGFRβ:
-
Рецептор фактора роста тромбоцитов β
- Apt-TDN:
-
Gint4.T-модифицированный TDN
- DOX @ TDN:
-
В TDN загружен DOX
- DOX @ Apt-TDN:
-
Модифицированный Gint4.T TDN, загруженный с помощью DOX
- CNS:
-
Центральная нервная система
- ANG:
-
Angiopep-2
- LRP-1:
-
Low-density lipoprotein receptor-related protein 1
- SELEX:
-
Exponential enrichment
- PNPs:
-
Polymeric nanoparticles
- GBM:
-
Glioblastoma
- FBS:
-
Foetal bovine serum
- DMEM:
-
Модифицированный носитель Орла, созданный Дульбекко
- DLS:
-
Динамическое рассеяние света
- AFM:
-
Атомно-силовая микроскопия
- CCK-8:
-
Cell Counting Kit-8
- Cy3:
-
Sulfo-Cyanine3
- DAPI:
-
4′,6-Diamidino-2-phenylindole
Наноматериалы
- Оптимизация эффективности передачи с помощью роботизированной ячейки
- Нацеливание на эндотелиальные клетки с помощью многофункциональных наночастиц GaN / Fe
- Сверхчувствительный биосенсор для обнаружения ДНК холерного вибриона с помощью композитных наносфер полист…
- Стимуляция роста клеток SH-SY5Y наночастицами золота, модифицированными 6-меркаптопурином и проникающим в нейро…
- Исследование солнечного элемента из кристаллического кремния с черным слоем кремния на задней панели
- Нанокомпозиты на основе оксида графена, украшенные наночастицами серебра в качестве антибактериального аге…
- Изготовление монокристаллического кремниевого солнечного элемента с эффективностью 20,19% с микроструктурой …
- Влияние фуллерена C60 на взаимодействие дифенил-N- (трихлорацетил) амидофосфата с ДНК in Silico и его цитотоксическ…
- Улучшенные фотоэлектрические свойства в солнечном элементе с плоским гетеропереходом Sb2S3 с подходом быстрог…
- Оптимизация вашего сварочного приложения с помощью специального сварочного модуля RW950