


Длинная некодирующая РНК MALAT1 / microRNA-143 / VEGFA Сигнальная ось модулирует внутричерепную аневризму, вызванную повреждением эндотелия сосудов
Аннотация
Роль некоторых длинных некодирующих РНК (lncRNAs) во внутричерепной аневризме (IA) изучалась во многих исследованиях. Целью данного исследования является выяснение механизма передачи сигнала транскрипта 1 аденокарциномы легких, связанного с метастазами днРНК (MALAT1) / микроРНК-143 (miR-143) / фактора роста эндотелия сосудов (VEGFA), в сосудистой эндотелиальной оси, вызванной повреждением эндотелия. . Выявлена экспрессия MALAT1, miR-143 и VEGFA в тканях IA и нормальных тканях артерий. Были обнаружены матриксная металлопротеиназа 9 (MMP-9) в тканях, фактор фон Виллебранда (vWF) в сыворотке и тканях и эндотелин-1 (ET-1) в сыворотке. Смоделированным крысам IA вводили молчащий или сверхэкспрессированный MALAT1 для обнаружения повреждения эндотелия сосудов. Эндотелиальные клетки сосудов от пациентов с IA были извлечены и трансфицированы подавленным или сверхэкспрессированным MALAT1, чтобы проверить влияние MALAT1 на жизнеспособность клеток и апоптоз. Связи между MALAT1, miR-143 и VEGFA были проверены с помощью онлайн-предсказания, активности люциферазы и анализа РНК-pull down. Сверхэкспрессия MALAT1 и VEGFA и плохая экспрессия miR-143 были обнаружены в тканях IA. Подавление MALAT1 подавляло артериальное давление, экспрессию ET-1, vWF и MMP-9, а также индекс апоптоза эндотелиальных клеток сосудов крыс с IA. Подавленный MALAT1 ингибировал апоптоз и способствовал жизнеспособности эндотелиальных клеток сосудов в IA. MALAT1, связанный с miR-143, и miR-143 нацелен на VEGFA. Это исследование предполагает, что MALAT1 повышает экспрессию VEGFA за счет конкурентного связывания с miR-143, тем самым усиливая апоптоз и снижая жизнеспособность эндотелиальных клеток сосудов в IA.
Введение
Внутричерепная аневризма (IA), также известная как церебральная аневризма, вызывается повышением внутричерепного давления, вызванным локальным аномальным расширением полости или стенки артерии головного мозга [1]. IA - это тяжелое заболевание с высокой смертностью и заболеваемостью, а уровень распространенности составляет около 1–3% в общей популяции [2]. Основными клиническими признаками ИА являются церебральный вазоспазм, спонтанное кровоизлияние в мозг и паралич глазодвигательного нерва [3]. На сегодняшний день общие факторы риска, приводящие к возникновению и развитию ИА, включают гемодинамические нарушения, ген, старение, инфекции и врожденные факторы [4]. Основные клинические методы лечения, в основном хирургическое клипирование и / или эндоваскулярная намотка, предназначены для предотвращения разрыва аневризмы [5]. Однако подробный механизм, лежащий в основе ВА, еще предстоит выяснить, что отражает острую потребность в более эффективных методах управления ВА.
Длинные некодирующие РНК (днРНК) длиннее 200 нуклеотидов, которые принадлежат к разновидностям некодирующих РНК [6]. Сообщается, что экспрессия lncRNA, связанная с остановкой роста lncRNA 1 в IA подавляется транскриптом 1 аденокарциномы ancung (MALAT1) - высокообогащенной и широко экспрессируемой lncRNA, длина которой составляет около 8000 нт [7]. Было зарегистрировано, что MALAT1 модулирует дисфункцию гладких мышц при аневризме грудной аорты [8]. Кроме того, исследование представило аномальную экспрессию днРНК и матричную РНК (мРНК) в IA, а сети коэкспрессии днРНК-мРНК предоставляют ключи к разгадке патогенеза IA [9]. MALAT1, как полагают, ускоряет экспрессию остериксов и модулирует остеогенную дифференцировку посредством нацеливания на экспрессию miRNA-143 (miR-143) в мезенхимальных стволовых клетках костного мозга человека [10]. Исследование также представило роль кластера miR-143/145 в обращении регуляции krüppel-подобного фактора 5 в гладкомышечных клетках и его сократимости и пролиферации в IA [11]. Согласно Feng et al., Подавление уровня miR-143/145 и более высокого уровня матриксной металлопротеиназы-9 (MMP-9) во время циркуляции может быть связано с образованием и разрывом IA [12]. Анализ показал, что наиболее неконтролируемые miRNA (miR-143 и miR-145) являются общими для генов-мишеней, являющихся сигнальными путями, таких как фактор роста эндотелия сосудов (VEGF) и других генов, регулирующих движение или адгезию клеток [13]. . Исследование показало прогностическую важность вариаций фактора роста эндотелия сосудов А (VEGFA) при ИА [14]. Следовательно, мы стремились оценить механизм сигнальной оси MALAT1 / miR-143 / VEGFA в IA, вызванной повреждением эндотелия сосудов.
Материалы и методы
Этика
Исследование было одобрено институциональным наблюдательным советом Первой дочерней больницы USTC, Отделением наук о жизни и медицине, Научно-технологический университет Китая, и проводилось в соответствии с принципами Хельсинкской декларации. В этом исследовании участники предоставили письменное информированное согласие. Все эксперименты на животных проводились в соответствии с Руководством по уходу и использованию лабораторных животных Национального института здоровья. Протокол был разрешен Комитетом по этике экспериментов на животных Первой дочерней больницы USTC, Отделения наук о жизни и медицины, Научно-технологический университет Китая.
Темы исследования
Для наших экспериментов были отобраны 20 пациентов IA (группа IA), которым был поставлен диагноз визуализирующим исследованием и которым были сделаны нейрохирургические вырезки в Первой дочерней больнице USTC, Отделения наук о жизни и медицине Китайского университета науки и технологий. Всего было 11 мужчин и 9 женщин в возрасте 43,27 ± 6,25 года. Были взяты образцы тканей IA. Между тем, у 20 пациентов (контрольная группа) с височной эпилепсией, вызванной миндалевидным телом и склерозом гиппокампа, были резецированы сосудистые ткани височной кортикальной артерии со стороны височного полюса. Резецированные ткани после операции были исследованы как нормальные артериальные ткани по гистопатологическому исследованию, в них было 13 мужчин и 7 женщин в возрасте 44,18 ± 5,91 года. Не было обнаружено значительных различий по полу и возрасту между группой IA и контрольной группой (обе P > 0,05). Образцы венозной крови (2 пробирки) были взяты у всех испытуемых натощак в одно и то же время утром перед операцией.
Создание крысиных моделей IA
Шестьдесят крыс-самцов Sprague-Dawley чистой чистоты (Hunan SJA Laboratory Animal Co., Ltd., Хунань, Китай), весом от 200 до 250 г, выращивали в течение 7 дней (25 ± 2 ° C, относительная влажность 65-70). %, 12 ч светового и темного цикла, бесплатная вода и прием пищи). Крыс сбрасывали с эластазой поджелудочной железы свиньи на наружную сонную артерию и вокруг стенки бифуркационной артерии. Наружную сонную артерию лигировали двумя хирургическими линиями на ветви внешней сонной артерии около 1,5 мм. Внешнюю сонную артерию разрезали между двумя линиями, чтобы сформировать внутреннюю аневризму сонной артерии в слепом сегменте наружной сонной артерии. Крыс кормили 1% физиологическим раствором в течение 1 недели после операции. Церебральная ангиография была выполнена через 1 месяц, и было замечено образование аневризмы.
После создания моделей крыс IA, 50 крыс были случайным образом распределены в пустую группу ( n =10, моделируемым крысам вводили стереотаксическую инъекцию 100 мкл фосфатно-солевого буфера (PBS) один раз в день), группа коротких шпилечных РНК (sh) -отрицательный контроль (NC) ( n =10, моделируемым крысам вводили стереотаксическую инъекцию 100 мкл sh-MALAT1 NC один раз в день), группа sh-MALAT1 ( n =10, моделируемым крысам вводили стереотаксическую инъекцию 100 мкл плазмиды sh-MALAT1 один раз в день), группа сверхэкспрессии (Oe) -NC ( n =10, моделируемым крысам вводили стереотаксическую инъекцию 100 мкл плазмиды Oe-MALAT1 NC один раз в день), а группе Oe-MALAT1 ( n =10, моделированным крысам вводили стереотаксическую инъекцию 100 мкл плазмиды Oe-MALAT1 один раз в день) [15]. Вышеупомянутые плазмиды были созданы Shanghai Genechem Co., Ltd. (Шанхай, Китай).
Тест артериального давления у крыс
Артериальное давление в хвостовой артерии крыс измеряли на 1-й, 4-й и 12-й неделях после операции. Перед измерением артериального давления крыс на некоторое время помещали в устройство для нагрева с постоянной температурой, чтобы предотвратить нарушение внешней температуры. Во-вторых, крыс в течение нескольких минут держали в тишине в специальной клетке, чтобы предотвратить вмешательство в их жизнедеятельность. Если артериальное давление сильно варьировалось, его определяли дважды или трижды в разное время, чтобы получить среднее значение.
Сбор аневризмы ткани
Через 3 месяца крыс анестезировали 1% пентобарбиталом натрия (40 мг / кг) путем внутрибрюшинной инъекции для получения образцов крови из вен. Крыс умерщвляли, открывали грудную клетку, интубировали левый желудочек в аорту и разрезали полую полую часть для выпуска крови. Между тем, 30 мл физиологического раствора, содержащего гепарин натрия, использовали для быстрой сердечной перфузии для промывания крови, а затем в мозг вводили 10 мл 10% полиформальдегида / 0,1 М PBS (pH 7,4). После перфузии и фиксации мозг крысы вскрывали. Артериальное кровообращение в основании черепа тщательно наблюдалось, ткань аневризмы была отделена, и изменения аневризмы наблюдались под микроскопом.
Иммуноферментный анализ (ELISA)
Индексы, связанные с сывороткой, тестировали с помощью набора для ELISA. Собранные образцы крови помещали в термостат 37 ° C на 1 час и центрифугировали при 3000 об / мин в течение 10 минут. Обнаружение экспрессии эндотелина-1 (ЕТ-1) и фактора фон Виллебранда (vWF) проводили в соответствии с инструкциями набора (все наборы были приобретены в NanJing JianCheng Bioengineering Institute, Цзянсу, Китай).
Окрашивание гематоксилином-эозином (HE)
Образцы фиксировали 10% -ным формалином более 24 ч и хранили в парафиновых блоках. Парафиновые блоки депарафинизировали ксилолом в течение 20 минут, дегидратировали с помощью нисходящего градиента спирта (100%, 95%, 80%, 75%) в течение 1 минуты и окрашивали гематоксилином в течение 10 минут. Затем ткани промывали дистиллированной водой, дифференцировали соляной кислотой и этанолом в течение 30 с и замачивали в теплой воде при 50 ° C на 5 мин. Окрашенные раствором эозина, ткани промывали дистиллированной водой, обезвоживали 70% и 90% спиртом, очищали ксилолом и герметизировали нейтральной камедью. Морфологию тканей наблюдали под мощным микроскопом.
Наблюдение с помощью просвечивающего электронного микроскопа
Запасные ткани фиксировали 2,5% глутаровым диальдегидом и 1% осмитиновой кислоты, дегидратировали и затем заливали смолой Epon812. Полутонкие срезы окрашивали толуоловым синим, обрезали и делали ультратонкие срезы. Срезы окрашивали уранилацетатом и цитратом свинца и наблюдали с помощью просвечивающего электронного микроскопа JME-2000EX (Hitachi High-Technologies Co., Ltd., Шанхай, Китай).
Конечная дезоксинуклеотидилтрансфераза-опосредованная дезоксиуридинтрифосфат-биотиновая окраска концевых меток (TUNEL)
Окрашивание TUNEL предполагалось для наблюдения апоптоза эндотелиальных клеток у крыс IA на основе набора TUNEL (Roche, Базель, Швейцария). Подготовленные срезы аневризмы крысы дважды промывали ксилолом (5 мин / время) и обезвоживали нисходящей серией спирта (100%, 95%, 80%, 75%) 3 раза (5 мин / время). Ткани обрабатывали раствором протеазы К с концентрацией 20 мкг / мл без ДНКазы в течение 15–30 мин, добавляли по каплям 50 мкл реакционного раствора TUNEL в течение 60 мин и проявляли с помощью 50 мкл диаминобензидина (DAB) при 25 ° C в течение 10 мин. Затем срезы контрастировали гематоксилином, дегидратировали градиентным спиртом, очищали ксилолом и герметизировали нейтральной камедью. Срезы наблюдали под оптическим микроскопом и рассчитывали индекс апоптоза.
Выделение и идентификация сосудистых эндотелиальных клеток аневризмы
Выделение эндотелиальных клеток выполняли в соответствии с методом, проведенным Boscolo et al. [16]. Ткани IA были разрезаны на 3 мм 2 фрагменты. Ткани инкубировали с 0,1% коллагеназы B / 0,1% диспазы (Roche) в течение 25 мин при 37 ° C. Затем предварительно отделенные ткани растирали в порошок в течение 2 минут с помощью пипетки на 2 мл и фильтровали с помощью сетчатого фильтра 100 мкм (Thermo Fisher Scientific, Рокфорд, Иллинойс, США). Затем клеточную суспензию центрифугировали, а затем ресуспендировали в среде MV2 (включая факторы роста и 20% фетальную бычью сыворотку) (PromoCell, Гейдельберг, Германия). Клетки засевали в количестве 1 × 10 4 . клеток / мл в культуральной колбе, покрытой 1 мкг / см 2 фибронектин. Следуя методу, описанному Jackson et al. [17] клетки с 80–100% слияниями разделяли с помощью бусинок (Dynabeads M-450 Tosylactived, Oxoid, Hampshire, UK), покрытых Ulex europaeus Agglutinin I (UEA) (Vector Laboratories, Ltd., Питерборо, Великобритания). . Эндотелиальные клетки, прикрепленные к покрытым лектином шарикам, собирали с помощью концентратора магнитных частиц, а неконъюгированные клетки промывали базальной средой. Положительные по UEA клетки ресуспендировали в культуральной среде и высевали в покрытые фибронектином культуральные колбы для улучшения адгезии и скорости роста клеток.
Клетки выращивали в MV2 на предметных стеклах, покрытых фибронектином. Когда слияние клеток достигало 80–100%, клетки закрепляли в ацетоне при 4 ° C и обрабатывали 1% Triton X-100 в течение 5 минут, а затем 0,5% бычьим сывороточным альбумином (BSA) в течение 15 минут. Клетки капали с первичным антителом против vWF (1:300, Abcam, Кембридж, Массачусетс, США) и инкубировали в течение 2 часов (NC выполняли в отсутствие первичного антитела), капая с предполагаемым пероксидазой хрена иммуноглобулином G (1:150, Abcam) и инкубировали 30 мин. Затем клетки проявляли с помощью 50 мкл DAB при 25 ° C в течение 5 минут, контрастировали гематоксилином, дифференцировали 0,1% соляной кислотой, обезвоживали спиртом с последующим удалением ксилола и герметизацией нейтральной десны. После высыхания клетки фотографировали под инвертированным микроскопом.
Группирование и трансфекция клеток
Эндотелиальные клетки сосудов аневризмы в логарифмической фазе были разделены на 5 групп:пустая группа (эндотелиальные клетки сосудов аневризмы без какого-либо лечения), группа sh-NC (эндотелиальные клетки сосудов аневризмы, трансфицированные плазмидой sh-MALAT1 NC), группа sh-MALAT1 ( эндотелиальные клетки сосудов аневризмы, трансфицированные плазмидой sh-MALAT1), группа Oe-NC (эндотелиальные клетки сосудов аневризмы, трансфицированные плазмидой Oe-MALAT1 NC) и группа Oe-MALAT1 (эндотелиальные клетки сосудов аневризмы, трансфицированные плазмидой Oe-MALAT1). Вышеупомянутые плазмиды были синтезированы Genechem. Трансфекцию клеток проводили в соответствии с инструкциями липофектамина TM Реагент 2000 (11668-027, Invitrogen, Carlsbad, California, USA).
Анализ 3- (4, 5-диметилтиазол-2-ил) -2,5-дифенилтетразолийбромида (МТТ)
Эндотелиальные клетки сосудов каждой группы высевали на 96-луночный планшет с плотностью 3 × 10 4 . клеток / мл и культивировали при 37 ° C, 5% CO 2 за 48 ч. В каждой группе было 5 параллельных лунок, и к каждой лунке добавляли 20 мкл свежего раствора МТТ (5 мг / мл, Sigma, Сент-Луис, Миссури, США). После 4-часовой реакции клетки смешивали с 200 мкл диметилсульфоксида. После полного растворения значение оптической плотности клеток в каждой группе измеряли с помощью считывающего устройства для микропланшетов (BioRad, Hercules, Калифорния, США) при 490 нм.
Проточная цитометрия
Распределение клеточного цикла проверяли окрашиванием йодидом пропидия (PI). Эндотелиальные клетки сосудов отделяли, центрифугировали, ресуспендировали в предварительно охлажденном 75% этаноле и закрепляли в течение ночи при -20 ° C. Клетки центрифугировали, чтобы удалить супернатант. К клеткам добавляли 450 мкл PBS, добавляли 100 мкл РНКазы A и окрашивали 400 мкл PI при 4 ° C в течение 30 минут, избегая света. Проточный цитометр (FACSCalibur, Becton, Dickinson and Company, Нью-Джерси, США) был использован для тестирования распределения клеточного цикла.
Апоптоз клеток тестировали двойным окрашиванием аннексином V / PI. Отделенные эндотелиальные клетки собирали и промывали 3 раза PBS. Клетки ресуспендировали в 100 мкл предварительно охлажденного 1 × связывающего буфера и смешивали с 5 мкл аннексина и 5 мкл ИП соответственно. Апоптоз клеток проверяли с помощью проточного цитометра. С AnnexinV в качестве поперечной оси и PI в качестве продольной оси левый верхний квадрант обозначал механически поврежденные клетки (AnnexinV - / PI + ), правый верхний квадрант для клеток с поздним апоптозом или некротических клеток (AnnexinV + / PI + ), левый нижний квадрант для отрицательных нормальных клеток (AnnexinV - / PI - ) и правый нижний квадрант для клеток с ранним апоптозом (AnnexinV + / PI - ). Общее количество апоптотических клеток (AnnexinV + / PI - и AnnexinV + / PI + ) были рассчитаны и выражены в процентах.
Количественная полимеразная цепная реакция с обратной транскрипцией (RT-qPCR)
Суммарные РНК в тканях и клетках извлекали с помощью Trizol (Takara Biotechnology Ltd., Далянь, Китай) и определяли концентрацию и чистоту РНК. Процесс обратной транскрипции РНК в комплементарную ДНК проводили в соответствии с инструкциями набора для обратной транскрипции (K1621, Fermentas, Maryland, NY, USA). Последовательности праймеров MALAT1, miR-143 и VEGFA (таблица 1) были разработаны и составлены Genechem. Чтобы оценить экспрессию lncRNA, miRNA или мРНК, RT-qPCR проводили с использованием SYBR GreenPCR Master Mix (Takara, Tokyo, Japan) с системой Roche Real-Time PCR (Roche). U6 был установлен как внутренний параметр miR-143, тогда как MALAT1 и VEGFA с глицеральдегид-3-фосфатдегидрогеназой (GAPDH) как внутренние параметры. Относительные уровни транскрипции генов-мишеней вычисляли по 2 - △△ Ct метод.
Вестерн-блоттинг
Общий белок из тканей и клеток извлекали. Концентрацию белка определяли согласно инструкциям набора для бицинхониновой кислоты (Boster Biological Technology Co. Ltd., Ухань, Хубэй, Китай). Белок разделяли электрофорезом в 10% полиакриламидном геле (Boster Biological Technology). Мембрану переносили на мембрану из поливинилиденфторида, а затем герметизировали 5% БСА на 1 час. Мембрану инкубировали с первичным антителом против Ki-67 (1:1000), VEGFA (1:1000), vWF (1:1000) и матриксной металлопротеиназой (MMP) -9 (1:1000, Abcam, Cambridge, UK). , Cyclin D1 (1:1000), Bax (1:1000) и Bcl-2 (1:1000, Santa Cruz Biotechnology, Санта-Крус, Калифорния, США) и GAPDH (1:2000, Jackson Immuno Research, Grove, Пенсильвания, США) и вторичным антителом (1:500, Jackson Immuno Research), меченным перекисью хрена. Мембрана была получена с помощью двухцветной инфракрасной флюоресцентной сканирующей системы визуализации Odyssey, а значения серого в полосах были измерены с помощью программного обеспечения для анализа изображений Quantity One.
Анализ двойного люциферазного репортерного гена
Сайты связывания MALAT1 и miR-143 были спрогнозированы и изложены на сайте биоинформатики (https://cm.jefferson.edu/rna22/Precomputed/). Связь связывания между MALAT1 и miR-143 дополнительно подтверждали с помощью анализа репортерного гена люциферазы. Фрагмент гена синтетической 3'-нетранслируемой области (3'UTR) MALAT1 вводили в репортерные векторы люциферазы pmiR-Report (Thermo Fisher Scientific) для генерации MALAT1 дикого типа (MALAT1-WT) по участкам эндонуклеазы Bamh1 и Ecor1. Сайт мутации комплементарной последовательности последовательности был разработан на MALAT1-WT, и целевой фрагмент был вставлен в репортерные векторы люциферазы pmiR-Report для получения мутантного типа MALAT1 (MALAT1-MUT) с помощью ДНК-лигазы Т4 после расщепления рестрикционной эндонуклеазой. Правильно секвенированные MALAT1-WT и MALAT1-MUT были котрансфицированы миметиком NC и miR-143 в эндотелиальные клетки сосудов. Клетки собирали и лизировали через 48 часов после трансфекции, и активность люциферазы тестировали с помощью набора для обнаружения люциферазы (BioVision, Сан-Франциско, Калифорния, США) с люминометром (Glomax20 / 20, Promega, Мэдисон, Висконсин, США).>
Целевое соотношение miR-143 и VEGFA и сайт связывания miR-143 и VEGFA 3'UTR прогнозировалось через веб-сайт биоинформатики (http://www.targetscan.org/vert_72/). Последовательность промоторной области 3'UTR VEGFA, содержащей сайт связывания miR-143, была составлена и клонирована в репортерные векторы люциферазы pmiR-Report для создания VEGFA-WT. На основе этого репортера сайт связывания VEGFA-WT и miR-143 был мутирован с образованием VEGFA-MUT. Репортер VEGFA-WT или VEGFA-MUT смешивали с имитатором NC или miR-143, а затем котрансфицировали в эндотелиальные клетки сосудов в течение 48 часов. После этого клетки лизировали и проверяли активность люциферазы с помощью набора для определения люциферазы.
Анализ РНК-Pull Down
Чтобы проверить взаимосвязь связывания между miR-143 и MALAT1, был реализован анализ РНК-pull down. Три биотин-меченные последовательности миРНК Bio-miR-143-WT, Bio-miR-143-MUT и Bio-miR-NC были сконструированы и переданы компании GenePharma (Шанхай, Китай). Эти биотинилированные олигонуклеотиды трансфицировали в клетки в течение 48 часов. Затем клетки собирали и инкубировали со специфическим клеточным лизатом (Ambion, Остин, Техас, США) в течение 10 мин. После этого лизат инкубировали гранулами стрептавидина M-280, предварительно покрытыми не содержащей РНКаз и дрожжевой тРНК (все от Sigma) при 4 ° C в течение 3 часов, затем дважды очищали холодным раствором для лизиса, трижды с низким солевой буфер и один раз с буферным раствором с высоким содержанием соли. Антагонистический зонд miR-143 был создан как NC. Суммарную РНК извлекали с помощью Trizol, а затем тестировали уровень обогащения MALAT1 с помощью RT-qPCR.
Статистический анализ
Все данные были обработаны программой SPSS 21.0 (IBM Corp. Armonk, NY, США). Данные измерений были представлены в виде среднего значения ± стандартное отклонение. Сравнение двух групп проводилось по независимой выборке t тест, в то время как сравнения между несколькими группами оценивались с помощью одностороннего дисперсионного анализа (ANOVA), а попарное сравнение осуществлялось с помощью теста множественных сравнений Тьюки. Связь между экспрессией MALAT1 и клинико-патологическими особенностями IA определялась с помощью критерия хи-квадрат. P значение менее 0,05 свидетельствует о статистически значимой разнице.
Результаты
MALAT1 и VEGFA сверхэкспрессируются, а miR-143 подавляется в тканях IA
Экспрессия ET-1 и vWF в сыворотке в группе IA и контрольной группе была обнаружена с помощью ELISA, и результаты показали, что экспрессия ET-1 и vWF увеличилась в группе IA по сравнению с контрольной группой (обе P <0,05) (рис. 1а, б).
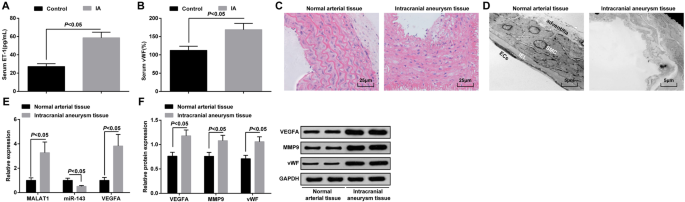
MALAT1 и VEGFA сверхэкспрессируются, а miR-143 подавляется в тканях IA. а Экспрессия ЕТ-1 в сыворотке крови пациентов с ИА и пациентов с височной эпилепсией с помощью ELISA. б Экспрессия vWF в сыворотке пациентов с ИА и пациентов с височной эпилепсией с помощью ELISA. c Патологическое наблюдение тканей IA и нормальных артериальных тканей по окрашиванию HE. г Морфологическое наблюдение тканей IA и нормальных артериальных тканей с помощью просвечивающего электронного микроскопа. е Экспрессия мРНК MALAT1, miR-143 и VEGFA в тканях IA и нормальных артериальных тканях с помощью RT-qPCR. е Экспрессия белков VEGFA, MMP-9 и vWF в тканях IA и нормальных артериальных тканях методом вестерн-блоттинга. Эндотелиальные клетки (ЭК), внутренняя эластическая пластинка (ВЭЛ), гладкомышечные клетки (ГМК). Данные измерений были представлены как среднее значение ± стандартное отклонение; сравнения между группами проводились по независимой выборке t тест
Патологические изменения тканей ИА наблюдали по окрашиванию НЕ. В нормальных артериальных тканях не наблюдалось явных повреждений энтодермы, эндотелиальных клеток и гладкомышечных клеток, клетки были аккуратно расположены и имели законченную структуру. В тканях IA присутствовали поврежденные эндотелиальные клетки, дегенерированные гладкомышечные клетки, ослабленная артериальная стенка, разорванные эластические волокна и инфильтрованные воспалительные клетки (рис. 1c).
Изменения ультраструктуры ИА и нормальных артериальных тканей наблюдали с помощью просвечивающего электронного микроскопа. Было обнаружено, что эндотелиальные клетки были полными, а структура адвентиции не была повреждена; в нормальных артериальных тканях не было обнаружено разрыва внутренней эластической мембраны или апоптозных гладкомышечных клеток. В тканях IA он был представлен тяжелой дегенерацией стенки кровеносного сосуда, в основном проявляющейся в исчезновении большинства эндотелиальных клеток, сильно нарушенном внутреннем эластическом слое, сильно поврежденных и деградированных гладкомышечных клетках, а также нарушении внешней мембраны кровеносного сосуда. (Рис. 1d).
RT-qPCR проводили для определения экспрессии мРНК MALAT1, miR-143 и VEGFA, в то время как вестерн-блоттинг анализ экспрессии белков VEGFA, MMP-9 и vWF в тканях IA и нормальных тканях артерий. Было продемонстрировано, что в отличие от нормальных артериальных тканей уровни экспрессии MALAT1, VEGFA, MMP-9 и vWF были повышены, а экспрессия miR-143 снижена в тканях IA (все P <0,05) (рис. 1д, е).
Оценка Ханта-Хесса, степень повреждения эндотелия и история курения коррелируют с экспрессией MALAT1 в тканях IA
В свете средней экспрессии MALAT1 пациенты были разделены на две группы:группу с низкой экспрессией и группу с высокой экспрессией. Была проанализирована взаимосвязь между экспрессией MALAT1 и клинико-патологическими особенностями пациентов с ИА. Результаты показали, что оценка Ханта-Хесса, степень повреждения эндотелия и история курения были связаны с экспрессией MALAT1 (все P <0,05), в то время как возраст, пол и хирургический режим не были связаны с экспрессией MALAT1 (все P > 0,05) (таблица 2).
Подавленный MALAT1 подавляет кровяное давление, экспрессию ET-1, vWF и MMP-9, а также индекс апоптоза эндотелиальных клеток сосудов крыс с IA
Как показано в Таблице 3, нокдаун MALAT1 ухудшился, в то время как восстановление MALAT1 повысило артериальное давление на 4-й и 12-й неделе.
ELISA показал, что подавление активности MALAT1 снижалось, тогда как повышающая регуляция MALAT1 повышала экспрессию ET-1 и vWF в сыворотке крови крыс с IA (рис. 2a, b).
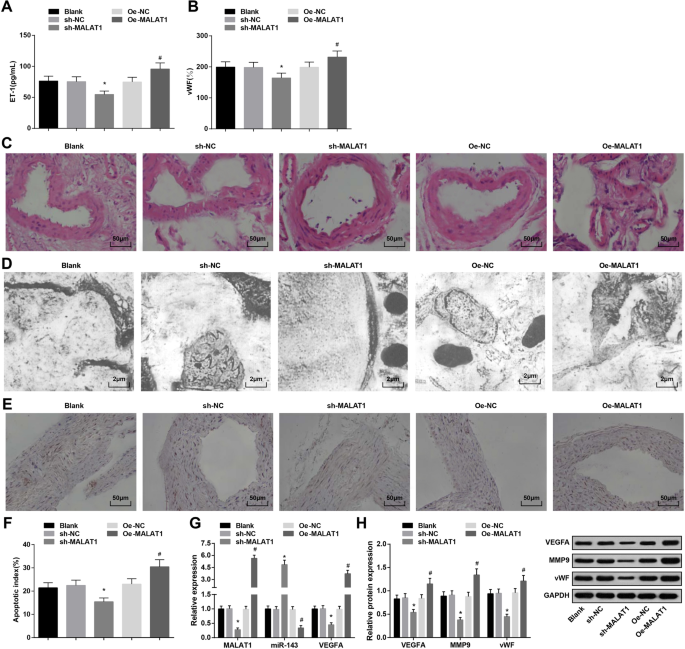
Подавленный MALAT1 подавляет кровяное давление, экспрессию ET-1, vWF и MMP-9, а также индекс апоптоза эндотелиальных клеток сосудов крыс с IA. а Экспрессия ЕТ-1 в сыворотке крови крыс с помощью ELISA. б Экспрессия vWF в сыворотке крови крыс с помощью ELISA. c Патологические изменения тканей ИА у крыс, наблюдаемые при окрашивании НЕ. г Ультраструктура тканей IA у крыс, наблюдаемая в просвечивающем электронном микроскопе. е Апоптоз эндотелиальных клеток сосудов при окрашивании TUNEL. е Индекс апоптоза эндотелиальных клеток сосудов крыс. г Экспрессия мРНК MALAT1, miR-143 и VEGFA в тканях IA крыс с помощью RT-qPCR. ч Экспрессия белков VEGFA, MMP-9 и vWF в тканях IA крыс методом вестерн-блоттинга. * P <0,05 по сравнению с группой sh-NC, # P <0,05 по сравнению с группой Oe-NC. Данные измерений были представлены как среднее значение ± стандартное отклонение, а сравнения между несколькими группами оценивались с помощью одностороннего дисперсионного анализа с последующим тестом множественных сравнений Тьюки
Патологические изменения тканей ИА в каждой группе исследовали с помощью окрашивания НЕ. Пустая группа, группа sh-NC и группа Oe-NC проявлялись повреждением внутренней мембраны, расслоением эндотелиальных клеток, дегенерированными клетками гладкой мускулатуры, уменьшенными клетками и слоями и истонченной стенкой артерии. В группе sh-MALAT1 энтодерма, эндотелиальные клетки, слой гладкомышечных клеток и слой наружной мембраны внутричерепной артерии были слегка повреждены, но расположены аккуратно. Группа Oe-MALAT1 показала исчезновение слоя интимы, расслоение эндотелиальных клеток, сломанные эластические волокна и инфильтрированные воспалительные клетки (рис. 2c).
Ткани IA крыс в каждой группе наблюдали с помощью просвечивающего электронного микроскопа. Было показано, что денатурированные эндотелиальные клетки, разрушенный субэндотелиальный слой, исчезновение внутреннего эластичного слоя и уменьшение гладкомышечных клеток были представлены в пустой группе, группе sh-NC и группе Oe-NC. Группа sh-MALAT1 продемонстрировала плоские эндотелиальные клетки, овальное ядро и увеличенные волокна коллагена, но без эластичного слоя. Группа Oe-MALAT1 была представлена исчезнувшими эндотелиальными клетками и отделенным эластичным слоем от мышечного слоя, который распался в просвете (рис. 2d).
Индекс апоптоза сосудистых эндотелиальных клеток у крыс IA тестировали с помощью окрашивания TUNEL. Выключение MALAT1 снижает апоптотический индекс сосудистых эндотелиальных клеток, в то время как сверхэкспрессия MALAT1 оказывает противоположный эффект (рис. 2e, f).
Обнаружение RT-qPCR экспрессии мРНК MALAT1, miR-143 и VEGFA и вестерн-блот-анализ экспрессии белков VEGFA, MMP-9 и vWF в тканях IA показал, что удаление MALAT1 подавляло экспрессию MALAT1, VEGFA, MMP-9 и vWF. и повышенная экспрессия miR-143. Напротив, повышение уровня MALAT1 оказывает противоположное влияние на экспрессию этих генов (рис. 2g, h).
Низкая экспрессия MALAT1 повышает жизнеспособность и сдерживает апоптоз эндотелиальных клеток сосудов в IA
Иммуногистохимическое окрашивание использовали для обнаружения экспрессии специфичного для эндотелия маркера vWF. Выявлено, что цитоплазма сосудистых эндотелиальных клеток IA была покрыта мелкими коричневыми частицами, что было положительным, в то время как цитоплазма в ее группе NC не имела коричневых частиц. Результаты подтвердили, что культивируемые клетки были эндотелиальными клетками (рис. 3a, b).
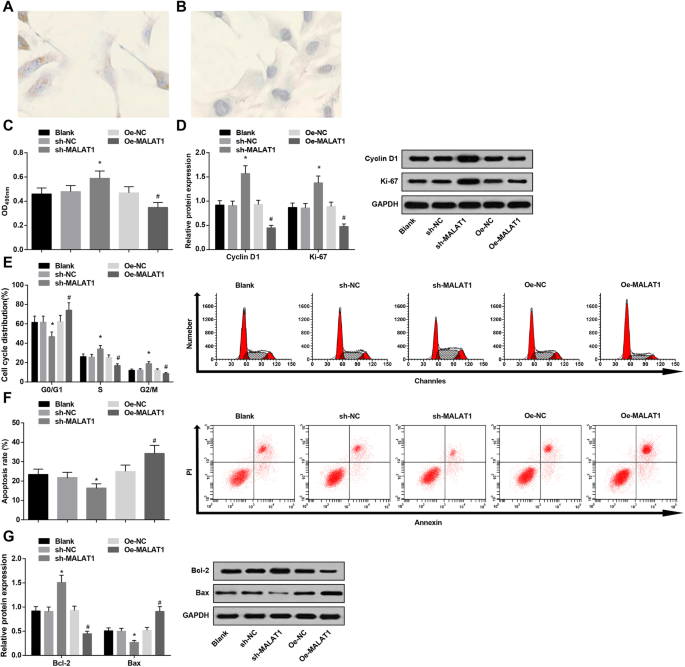
Низкая экспрессия MALAT1 увеличивает жизнеспособность и сдерживает апоптоз эндотелиальных клеток сосудов в IA. а Иммуногистохимическое окрашивание vWF в эндотелиальных клетках сосудов IA:эндотелиальные клетки сосудов IA были покрыты мелкими желтыми частицами. б Иммуногистохимическое окрашивание vWF в эндотелиальных клетках сосудов IA:эндотелиальные клетки сосудов IA не показали коричневых частиц в группе NC. c Жизнеспособность эндотелиальных клеток сосудов в каждой группе по методу МТТ. г Экспрессия белков CyclinD1 и Ki-67 в каждой группе по данным вестерн-блоттинга. е Изменения клеточного цикла в каждой группе при окрашивании ИП. е Скорость апоптоза клеток в каждой группе по двойному окрашиванию аннексином V / PI. г Экспрессия белков Bax и Bcl-2 в каждой группе по данным вестерн-блоттинга. * P <0,05 по сравнению с группой sh-NC, # P <0,05 по сравнению с группой Oe-NC. Данные измерений были представлены как среднее значение ± стандартное отклонение, а сравнения между несколькими группами оценивались с помощью одностороннего дисперсионного анализа с последующим тестом множественных сравнений Тьюки
МТТ-анализ, проточная цитометрия вместе с вестерн-блоттингом использовались для проверки жизнеспособности эндотелиальных клеток сосудов и апоптоза. Было показано, что уменьшение MALAT1 способствовало жизнеспособности эндотелиальных клеток сосудов (повышенная экспрессия циклина D1 и Ki-67) и подавляло апоптоз (уменьшалось количество клеток в фазе G0 / G1 и увеличивалось количество клеток в фазе S и G2 / M, уменьшалось Bax и повышалась экспрессия Bcl-2). . Однако активация MALAT1 действует противоположным образом по сравнению с уменьшением MALAT1 на жизнеспособность клеток и апоптоз (рис. 3c – g).
MiR-143 связан с MALAT1, а VEGFA является целевым геном miR-143
Экспрессию MALA1, miR-143 и VEGFA в эндотелиальных клетках сосудов каждой группы проверяли с помощью RT-qPCR и вестерн-блоттинга. Результаты показали, что нокдаун MALA1 подавлял экспрессию MALA1 и VEGFA и усиливал экспрессию miR-143. Напротив, активация MALA1 привела к увеличению экспрессии MALAT1 и VEGFA и снижению экспрессии miR-143 (рис. 4a, b).
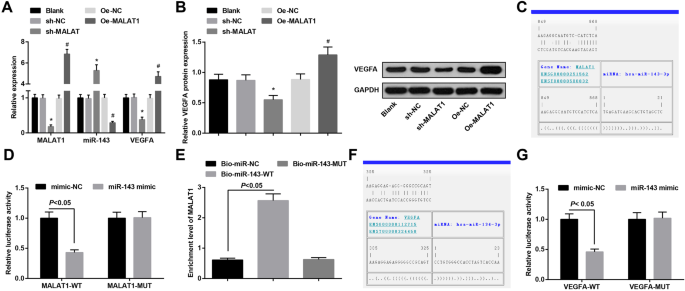
MiR-143 связан с MALAT1, а VEGFA является геном-мишенью для miR-143. а Экспрессия мРНК MALAT1, miR-143 и VEGFA в сосудистых эндотелиальных клетках аневризмы в каждой группе. б Экспрессия белка VEGFA в эндотелиальных клетках аневризмы сосудов в каждой группе. c Сайты связывания MALAT1 и miR-143 предсказаны сайтом биоинформатики. г Регуляторная связь MALLA1 и miR-143 подтверждена двойным анализом репортерного гена люциферазы. е Связь связывания между MALAT1 и miR-143 подтверждается анализом РНК-pull down. е Сайты связывания miR-143 и VEGFA предсказаны сайтом биоинформатики. г Регуляторная связь miR-143 и VEGFA подтверждена двойным анализом репортерного гена люциферазы. * P <0,05 по сравнению с группой sh-NC, # P <0,05 по сравнению с группой Oe-NC. Данные измерений были представлены как среднее значение ± стандартное отклонение, сравнения между двумя группами оценивались с помощью независимой выборки t тест, и сравнения между несколькими группами оценивались с помощью одностороннего дисперсионного анализа с последующим тестом множественных сравнений Тьюки
Область специфического связывания между MALAT1 и miR-143 была определена с помощью программного обеспечения для онлайн-анализа (рис. 4c). Результаты двойного анализа репортерного гена люциферазы показали, что активность люциферазы была нарушена в группе миметика MALAT1-WT + miR-143 по сравнению с группой миметика MALAT1-WT + мимик-NC ( P <0,05). Однако не было явной разницы в активности люциферазы в группе миметиков MALAT1-MUT + miR-143 по сравнению с группой миметиков MALAT1-MUT + mimic-NC ( P > 0,05), что указывает на то, что miR-143 специфически связывается с MALAT1 (рис. 4d). Результаты анализа РНК-pull down показали, что уровень обогащения MALAT1 в группе Bio-miR-143-WT был повышен по сравнению с группой Bio-miR-NC ( P <0,05), но не было явной разницы в уровне обогащения MALAT1 в группе Bio-miR-143-MUT ( P > 0,05) (рис. 4д).
Программа биоинформатики обнаружила целевую связь между miR-143 и VEGFA (рис. 4f). Результаты активности люциферазы показали, что относительная активность люциферазы подавляется после того, как миметик VEGFA-WT и miR-143 котрансфицирован в эндотелиальные клетки сосудов ( P <0,05). Однако на относительную активность люциферазы сосудистых эндотелиальных клеток не влияла котрансфекция VEGFA-MUT и miR-143-миметика ( P > 0,05) (рис. 4ж). Было указано, что VEGFA является прямым геном-мишенью miR-143.
Обсуждение
ИА - серьезное внутричерепное заболевание, которое в основном приводит к субарахноидальному кровоизлиянию [18]. Предыдущее исследование продемонстрировало участие связанных с lncRNA конкурентных эндогенных сетей РНК в IA [19]. Также недавнее исследование предоставило доказательство того, что функциональный полиморфизм в промоторной области гена miR-143/145 связан с риском IA [20]. В исследовании, проведенном Сюй и др. . , показано, что сверхэкспрессия ангиогенных факторов, таких как VEGFA, может быть связана с образованием и разрывом IA [21]. Чтобы объяснить молекулярный механизм MALAT1 в IA, была проведена серия анализов, и результаты показали, что IA, вызванное повреждением эндотелия сосудов, регулируется сигнальной осью MALAT1 / miR-143 / VEGFA.
Во-первых, наше исследование предоставило существенные доказательства того, что MALAT1 и VEGFA активируются, а miR-143 подавляется в тканях IA. Недавнее исследование показало, что MALAT1 является одной из днРНК с наибольшей активностью в процессе церебральной ишемии [22]. Другое исследование показало, что MALAT1 активируется в раковых клетках яичников и намеревается участвовать в процессах апоптоза, миграции и пролиферации раковых клеток яичников [23]. Исследование профиля экспрессии неразорвавшейся и разорванной ИА продемонстрировало, что экспрессия ангиогенных факторов, таких как VEGFA, повышается при разрыве аневризмы [21]. Более того, клиническое исследование показало, что кластер miR-143/145 у пациентов с IA подавлен по сравнению со здоровыми субъектами [11]. Кроме того, ранее обсуждалось, что miR-143/145 принимает участие в различных биологических процессах, связанных с формированием аневризмы, и подавляется у пациентов с IA [20]. Все эти выводы более или менее перекликаются с результатами предыдущих исследований.
За исключением вышеупомянутых результатов, в этом исследовании также изучалась функциональная роль MALAT1 в IA с помощью анализов потери функции и потери функции. Можно резюмировать, что подавление MALAT1 снижает артериальное давление, уровни ЕТ-1 в сыворотке крови и экспрессию vWF и MMP-9 в тканях IA. Ранее предполагалось, что подавление MALAT1 сдерживает повышающую регуляцию индуцированного глюкозой продукта транскрипции ЕТ-1 [24]. Также сообщается, что эктопическая экспрессия MALAT1 является индуктором генерации vWF [25]. Другое исследование подтвердило, что истощение MALAT1 в макрофагах костного мозга подавляет экспрессию MMP-9 [26].
Также были проведены клеточные эксперименты для дальнейшего подтверждения функций MALAT1 в IA. Результаты показали, что нокдаун MALAT1 способствует жизнеспособности эндотелиальных клеток сосудов и подавляет апоптоз в IA. Сходным образом было задокументировано, что нарушение MALAT1 улучшает архитектуру стенок аорты и замедляет рост аневризмы [8]. В дополнение к результатам нашего исследования, есть исследования, подчеркивающие, что плохая экспрессия MALAT1 вызывает апоптоз и сдерживает пролиферацию клеток острого миелоидного лейкоза [27]. Другое исследование также продемонстрировало ингибирующее действие нокдауна MALAT1 на пролиферацию хрящевых клеток остеоартрита человека [28]. Кроме того, предыдущие исследования в целом подтверждают, что подавление MALAT1 может вызывать апоптоз и ослаблять пролиферацию клеток глиомы [29]. Более того, низкая экспрессия MALAT1, индуцированная интерференцией РНК, способствует апоптозу и подавляет пролиферацию клеток множественной миеломы [30]. В совокупности эти исследования до некоторой степени объяснили молекулярный механизм MALAT1 в IA.
Кроме того, это исследование доказало, что miR-143 связывается с MALAT1, а VEGFA является геном-мишенью для miR-143. Сходным образом в статье утверждается, что MALAT1 связывается непосредственно с miR-143 и подавляет ее экспрессию [10]. Zhu et al. обнаружили, что MALAT1 выполняет свои функции посредством взаимодействия с miR-143 в клетках карциномы шейки матки [31]. Кроме того, подтверждено, что MALAT1 косвенно модулирует VEGFA через miR-200b-3p [32]. Более того, другое исследование показало, что miR-143-3p опосредует эффект ZFPM2 на ряд белковых мишеней в крови, включая VEGFA [33]. Тем не менее, взаимодействия между MALAT1, miR-143 и VEGFA в IA не обсуждались и требуют дальнейшего изучения.
Заключение
Из этих результатов ясно, что нокдаун MALAT1 подавляет апоптоз и способствует жизнеспособности сосудистых эндотелиальных клеток в IA путем модуляции оси miR-143 / VEGFA. Сеть коэкспрессии предполагает связь между MALAT1 и miR-143 с участием VEGFA. Результаты этого исследования частично раскрывают патогенез инициации и прогрессирования IA, а изученные цели могут быть особенно потенциальной отправной точкой для выявления патологии IA с другой точки зрения. В ограниченном объеме необходимы дальнейшие крупномасштабные исследования, чтобы всесторонне проиллюстрировать механизмы оси MALAT1 / miR-143 / VEGFA в IA.
Наноматериалы