


Мощный противоопухолевый иммуностимулирующий биосовместимый наногидрогель, изготовленный из ДНК
Аннотация
Неметилированные олигодезоксинуклеотиды CpG являются мощными иммуностимулирующими мотивами для активации как врожденной, так и приобретенной иммунной системы путем индукции антигенспецифических Т-клеточных ответов Th1-типа, но их нестабильность в сыворотке сильно влияет на эффективность их иммуностимуляторов. Здесь мы сконструировали новые наногидрогели иммуно-ДНК, состоящие из последовательностей тандемных повторов CpG-единиц, названных наногидрогелями CpG-MCA, посредством мультипримированной цепной амплификации. Было доказано, что наногидрогели CpG-MCA противостоят деградации и увеличивают пролиферацию и миграцию мышиных макрофагоподобных клеток RAW264.7. Кроме того, наногидрогели CpG-MCA эффективно индуцировали высокую экспрессию фактора некроза опухоли-α и интерлейкина-6 и заметно подавляли пролиферацию клеток U251, предполагая, что наногидрогели CpG-MCA, как ожидается, будут использоваться в качестве мощного противоракового иммуностимулятора. / P>
Бактериальная ДНК, содержащая неметилированные мотивы CpG, является чрезвычайно многообещающим адъювантом вакцины, антиаллергенами, а также иммунопротекторными и противораковыми агентами [1]; он может распознаваться рецепторами на эндосомальных внутренних клетках иммунной системы, такими как дендритные клетки (DC), макрофаги, Т-клетки, естественные киллеры (NK) и NKT-клетки [2]. Эти клетки врожденного иммунитета способны реагировать на неметилированные мотивы CpG посредством распознавания патоген-ассоциированных молекулярных паттернов (PAMP) для патоген-специфических молекул в патогенном микроорганизме. Было подтверждено, что олигонуклеотид CpG (CpG-ODN) может распознаваться Toll-подобным рецептором 9 (TLR9) [3] и индуцировать иммунный ответ Th1-типа через Myd88-зависимый сигнальный путь [4]. Однако эти недостатки CpG-ODN препятствовали его клиническому применению из-за его преждевременного созревания за счет адсорбции белка, переваривания эндонуклеазами в сыворотке [5] и нестабильности in vivo. Ожидается, что эти проблемы будут решены путем инкапсуляции однонитевой (ss) нуклеиновой кислоты в носитель для доставки или ее самосборки в наноструктуру, которая улучшит ее стабильность in vivo, а также повысит коэффициент интернализации для клеток врожденного иммунитета. В настоящее время для доставки CpG-ODN используются различные носители, такие как катионные полимерные полиэтиленимины (PEI) [6], липосомы [7, 8] и микрочастицы [9]; есть еще некоторые недостатки, которые необходимо исправить, например цитотоксичность, ограниченная скорость загрузки и т. д.
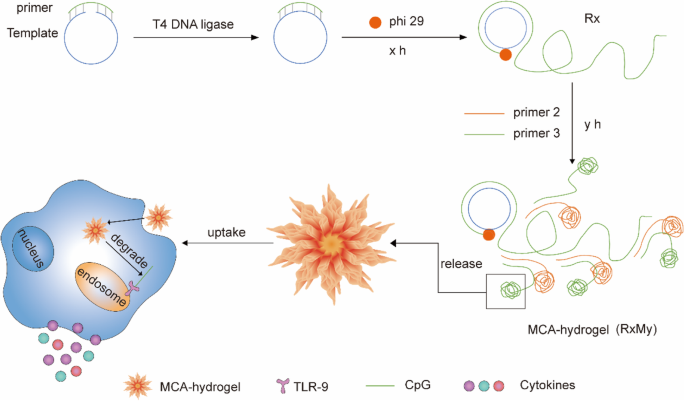
Материалы ДНК, состоящие из нуклеиновой кислоты, демонстрируют большой потенциал для использования в качестве носителей для доставки CpG-ODN. По сравнению с оцДНК материалы ДНК с двумерной или трехмерной структурой проявляют различные свойства, такие как легкое проникновение через клеточные мембраны и стимуляция макрофагов к секреции цитокинов [10]. X-образная ДНК была использована для доставки мотивов CpG и успешного увеличения иммуностимулирующей активности CpG-ODN за счет увеличения клеточного поглощения, тогда как повышенное клеточное поглощение частично связано с X-образной структурой [11]. Точно так же трехмерный тетраэдр ДНК, который может самоорганизовываться в наноструктуры с одинаковыми размерами, неинвазивно и эффективно вводился в клетки RAW264.7 для функционирования. Более того, согласно исследованиям [12], такой тетраэдр оказался механически устойчивым и нецитотоксичным. Недавно было продемонстрировано, что гидрогель CpG-RCA (гель CpG-RCA) с наноцветковой структурой, полученный посредством амплификации по катящемуся кругу (RCA), способен доставлять иммуностимулирующий сигнал, противостоять деградации нуклеаз, увеличивать секрецию иммунных цитокинов и подавляют пролиферацию клеток Т-лимфоцитов острого лимфоцитарного лейкоза человека (CCRF-CEM) [13]. Эти результаты свидетельствуют о том, что форма и структура материалов ДНК играют важную роль в усилении клеточного поглощения и повышении эффективности иммунной стимуляции. Поскольку клетки CCRF-CEM происходят от лейкоза Т-лимфобластов человека, типа гематологической злокачественной опухоли, она отличается от гематологической злокачественной опухоли; солидные опухоли часто окружены иммуносупрессивным микроокружением, которое может препятствовать действующему противоопухолевому иммунитету [14]. Для этого мы сконструировали иммуностимулятор ДНК, содержащий гораздо больше копий CpG-ODN посредством мультипримированной цепной амплификации (MCA) [15], а не RCA [16]. Пользуясь преимуществом принципа комплементарного спаривания оснований, CpG-вовлеченный праймер и специально сконструированная матричная последовательность были смешаны с лигазой и удлинены полимеразой phi29 в присутствии свободных dNTP. RCA (R) или MCA (M) отреагировали на x или y часы должны быть отдельно представлены как Rx или My (рис. 1); продукты идентифицировали электрофорезом в агарозном геле (рис. 2а). На основе реакции MCA полученные продукты мы назвали гидрогелями CpG-MCA (гелями CpG-MCA), обладающими сотнями или тысячами тандемных CpG из-за большого увеличения копий мотива CpG. Гели CpG-MCA также были мощным иммуностимулятором, который значительно увеличивал секрецию цитокинов из клеток RAW264.7 и эффективно ингибировал пролиферацию клеточных линий глиомы человека U251. Мы ожидали, что это исследование будет способствовать созданию нового иммуностимулятора наногидрогеля на основе материалов ДНК, и способствовали его применению для иммунотерапии опухолей [17].
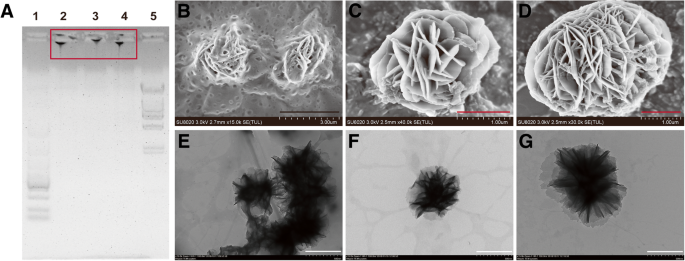
Изображения гелей CpG-MCA и гелей CpG-RCA. а Изображение электрофореза в агарозном геле гелей CpG-RCA (R12) и гелей CpG-MCA (R4M4 и R4M8). Дорожка 1:стандартный маркер ДНК MW, расщепленный λ-Hind III; дорожка 2, R12; дорожка 3, R4M4; дорожка 4, R4M8, дорожка 5, маркер ДНК DL5000. СЭМ-изображения R12 ( b ), R4M4 ( c ) и R4M8 ( d ). ПЭМ-изображения R12 ( e ), R4M4 ( f ) и R4M8 ( g ). Черная шкала 3 мкм; красная шкала 1 мкм; белая шкала - 500 нм
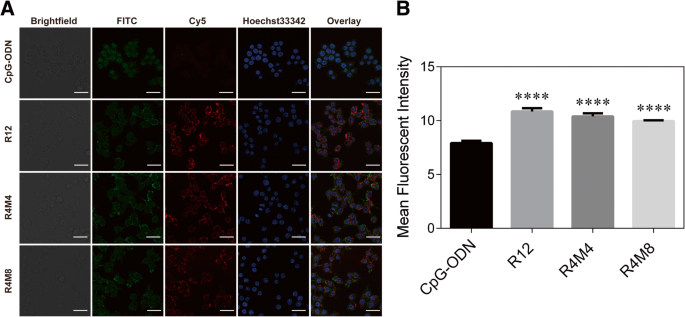
Изображения конфокального микроскопа и средняя интенсивность флуоресценции в клетках RAW 264.7, обработанных CpG-ODN, гелем CpG-RCA (R12) и гелями CpG-MCA (R4M4 и R4M8) (100 нМ эквивалентов CpG). Гели CpG-MCA и гель CpG-RCA были помечены Cy5 (красный) путем добавления Cy5-dCTP для их клеточного поглощения. CpG-ODN, меченный Cy5, использовали в качестве контрольной группы. а Изображения с конфокальной микроскопии гелей CpG-RCA и гелей CpG-MCA на клетках RAW 264.7 после инкубирования в течение 2 часов. б Средняя интенсивность флуоресценции Cy5 в клетках RAW264.7. Результаты выражены как среднее значение ± стандартное отклонение трех независимых экспериментов. **** P <0,0001
Все олигонуклеотиды были приобретены у Sangon Biotech (Shanghai) Co., Ltd. и очищены с помощью высокоэффективной жидкостной хроматографии. Наборы dNTP (по 100 мМ каждый), ДНК-полимераза phi29 (10 Ед / мкл) и буфер для реакции ДНК-полимеразы 10 × phi29 (330 мМ трис-ацетат (pH 7,9 при 37 ° C), 100 мМ ацетат магния, 660 мМ ацетат калия , 1% ( v / v ) Tween 20, 10 мМ DTT) были приобретены в Thermo Fisher Scientific (Уолтем, Массачусетс, США). ДНК-лигаза Т4 (400 Ед / мкл), 5'-трифосаденин (АТФ) и 10 × буфер для реакции ДНК-лигазы Т4 (50 мМ Трис-HCl, 10 мМ MgCl 2 , 1 мМ АТФ, 10 мМ DTT, pH 7,5 при 25 ° C) были приобретены в New England Biolabs, Inc. (Ипсвич, Массачусетс). Цианин 5-dCTP был приобретен у PerkinElmer, Inc. (Уолтем, Массачусетс, США). Центробежные фильтрующие устройства Amicon Ultra-0.5 были приобретены у Merck KGaA (Дармштадт, Германия). Вода, использованная в этой статье, была очищена с помощью системы очистки сверхчистой воды Millipore Synergy UV. Фетальная бычья сыворотка Gibco (FBS), среда Игла, модифицированная Дульбекко (DMEM, с высоким содержанием глюкозы, L-глутамина, фенолового красного, пирувата натрия, без HEPES), трипсин-ЭДТА (0,25%) и пенициллин (10000 Ед / мл) - стрептомицин (10000 мкг / мл) был приобретен в Thermo Fisher Scientific (Уолтем, Массачусетс, США). Наборы для ELISA были приобретены в R&D Systems, Inc. Набор для подсчета клеток-8 (CCK-8) был приобретен в Dojindo (Кумамото, Япония). Линия макрофагоподобных клеток мыши (клетки RAW264.7) была получена из банка клеток Китайской академии наук (Шанхай, Китай). Клеточные линии глиомы человека (клетки U251) были получены из банка клеток Китайской академии наук (Шанхай, Китай).
Длинную одноцепочечную ДНК с фосфорилированной группой на 5'-конце с равным соотношением праймеров CpG-ODN (праймер 1) смешивали в реакционном буфере 1 × phi29 и отжигали при 95 ° C в течение 5 мин и медленно охлаждали до 4 ° C при температуре скорость - 1 ° C / с с использованием термоциклера (Bio-Rad T100, Германия). После отжига добавляли 20 ед. / Мкл ДНК-лигазы Т4 с АТФ и реакционным буфером ДНК-лигазы Т4 и инкубировали при 4 ° C в течение ночи. Ферменты неактивны при 75 ° C в течение 10 мин.
Круговую матрицу ДНК (10 мкл) смешивали с 1 × реакционным буфером для ДНК-полимеразы phi29, 4 мМ dNTP для каждого и добавляли 5 ед. ДНК-полимеразы phi29 и стерильный DDW, всего 50 мкл. Смесь инкубировали при 30 ° C при встряхивании в течение 12 ч (R12). Для образования геля MCA после 4 ч реакции RCA к полученной смеси добавляли 500 пМ праймера 2 и праймера 3, соответственно, для инкубации в течение оставшихся часов при 30 ° C без добавления дополнительных реагентов (R4M4 и R4M8). Полимеразу phi29 инактивировали при 65 ° C в течение 10 мин. Гели CpG-RCA и CpG-MCA очищали ультрафильтрацией.
Концентрацию гелей CpG-MCA и геля CpG-RCA измеряли на основании количества копий CpG, задействованных во всех гидрогелях в группе лечения. Поскольку dNTP, добавленный в реакционную систему, составлял 4 мМ для каждого, одна кольцевая матрица содержала 81 нуклеотид и 1 копию CpG. Если Abs - поглощение dNTP при 260 нм и ε - коэффициент экстинкции dNTP при 260 нм, тогда можно измерить израсходованный dNTP в реакции и рассчитать общее количество копий CpG по следующему уравнению:CpG-копии =(4 мМ × 4 - Abs / ε × 1000000) / 81 [13]. Абс и ε dNTP измеряли с помощью NanoDrop 2000c.
Электрофорез в агарозном геле использовали для оценки образования и разложения гелей CpG-MCA и образования круглой матрицы. Гидрогели обрабатывали на 1% агарозном геле при 100 В в течение 60 минут, а круглый шаблон запускали на 3% агарозном геле при 100 В в течение 60 минут.
Просвечивающая электронная микроскопия (ТЕМ, Hitachi HT7700, Япония) была использована для характеристики внутренней структуры и приблизительного размера гелей CpG-MCA. Гели CpG-MCA исследовали ультразвуком в течение 30 мин перед нанесением на медь и сушкой. Тесты проводились в центральной лаборатории госпиталя Ренджи. Сканирующую электронную микроскопию (SEM, Hitachi SU8020, Япония) использовали для определения морфологии гелей CpG-MCA. Гели CpG-MCA были исследованы ультразвуком в течение 30 минут перед нанесением на чистую кремниевую пластину, и образец был покрыт металлом Au.
Захват клеток регистрировали с помощью конфокального микроскопа Leica. Клетки RAW264.7 высевали на конфокальную чашку Петри при плотности 2 × 10
5
. клеток / мл. После двукратной промывки фосфатным буфером (PBS) клетки инкубировали со 100 нМ Cy5-меченным CpG-ODN, гелем CpG-RCA и гелями CpG-MCA в свежей среде DMEM в течение 2 ч при 37 °. Затем клетки трижды промывали PBS и фиксировали 4% параформальдегидом в течение 30 минут, затем клетки окрашивали FITC-фаллоидином и Hoechst 33342. Все изображения получали с использованием лазерного конфокального микроскопа Leica. Полуколичественный показатель средней интенсивности флуоресценции был рассчитан Image J, Java-приложением для анализа изображений.
Клетки RAW264.7 засевали с плотностью 7 × 10
4
. клеток / мл в 24-луночном планшете, культивировали в течение 24 ч перед использованием. Клетки инкубировали в присутствии гелей CpG-MCA и других групп при 37 ° C в течение 8 часов для TNF-α и 24 часов для IL-6; собирали супернатанты. Уровни цитокинов в супернатантах определяли с помощью иммуноферментного анализа (ELISA) в соответствии с протоколами, предложенными производителем.
Уровни экспрессии генов оценивали с помощью количественной ПЦР в реальном времени (Q-PCR). Клетки RAW264.7 засевали с плотностью 1 × 10
6
. клеток / мл в 6-луночном планшете, культивировали в течение 24 ч перед использованием. Клетки инкубировали в присутствии гелей CpG-MCA и других групп при 37 ° C в течение 2 часов для TNF-α и TLR9 и 8 часов для IL-6 и других. Выделение и очистку мРНК проводили с использованием TRIzol Reagent (Thermo Fisher Scientific). Выделенную мРНК количественно оценивали с помощью NanoDrop 2000c. Один микрограмм общей РНК подвергали обратной транскрипции с использованием набора реагентов PrimeScript RT с gDNA Eraser (Takara Bio Inc.); амплификацию проводили в общем реакционном объеме 20 мкл с использованием TB Green Premix Ex Taq II (Takara Bio Inc) в соответствии с инструкциями производителя. Праймеры для генов следующие:GAPDH:F:AGGTCGGTGTGAACGGATTTG; R:TGTAGACCATGTAGTTGAGGTCA; TLR9:F:ATGGTTCTCCGTCGAAGGACT; R:GAGGCTTCAGCTCACAGGG; TNF-α:F:GACGTGGAACTGGCAGAAGAG; R:TTGGTGGTTTGTGAGTGTGAG; ИЛ-6:F:CCAAGAGGTGAGTGCTTCCC; R:CTGTTGTTCAGACTCTCTCCCT; CD86:F:GAGCTGGTAGTATTTTGGCAGG; R:GGCCCAGGTACTTGGCATT; CD206 (MRC1):F:CTCTGTTCAGCTATTGGACGC; R:CGGAATTTCTGGGATTCAGCTTC.
Клетки RAW264.7 засевали 70 мкл при плотности 5 × 10
5
. клеток / мл во вкладышах для культур в течение 24 ч перед использованием. Затем вставки осторожно удаляли и клетки дважды промывали перед обработкой каждой группы в свежей среде. Фотографии были собраны через 0 часов, 6 часов и 24 часа; площадь раны измеряли с помощью ImageJ. В каждой группе было по три повтора, и эксперимент был повторен трижды.
Цитотоксичность оценивали с использованием CCK8. Клетки RAW264.7 высевали в 96-луночные планшеты и обрабатывали каждой группой в течение 24 часов. Затем в каждую лунку добавляли 10 мкл раствора CCK8 с последующей 1-2 ч инкубации при 37 ° C. Поглощение измеряли при 450 нм, в каждой группе было три повтора, и эксперимент повторяли трижды.
Клетки RAW264.7 засевали в верхние камеры, а клетки U251 засевали в нижние камеры отдельно и инкубировали в течение 24 ч перед обработкой. После трехкратной промывки PBS в верхнюю и нижнюю камеры на указанное время добавляли 1 мкМ GpC-ODN, CpG-ODN, гель CpG-RCA и гели CpG-MCA в свежей среде. Клетки U251 из нижних камер собирали и проводили эксперимент по клонированию на планшете.
Влияние на пролиферацию клеток U251, совместно культивируемых с клетками RAW264.7, анализировали с помощью экспериментов по клонированию на планшете. Клетки U251 в нижних камерах собирали и разводили в нескольких пропорциях в среде DMEM, содержащей 15% FBS, до конечного количества 200 клеток / лунку на 6-луночном культуральном планшете и продолжали культивировать в течение 2 недель, пока кластеры клонов не наблюдались с помощью невооруженным глазом (более 50 клеток / клон). После осторожной промывки PBS клетки фиксировали 4% параформальдегидом в течение 30 минут, затем окрашивали кристаллическим фиолетовым в течение 1 часа. Кластеры, содержащие более 50 клеток, будут включены в подсчет как успешное формирование клона. Эксперименты повторяли трижды, и в каждом эксперименте было три повторяющихся лунки:скорость образования клонов =(количество образованных клонов / количество инокулированных клеток) × 100%.
Лиофилизированные гели CpG-MCA или гель CpG-RCA помещали в 400 мкл DMEM, содержащей 10% фетальной телячьей сыворотки (10% FBS-DMEM), соответственно, и инкубировали при 37 ° C в течение 24 часов; концентрации ДНК в супернатанте измеряли с помощью NanoDrop 2000c после центрифугирования при 1000 об / мин в течение 10 с. Остальные гидрогели рассчитывали в соответствии с концентрациями ДНК в супернатанте. Оставшийся гель =( м - c × V ) / м × 100%, где м это масса геля, которую мы добавили, c - концентрация ДНК в супернатанте, а V объем супернатанта. Гели CpG-MCA или гель CpG-RCA помещали в 10% FBS-DMEM или PBS соответственно и инкубировали при 37 ° C в течение 12 или 24 часов. После инкубации гели запускали в 1% агарозном геле при 100 В в течение 60 мин.
Одноцепочечный CpG-ODN был синтезирован Sangon Company (Пекин, Китай) и очищен с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ). CpG-ODN, использованный в этом исследовании, был CpG-ODN 1668 [18]:5'-TCCATGACGTTCCTGATGCT, а олигонуклеотиды, не относящиеся к CpG, были названы GpC-ODN [18]:5'-TCCATGAGCTTCCTGATGCT.
Все данные в этом исследовании представлены как средние значения ± стандартное отклонение (среднее ± стандартное отклонение) из двух или трех независимых экспериментов. Статистический анализ между разными группами проводился с помощью t Стьюдента. контрольная работа. Статистическая значимость была установлена на уровне P . <0,05 (уровень достоверности 95%).
Иммуностимулятор ДНК был успешно сконструирован посредством реакции MCA [16]. В этой реакции первым и важным этапом перед изотермической амплификацией является реакция циклизации длинной однонитевой (ss) матрицы (Схема 1). Мы использовали электрофорез в агарозном геле, чтобы подтвердить образование кольцевой ДНК после отжига и лигирования с праймером (дополнительный файл 1:Рисунок S1). Расстояние миграции между праймером и длинной матрицей оцДНК стало короче, поскольку структура изменилась после того, как матрица превратилась в цикл и лигировалась с праймером. Реакция MCA содержит цепочку последовательно повторяющихся звеньев и легко приводит к появлению наноцветков из-за непрерывной намотки (схема 1). Результаты электрофореза в агарозном геле показали, что продукты реакции RCA (гель CpG-RCA или R12) и реакции MCA (гель CpG-MCA или R4M4 / R4M8) были получены успешно (рис. 1а). Гель CpG-RCA и гели CpG-MCA было трудно мигрировать через агарозный гель, и они оставались удерживаемыми в исходном положении. Оба геля были слишком липкими, поэтому их вытащили как шелк, когда мы их пипетировали (дополнительный файл 1:рисунок S2A – C). Эти результаты предполагают, что матрица ДНК экспоненциально амплифицируется свободными дНТФ с образованием гидрогеля [15]. Стоит отметить, что расстояние миграции не имело значительной разницы между гелями CpG-RCA и гелями CpG-MCA. Кроме того, результаты SEM и TEM дополнительно показали, что диаметр гелей CpG-RCA (рис. 1b, e) и гелей CpG-MCA (рис. 1c, d, f и g) варьировался от нанометрового до микрометрового масштаба, и демонстрирует морфологию наноцветков. CpG-ODN необходимо интернализовать в TLR9-положительные иммунные клетки и взаимодействовать с TLR9, который локализован в эндосомах внутри иммунных клеток, чтобы выполнять свою иммуностимулирующую активность. Для этого мы использовали Cy5-меченный CpG-ODN, гель CpG-RCA и гели CpG-MCA для исследования их поглощения клетками RAW264.7 с помощью конфокальной микроскопии. Как видно на фиг. 2а, гели CpG-MCA, меченные Cy5, эффективно интернализовались и распределялись в цитоплазме клеток RAW264.7. На основании измеренной средней интенсивности флуоресценции эффективность поглощения гелей CpG-MCA была значительно увеличена ( P <0,0001) по сравнению с CpG-ODN (фиг. 2b), демонстрируя эффективное поглощение гелей CpG-MCA, что благоприятно для оказания более сильной иммунной стимуляции. Сообщается, что поглощение ДНК клетками мышиных макрофагов RAW264.7 было увеличено за счет создания различных наноструктур ДНК. Такие как тетраподоподобная структурированная ДНК (тетраподна), тетраэдрическая ДНК (тетраэдр) и тетрагональная ДНК (тетрагон) [19]. Аналогичным образом было доказано, что Х-образная ДНК, Y-образная ДНК и Х-ДНК гидрогель увеличивают поглощение ДНК клетками [20, 21]. Эти результаты свидетельствуют о том, что структуры ДНК более высокого порядка являются более эффективными формами для доставки функционализированных фрагментов ДНК. Поэтому мы предположили, что высокое клеточное поглощение гелей CpG-MCA происходит из-за изменения формы гелей CpG-MCA в результате образования геля. Затем мы проверили стабильность гелей CpG-MCA. Гели CpG-MCA инкубировали при 37 ° C в течение разного времени в разных растворах. Результаты электрофореза в агарозном геле показали, что гели CpG-MCA были относительно стабильными в растворе PBS и частично разлагались в сыворотке, содержащей среду (дополнительный файл 1:рисунок S3A). Гели CpG-MCA по-прежнему выглядели как лестница, а не как одна полоса, что свидетельствовало о неполной деградации. Впоследствии мы исследовали стабильность гелей в 10% FBS-DMEM. Результаты ПЭМ подтвердили, что гели CpG-MCA были частично переварены и больше не выглядели как цветы, но все еще сохраняли форму (дополнительный файл 1:рисунок S2D – F). Кривую деградации гелей CpG-MCA строили путем определения концентрации ДНК в супернатанте в течение 24 часов (дополнительный файл 1:рисунок S3B). Было продемонстрировано, что после инкубации в течение 24 часов в 10% FBS-DMEM, R12 оставался почти 80%, в то время как R4M4 и R4M8 оставались почти 85%, что соответствовало результатам электрофореза в агарозном геле. Чтобы подтвердить разложение гелей, гели CpG-MCA инкубировали при 37 ° C до 48 часов в сыворотке, а определение разложения проводили электрофорезом в агарозном геле (дополнительный файл 1:Рисунок S4). Гели больше не выглядят как лестницы, поскольку они были расщеплены на части ферментом в сыворотке, потеряли свою вязкость и стали даже кусочками с одним нуклеотидом, что позволяет предположить, что гели CpG-MCA являются биоразлагаемыми, поскольку они могут расщепляться в присутствии фетальной бычьей сыворотки ( FBS). Соответственно, гели CpG-MCA эффективно сопротивляются перевариванию сыворотки в течение 24 часов и, наконец, полностью разрушаются. Высокая способность противостоять деградации поможет повысить его иммуностимулирующую эффективность; уровни экспрессии TNF-α и IL-6 в супернатанте клеток RAW264.7 также ясно показали, что гели CpG-MCA могут эффективно стимулировать клетки к выработке иммунных цитокинов.
Схематическая иллюстрация получения и клеточного поглощения гелей CpG-MCA и гелей CpG-RCA. Длинную одноцепочечную ДНК с фосфорилированной группой на 5'-конце смешивали с праймерами, состоящими из мотивов CpG, сначала отжигали и лигировали ДНК-лигазой Т4 с образованием кольцевой матрицы. Реакция RCA и MCA была проведена ДНК-полимеразой phi29 для создания большого количества последовательной конкатемерной ДНК и свертывания, как многие наноцветки. Затем гели могут поглощаться макрофагами и стимулировать секрецию цитокинов
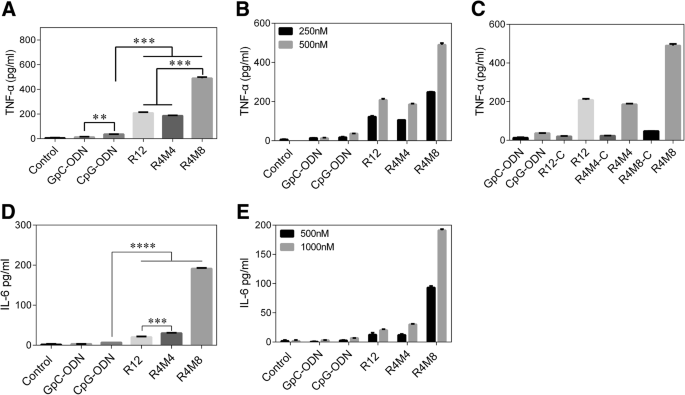
Иммуностимулирующая эффективность гелей CpG-MCA была дополнительно оценена, мы обработали макрофаги RAW264.7 GpC-ODN (последовательность изменена с эффекта GACGTT на GAGCTT) [22], CpG-ODN, гель CpG-RCA и CpG- MCA гели, а затем обнаружил эксперимент TNF-α и IL-6. Концентрация TNF-α в супернатанте клеток RAW264.7, инкубированных в течение 8 часов с гелем CpG-RCA (R12) и гелями CpG-MCA (R4M4 и R4M8), вызвала гораздо более высокий уровень TNF-α, чем CpG-ODN в такая же концентрация обработки (рис. 3а). Для гелей CpG-MCA R4M8, а не R4M4 индуцировал более высокий уровень секреции TNF-α ( P <0,001), что позволяет предположить, что секреция TNF-α усиливается по мере увеличения количества копий. Кроме того, R4M8 вызвал значительное ( P <0,001) более высокой концентрации TNF-α, чем R12, что указывает на то, что гели CpG-MCA являются более мощными стимуляторами, чем гели CpG-RCA, когда общее время реакции равно. Такая же тенденция была обнаружена и при обнаружении IL-6 (рис. 3d). Секреция IL-6, стимулированная гелями CpG-MCA и гелем CpG-RCA, была значительно увеличена. Секреция IL-6 в группах R12, R4M4 и R4M8 была в 2,97 раза выше ( P <0,0001), 4,39 раза ( P <0,0001) и 27,81 раза ( P <0,0001) от такового в группе CpG-ODN, соответственно. Секреция IL-6 в группе R4M8 была 6,33 раза ( P <0,0001) из отмеченных в группе R4M4 и 9,36 раза ( P <0,001) от отмеченного в группе R12. Стоит отметить, что секреция IL-6 не имела значительных различий между CpG-ODN, GpC-ODN и контрольными группами. Эти результаты подтвердили, что более высокая эффективность высвобождения TNF-α и IL-6 наблюдалась из клеток RAW264.7, обработанных гелями CpG-MCA, а не из CpG-ODN. Кроме того, было исследовано влияние гелей CpG-MCA с различной концентрацией на секрецию цитокинов, и мы обнаружили, что секреция TNF-α и IL-6 увеличивалась с увеличением концентрации гелей (фиг. 3b, e). Результат иммуностимулирующей способности гелей без CpG (обозначенных как R12-C, R4M4-C и R4M8-C на фиг. 3c) и GpC-ODN продемонстрировал, что они почти не индуцируют секрецию TNF-α (фиг. 3c), что указывает на то, что иммунные ответы, индуцированные гелями CpG-MCA, являются результатом мотивов CpG.
Обнаружение цитокинов, высвобождаемых из клеток RAW264.7, стимулированных CpG-ODN, гелем CpG-RCA и гелями CpG-MCA, с использованием набора для ELISA. а Секреция TNF-α из клеток RAW264.7. б Секреция TNF-α из клеток RAW264.7 после обработки гелями CpG-MCA в различных концентрациях. c Обнаружение иммуностимулирующей способности гелей CpG и гелей не-CpG (R12-C, R4M4-C и R4M8-C). г Секреция IL-6 из клеток RAW264.7. е Секреция IL-6 из клеток RAW264.7 после обработки гелями MCA в различных концентрациях. Результаты выражены как среднее значение ± стандартное отклонение двух независимых экспериментов. ** P <0,01, *** P <0,001, **** P <0,0001
Затем мы проверили экспрессию мРНК цитокинов, маркеров клеточной поверхности и TLR-9. МРНК TNF-α в группе CpG-ODN была в 1,25 раза выше экспрессии, обнаруженной в контрольной группе, а кратное изменение в группах R12, R4M4 и R4M8 было в 3,28 раза ( P <0,01), 2,53 раза ( P <0,05) и 4,57 раза ( P <0,001) из тестированных в группе CpG-ODN отдельно (дополнительный файл 1:рисунок S5A). Экспрессия мРНК IL-6 в группе CpG-ODN была в 1,01 раза выше, чем в контрольной группе. Экспрессия мРНК IL-6 в группах R12, R4M4 и R4M8 была в 3,87 раза ( P <0,01), 4,63 раза ( P <0,05) и 23,04 раза ( P <0,0001), как и в группе CpG-ODN соответственно (дополнительный файл 1:рисунок S5B).
Экспрессия мРНК TLR-9, рецептора CpG внутри клеток [4], также увеличилась во всех группах по сравнению с контрольной группой, но группы геля CpG-MCA заметно усилили экспрессию мРНК TLR-9 по сравнению с CpG. -ODN группа (дополнительный файл 1:рисунок S5C). Помимо цитокинов и TLR9, мы также обнаружили костимулирующие молекулы (CD) CD86 и CD206, которые были использованы для маркировки провоспалительных (M1) и стимулирующих рост (M2) макрофагов, соответственно [23]. Было обнаружено, что не было значительной разницы между группой CpG-ODN и контрольной группой ни по CD86, ни по CD206 (дополнительный файл 1:рисунок S5D, E). По сравнению с группой геля CpG-RCA, CD86 в группах геля CpG-MCA увеличивался в разной степени, тогда как CD206 снижался в разной степени, а группа R4M8 увеличивалась и уменьшалась больше всего, предполагая, что клетки RAW264.7, обработанные гелями CpG-MCA склонны к дифференцировке в популяцию M1 с поверхностным маркером CD86, что дополнительно подтвердило способность гелей CpG-MCA к иммунной стимуляции.
Секреция цитокинов макрофагами чрезвычайно важна в ответе на иммунную стимуляцию, а также в пролиферации и миграции макрофагов. Стимуляция макрофагов с помощью CpG-ODN увеличила бы продукцию противовоспалительных цитокинов через TLR9-зависимый путь. Цитокины, индуцированные CpG-ODN, увеличивают миграцию макрофагов и способствуют пролиферации макрофагов, подавляя экспрессию негативного регулятора клеточного цикла [24]. CpG-ODN также индуцировал экспрессию ингибитора активатора плазминогена типа 1 (PAI-1) в макрофагах, что приводило к усиленной миграции через витронектин [25]. Мы также дополнительно исследовали способность гелей CpG-MCA стимулировать миграцию макрофагов и оценили их цитотоксичность. Миграция клеток RAW264.7 индуцировалась гелями CpG-MCA как в системе миграции через лунки (фиг. 4a), так и в анализе миграции из царапинной раны (фиг. 4b). Клетки RAW264.7 культивировали в присутствии или в отсутствие гелей CpG-MCA и позволяли им мигрировать в течение 24 часов. Число миграции макрофагов в гелевых группах CpG-MCA было намного больше, чем в группе CpG-ODN (фиг. 4a). Затем мы приступили к анализу миграции царапин из ран; клеткам позволяли мигрировать в течение 24 часов, а снимки делали через 0, 6 и 24 часа. Результаты показали, что площадь царапины уменьшилась по мере заживления раны. Миграция макрофагов через 6 часов показала, что гели CpG-MCA были более эффективными, чем группа CpG-ODN, поскольку коэффициент заживления ран был выше, но нет четкой значимости среди групп R12, R4M4 и R4M8 (дополнительный файл 1:Рисунок S6A, B). И область царапины через 24 часа подтвердила, что скорость заживления области царапины в группе CpG-ODN была 1,70 раза ( P <0,05) от таковой в контрольной группе, а скорость заживления в группах R12, R4M4 и R4M8 была 1,63 раза ( P <0,001), 1,63 раза ( P <0,0001) и 2,23 раза ( P < 0.0001) of that measured in the CpG-ODN group separately. The healing rate in the R4M8 group was 1.36 times (P < 0.01) of that in the R12 group. CpG-MCA gels strongly promoted the migration of RAW264.7 cells compared to that treated with the CpG-ODN or control groups (Fig. 4c). In addition, RAW264.7 cells were cultured with a series concentration of gels for 24 h. We found that the CpG-MCA gels exhibited a negligible cytotoxicity to RAW264.7 cells due to the non-toxicity of DNA itself; on the contrary, CpG-MCA gels could stimulate RAW264.7 cell proliferation with a dose-dependent effect, which was a benefit to enhance the production of immune cytokines (Fig. 4d).
The analysises of migration and cell viability of CpG-ODN, CpG-RCA gel (R12), and CpG-MCA gels (R4M4 and R4M8) in RAW264.7 cells. а The migration assay of RAW264.7 cells induced with CpG-ODN, R12, R4M4, and R4M8. б The migration assay of RAW264.7 cells stimulated with CpG-ODN, R12, R4M4, and R4M8 for 24 h. c The scratch area analysis in the scratch migration experiment. The percentages are calculated as the ratio of the original area. г The cell viability of RAW264.7 cells treated with CpG-ODN, R12, R4M4, and R4M8 for 24 h. Results are expressed as the mean ± SD of three independent experiments. ** P <0,01, *** P < 0.001, ****P < 0.0001
For further verifying the inhibited efficiency of CpG-MCA gels for the proliferation of solid tumor cells, we estimated the inhibitory effects of CpG-MCA gels as immune stimulators for the U251 human brain glioma cells. The U251 cells was first co-cultured with RAW264.7 macrophages for 24 h, and the clone formation rate was used to check the proliferate ability of the U251 cells. As shown in Fig. 5a–f, the clone formation rate in the CpG-ODN group was 74.1% of that observed in the control group (P < 0.05), and the rates in R12, R4M4, and R4M8 groups were 45.4% (P < 0.01), 15.3% (P < 0.001), and 12.0% (P < 0.001) of that in the CpG-ODN group, respectively. The results demonstrated that the U251 cells treated with CpG-MCA gels presented a significantly lower percentage of clone formation rate compared with that treated with the CpG-ODN or control groups (Fig. 5g). It is notable that the trend of inhibitory results was in accordance with the secretion of TNF-α among groups, CpG-MCA gels exhibited a stronger effect on stimulating RAW264.7 cells to secrete cytokines to inhibit proliferation of U251 cells than CpG-ODN. Our research provided preliminary evidence to demonstrate the CpG-MCA gels have the potential to be used as an immunostimulant.
The assessment on the therapeutic effect of immunostimulatory CpG-RCA gel (R12) and CpG-MCA gels (R4M4 and R4M8) on cancer cells in vitro by plate clone formation assay. U251 cells treated with a none, b RAW264.7 cells, c RAW264.7 cells treated with CpG-ODN, d RAW264.7 cells treated with R12, e RAW264.7 cells treated with R4M4, and f RAW264.7 cells treated with R4M8. г Results of the proliferation percentage of U251 cells after co-cultured with RAW264.7 cells for 24 h. Results are expressed as the mean ± SD of three independent experiments. ** P <0,01, *** P < 0.001
In summary, we have successfully preparedCpG-MCA nanohydrogels that consist of hundreds of immunostimulatory CpG motifs to effectively deliver immune stimulus signal into cells and significantly induce the expression of immune cytokines. CpG-MCA nanohydrogels exhibited powerful anti-tumor immunity against human glioma cells, demonstrating that CpG-MCA nanohydrogels have the potential to be used as an immunostimulant for the therapy of cancer.
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
CpG-MCA hydrogel
CpG-RCA hydrogel
Multi-primed chain amplification
CpG-RCA gel
Hydrogels not containing CpG motifs
CpG-MCA gels
Rolling circle amplification Введение
Методы / экспериментальные
Материалы
Подготовка круговых шаблонов ДНК
Приготовление гелей CpG-MCA и гелей CpG-RCA
Концентрация гелей CpG-MCA и гелей CpG-RCA
Электрофорез в агарозном геле
Характеристика гелей CpG-MCA и гелей CpG-RCA
Конфокальная микроскопическая визуализация
Анализ ELISA
Анализ экспрессии генов
Анализ миграции царапин из раны
Анализ цитотоксичности
Клетки U251, культивированные совместно с клетками RAW264.7
Анализ образования клонов на планшете
Стабильность гелей CpG-MCA и гелей CpG-RCA
CpG-ODN
Статистический анализ
Результаты и обсуждение


Conclusions
Доступность данных и материалов
Сокращения
Наноматериалы
- Вы знали? Как сталь делают из железа
- 3D-печать пластика в космосе - новейшая разработка Made in Space
- Наноцеллюлоза из сине-зеленых водорослей
- 3D-наноструктуры ДНК
- Нано-гетеропереходы для солнечных элементов
- Армированный непрерывным волокном термопластический композит, полностью сделанный из природных ресурсов
- 7 удивительных вещей, сделанных из бронзы
- 3D-печатные объекты чувствуют, как пользователь взаимодействует с ними
- Метод мечения клеток с помощью микроскопии, адаптированный для использования в визуализации всего тела
- Швейцарская обработка медицинских электродов из MP35N®